Abstract
Swiss stone pine (Pinus cembra L.) may represent a vital species for afforestation at the upper altitudinal limits of forests due to its adaptability to challenging environmental conditions. In this study, we aimed to analyze the variability in ring width (radial growth, RW, earlywood, EW, latewood, LW, and latewood proportion, LWP) and the influence of different sites on the climatic resilience of twelve European provenances of Swiss stone pine to identify the best-adapted provenances. Five provenances from the Alps and seven from the Carpathians were tested across two 27-year-old field trials conducted in the Eastern and Southern Carpathians. Moderate genetic differentiation among the provenances was noted, which could ensure the success of breeding programs. Based on ring width and latewood proportion, we successfully selected the 25% best-performing provenances and recommend them for use in the provenance regions where have performed. The significant influence of the testing site and the different outcomes of the provenances necessitate extreme attention during the transfer of forest reproductive materials. Additionally, the breeding strategies applied in the two trials differed, consisting in individual selection in the Cugir trial and provenance selection in the Cârlibaba trial. The low resistance of all provenances shown in both trials, particularly in the Cugir trial, is worrying in the context of global warming.
1. Introduction
Under the influence of climate change, increases in temperature and variations in precipitation regimes are predictable outcomes influencing the future distribution of tree species [1,2], in particular at high altitudes. The severity of these changes can be seen at regional, continental, and global scales [3,4,5,6]. At present, solutions to these issues are being sought in order to identify species and varieties with high resilience to climate change [7,8,9,10,11]. Management strategies could include actions to increase the resistance of forest ecosystems to adverse conditions [12,13]. One feasible solution for stand resistance could be the inclusion of a mixture of species, including Swiss stone pine (Pinus cembra L.), at the upper altitudinal limit of forests and even higher [14,15,16,17,18,19,20,21,22,23].
The results of molecular genetic investigations have indicated low diversity among the Pinus cembra populations located in the Carpathians and Alps, especially in isolated populations [24,25,26]. Recommendations have been made to increase the number of genetically improved seed sources of Pinus cembra [27,28,29,30] to ensure high genetic diversity in future forests, an aim to which our study can contribute.
Tree ring research has been extensively performed to investigate the magnitude of climate change [4,31]. In some cases, researchers have focused on analyzing the influence of climate change on tree ring growth [6]. In the majority of studies, however, the effects of disturbances (caused by biotic and abiotic factors) have not been elucidated [32,33]. Due to its significant longevity and the extreme environmental conditions of its distribution range, Swiss stone pine is particularly suitable as a subject of tree ring research. Over the last twenty years, numerous studies concerning radial increments in Swiss stone pine took place in central Europe [34,35,36,37,38,39,40]. However, the expansion of provenance studies and the selection of Swiss stone pine provenances with high adaptability, based on wood traits, have not been achieved thus far.
In Romania, a breeding program for Swiss stone pine was initiated in 1979 and developed through the establishment of nine field trials [16,41,42,43], two of which are analyzed in this study. The challenges of the Romanian breeding program involved the initiation and maintenance of the trials in complex environmental conditions, at the upper altitudinal limit of forests, and on lands with high inclination and frequently suffering not only from competition with naturally regenerated Norway spruce but also from damage caused by large carnivores, namely, deer and bears [16,43].
In the present study, we aim to analyze the radial increment variability, site influence, and drought resilience of twelve European provenances (five from the Alps and seven from the Carpathians) of Swiss stone pine tested in two 27-year-old field trials in order to select the best-performing provenances for the next generation to be used in the breeding program. The study aims were as follows: (i) investigating the variability and genetic differentiation among the twelve provenances for annual radial growth, earlywood, latewood, and latewood proportion, in the two groups (provenances from the Alps and the Carpathians) and under the two different site conditions; (ii) analyzing the influence of climatic factors (annual and monthly air temperature and precipitation) on the radial growth of the twelve provenances; (iii) estimating the resistance, recovery, and resilience indices of the provenances after a drought event; (iv) providing recommendations of the best-adapted provenances.
2. Materials and Methods
Twelve European provenances of Swiss stone pine were tested, with seven originating from the Romanian Carpathians, and five originating from the Alps (Austria, Switzerland, and France), an experiment that is singular in Europe. These provenances cover a substantial portion of the species’ natural distribution, with a north latitude range of 4°36′ (from 45°12′ to 49°48′) and a larger east longitude range of 20°29′ (from 4°48′ to 25°17′); the altitudinal amplitude was also considerable, being of 695 m (Figure 1). The seedlings were obtained from ten to twenty-plus trees, situated at a minimum of 50 m from each other, to avoid inbreeding and ensure a high level of genetic diversity [44,45].
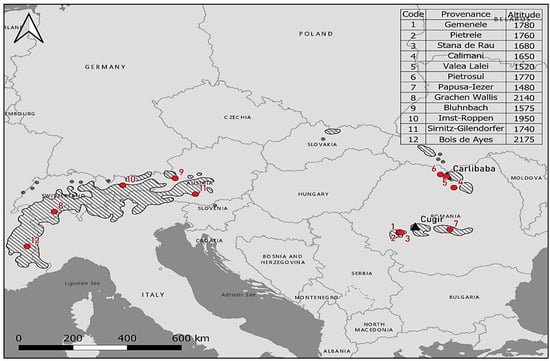
Figure 1.
Location of the Cugir and Cârlibaba comparative trials (black triangles) and provenance origins (red circles) on the Swiss stone pine natural distribution map (hatched) [46].
The Cugir (Southern Carpathians) and Cârlibaba (Eastern Carpathians) trials were established in the autumn of 1997 (using 7-year-old seedlings), in a fully randomized block design with three replications and 30 trees per subdivided plot, planted at a 2.5 × 2.5 m spacing and covering an area of 0.9 ha [44,45]. The Cugir test was performed in the Șureanu Mountains of the Southern Carpathians in a forest represented by high-altitude spruces with mull flora. The Cârlibaba trial was established in the Inău Mountains of the Eastern Carpathians in the same forest type and soil, typically districambosoil (Table 1). The average yearly temperature in the analyzed period was 54% higher in the Cugir test, whereas the amount of annual precipitation was 8% higher in the Cârlibaba trial. Over the last three decades, in both areas, while precipitation has shown no clear trend, temperatures have increased exponentially [47].

Table 1.
Environmental characteristics of the Cugir and Cârlibaba trials.
In the spring of 2024 (prior to the growing season), from nine trees of each of the twelve provenances, three in every block, cores were taken at breast height using 5 mm width increment borers, from the average Dbh trees in the subplot. The drill direction was along the level curve. The cores were dried and polished to facilitate sufficient identification of the tree rings. Thereafter, they were scanned at 2400 dpi, and ring width (RW), earlywood (EW), and latewood (LW) were measured using the CooRecorder 7.4 program [47]. The ring series were cross-dated with CDendro 7.4 software [48]. The LWP was determined as the percentage of LW from the RW. Next, all individual growth series were standardized according to ref. [49]. To remove the age effect, the methodology proposed by refs. [50,51] was applied. The standardized ring width indices (RWIs) were joint into mean chronology for each Swiss stone pine provenance with biweight [50,52]. Standardization and chronology development were achieved by the detrend and chron functions of the dplrR software [53], in R [54].
The high quality of the 2004–2023 RWI chronologies was verified using different dendrochronological parameters (Ar 1 = first-order autocorrelation, EPS = expressed population signal, and Rbar = mean correlation among trees) (Table 2). With EPS > 0.98, the resulting chronologies were reliable for estimating climate–growth relations. The climatic data were downloaded with the climate downscaling tool [46].
Table 2.
Radial increments (±SD) and core signal of the Swiss stone pine provenances.
Mean-effect ANOVA (in each trial, with replication and provenance as factors) and factorial ANOVA (between trials, with locality and provenance as factors) were performed using the mathematical models recommended by Nanson [55]. Provenance ranking and related homogenous groups separation were conducted using Duncan’s test at a 5% transgression possibility. To capture and more accurately assess the reactions of provenances originating from different geographical regions, we divided the 12 provenances into two origin groups: Carpathians and Alps.
To estimate the quantitative genetic differentiation between provenances, the QST coefficients were determined according to Morgan [56]:
where σb2 and σw2 represent the genetic variances between and within provenances [45].
QST = σb2/(σb2 + σw2)
To analyze the climate effects, the RWIs of all provenances at both sites were correlated with monthly temperature and precipitation. For each wood trait, data from the previous (June–December) and current year (January–October) were correlated with the current-year RWIs. The correlations were performed with the treeclim package [57] in the R program [54].
To determine whether the Swiss stone provenances exhibited different responses to climatic excesses, the resistance (Rt), recovery (Rc), and resilience (Rs) indices were calculated according to the method of Lloret et al. [58]. The years 2012 (Cugir) and 2013 (Cârlibaba) were those in which the provenances exhibited an abrupt decline in growth changes; as a result, they were marked as pointer years by the poiter.norm() function in the dplR package [53]. All indicators were determined using the untransformed RW [59,60], with the pointRes package in R [61].
The software utilized for data processing included R [54], Statistica 10.0 [62], Microsoft Office Excel 15, and ArcGIS 10.3.
3. Results
3.1. Wood Trait Variability and Genetic Differentiation Among the Provenances
In the Eastern Carpathians (EC, Cârlibaba trial), the average ring width (RW) of the twelve Swiss stone provenances was 3.547 mm/year, 3.1% lower (statistically insignificant) than the average value recorded in the Cugir trial in the Southern Carpathians (SC). In the Cârlibaba trial, the Carpathian provenances were superior, with a 4.5% increase in RW, compared to provenances of Alps origin. In the Cugir trial, the opposite trend was shown, with an increase of 1% in RW for the Alps provenances (Table 2); both differences were statistically insignificant. The best-performing provenance in both trials, with almost the same RW average values, was the French provenance 12—Bois des Ayes. The Romanian 4—Călimani (EC)—and the Austrian 9—Bluhnbach—provenances also performed better in both trials than the other provenances. In contrast, the Austrian 11—Sirnitz-Gilendorfer—the Swiss 8—Grachen Wallis—and the Romanian 6—Pietrosul (EC)—provenances registered low radial increments in both trials (Table 3).
Table 3.
Ranking of the provenances for ring width (RW). Duncan’s test, homogeneous groups (Hg) for α = 5%.
Regarding ring width (RW) and earlywood (EW), significant to highly significant influences of locality (testing site) and provenance were registered (0.05 < p < 0.001) by applying factorial ANOVA over the locations; in comparison, for latewood (LW), only the provenance played a significant role. For latewood proportion (LWP), no significant influences of locality, provenance, and their interaction were found. The RW measured in successive five-year intervals was also significantly influenced by locality and, in some cases, provenance (three of the four intervals, except for the period of 2009–2013) and the locality–provenance interaction (2009–2013 and 2019–2023). When mean-effect ANOVA was applied in each trial, some different results were recorded. Thus, in the Cugir trial, no significant influences of environmental (replication) and genetic (provenance) factors were noted. In contrast, in the Cârlibaba trial, provenance played a significant role in determining RW (p = 0.0044) and EW (p = 0.0243). In this case, none of the factors significantly influenced LW and LWP (Table S1).
In the Cugir trial, the average earlywood (EW) of the twelve provenances was 2.648 mm/year; in contrast, in the Cârlibaba trial, the average EW value was 4% lower (distinctly significant difference, p < 0.01). The Alps provenances performed 5% better (statistically significant, p < 0.05) than those of Carpathian origin in the southern trial (Cugir); in comparison, in the Eastern Carpathians trial, the provenances of Carpathian origin performed better by 4.5% (Table 2), but this difference was statistically insignificant. In both trials, the Eastern Carpathian provenances were superior to those of southern origin (2%–5%, statistically insignificant). The French provenance 12—Bois des Ayes—and the Romanian provenance 4—Călimani (EC)—performed better than the others in both trials, with the Austrian provenance 11—Sirnitz-Gilendorfer—showing the opposite trend (Table 4).
Table 4.
Ranking of the provenances for earlywood (EW). Duncan’s test, homogeneous groups (Hg) for α = 5%.
For latewood (LW), almost the same mean values were registered in both trials, with a greater value (statistically significant in the Cugir trial) for the Carpathian provenances (contributed to by those in the south) compared to the Alps ones in both trials (Table 2). Two Southern provenances, 3—Stâna de Râu—and 7—Păpușa-Iezer—exhibited superior radial growth in the late period of the growing season and in both trials; in comparison, the previously highlighted provenances, 12—Bois des Ayes—and 4—Călimani—only performed well in one trial, with the French origin ranking top in the Cârlibaba trial (Table 5).
Table 5.
Ranking of the provenances for latewood (LW). Duncan’s test, homogeneous groups (Hg) for α = 5%.
The latewood proportion (LWP), a key indicator of wood strength, registered an average of 28.2% in the Cârlibaba trial and a 2.2% lower value in the Cugir trial. The Carpathian provenances were superior in both trials (Table 2); however, the statistic was only significant (p < 0.01) in the Cugir trial. The Southern Carpathian provenances, particularly 3—Stâna de Râu—together with the Alps provenance 10—Imst–Roppen—registered higher LWP values in both trials, an opposite trend compared to that of the Eastern Carpathian provenances and of some provenances originating from the Alps. In addition, two Alps provenances, one from Austria (9—Bluhnbach) and one from France (12—Bois des Ayes), exhibited opposite results at the two testing sites, being ranked at the top position in the Eastern Carpathians trial (Figure 2).
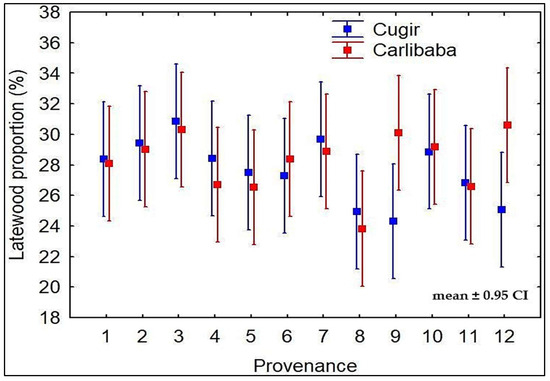
Figure 2.
Latewood proportion of the provenances in the Cugir and Cârlibaba trials (mean ± 0.95 CI).
Quantitative genetic differentiation (QST) among the provenances (Table 6) was, on average, 19.5% higher in the Cârlibaba trial, with the highest value (0.28) recorded for EW and LWP in the Cugir trial and for RW (0.47) in the Cârlibaba trial (Table 6). With regard to these ring width traits, in the Southern Carpathians trial (Cugir), the within-provenance variances were 1.3 to 1.7 times higher than the among-provenance variances, suggesting superior within-provenance variability and emphasizing the necessity for promoting individual selection. In contrast, in the Eastern Carpathians trial (Cârlibaba), high genetic variability among the provenances was registered for RW and EW, suggesting the necessity to promote adequate provenance selection. For LW and, particularly, for LWP (within-provenance variances were 2.5 times higher than among-provenance variances), an individual selection process could be applied.
Table 6.
Quantitative genetic differentiation (QST) among the provenances.
3.2. Climate and Growth Patterns of the Provenances
For the 2003–2023 period, in both trials, the De Martonne aridity index (AI) indicated a favorable climate for Swiss stone pine (85 in the Cugir trial and 102 in the Cârlibaba trial). However, in the Cugir trial, the smallest AI value (64) was registered in the year prior to the year with the lowest average annual RW; in the Cârlibaba trial, the lowest RW was registered after two consecutive years with a low AI value.
Based on the Rbar results (Table 2), the synchronicity of tree-ring series in the two groups (Alps and Carpathian provenances) was almost similar in both trials. The two groups of provenances showed similar patterns for the standardized growth indices, with the highest growth depression of the 2010–2017 period occurring in 2012 (Cugir) and 2013 (Cârlibaba), the years that followed the driest year/two years in the analyzed time period (2011 in the Cugir trial, with 842 mm of precipitation (−29% of the average trial precipitation) and in the Cârlibaba trial, with 989 mm of precipitation (−22% of the average trial precipitation).
In both trials, the two groups of Swiss stone pine provenances showed, in general, a similar response to all of the climatic variables (Figure 3). Between the testing sites, some different influences of climate on the detrended ring width indices (RWIs) were registered. Thus, regarding the current year’s monthly air temperature, in the Eastern Carpathians trial (Cârlibaba), the temperature in May had a positive influence on the RWI (r = 0.47 *, almost similar for the Alps and Carpathians provenances), whereas the influence of temperature in October was negative (r = −0.45 *) (Figure 3). In addition, in the Southern Carpathians trial (Cugir), precipitation in October of the current year was significantly and positively correlated with radial growth (r = 0.44 *); in comparison, temperature in the same month negatively influenced the RWI (r = −0.50 *). The amount of annual precipitation in the previous year had a direct and significant effect on the RWI (r = 0.47 *). In the Cugir trial, the level of precipitation in August of the previous year had a direct and significant effect on the RWI (r = 0.44 *); in comparison, temperature in the same month negatively influenced the RWI (r = −0.50 *). Only in the Cugir trial did the De Martonne index of the preceding year had a direct influence (r = 0.52 *) on the RWI (Figure 3).
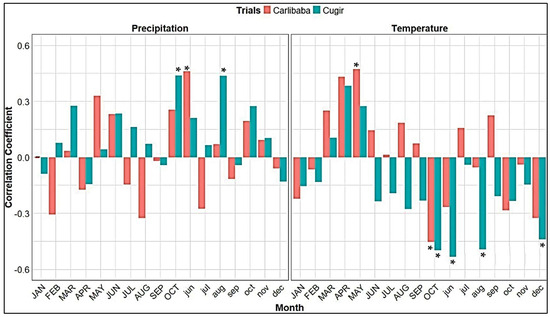
Figure 3.
Correlation coefficients between detrended ring-width series of the Swiss stone pine and monthly precipitation (left) and temperature (right). Months shown in lowercase letters refer to the previous year, and those shown in uppercase letters refer to the current year. *—Significant correlations at p < 0.05.
In both trials, LWP was negatively influenced by temperature in August of the current year (significant only in the Cârlibaba trial, r = −0.44 *; in Cugir, r = −0.39) and positively influenced by the precipitation rate in the same month (significant for the Alps provenances, r = 0.56 **, in the Cugir trial, and for the Carpathians provenances, r = 0.51 *, in the Cârlibaba trial).
In the Cugir and Cârlibaba trials, the twelve Swiss stone pine provenances did not differ significantly based on the recovery index results; in comparison, for the resistance and resilience indices, more significant results were registered in the Eastern Carpathians trial (Figure 4). The Carpathian provenances showed 13.5% better resilience in the Cugir trial; in the Cârlibaba test, the Alps provenances presented slightly better resilience (+4.3%). However, all provenances in both trials (particularly in the Cugir test) exhibited low resistance to drought, as shown by values lower than 1. Furthermore, the values were slightly higher than 1 for the recovery indices, and in both trials, these results can indicate that the Swiss stone pine trees could be able to recuperate after a drought episode. A high resilience of the best-performing populations for wood traits, i.e., 3—Stâna de Râu (from the Southern Carpathians)—4—Călimani (from the Eastern Carpathians)—and 12—Bois des Ayes (from the French Alps)—was registered (Figure 4).
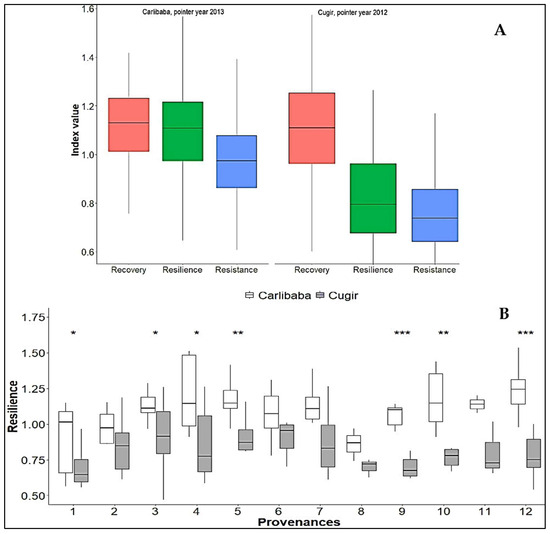
Figure 4.
Box plot showing the min, median, and max values and the quartiles for the recovery, resistance, and resilience indices of the ring-width series at the trial level (A) and the resilience of the twelve provenances at the two testing sites (B). Significant differences (*, **, ***) at p < 0.05, p < 0.01, and p < 0.001.
3.3. Predictions for Future Selection
In the Cugir trial, which took place in the Southern Carpathians, the mean performance of the provenances for the radial growth (RW) and wood strength (LWP) traits indicated that the best-performing provenances were 3—Stâna de Râu (local provenance)—4—Călimani (Eastern Carpathians)—and 10—Imst–Roppen (Austrian Alps). In comparison, 12—Bois des Ayes (French Alps)—was the best-performing provenance in terms of RW but ranked tenth for LWP. In the Cârlibaba trial, which took place in the Eastern Carpathians, the best-performing provenances for RW and LWP were 12—Bois des Ayes (the best for both traits)—9—Bluhnbach (Austrian Alps)—and 7—Păpușa–Iezer (Southern Carpathians). In comparison, the local provenance, 5—Valea Lalei—which ranked second for RW, was ranked in the eleventh place for LWP.
In the Cugir trial, quantitative genetic differentiation (QST) among the provenances indicated the necessity to promote individual selection (particularly for the best three provenances) for LWP; in comparison, as shown in the Cârlibaba trial, provenance selection (best three, 25%) for RW could be performed.
4. Discussion
Promoting Swiss stone pine provenances with active growth and a high proportion of latewood at the upper altitudinal limit of the Carpathians and Alps forests, together with local provenances, can increase mixed stands’ resistance to windthrow and snow breaks and could also ensure superior species resistance to the strong competition with fast-growing species, especially Norway spruce [11,43,45,63]. Nevertheless, the significant influence of the testing site and the different results obtained for the provenances examined in the two trials indicate the necessity of exerting extreme caution when moving forest reproductive materials. In addition, it is essential to ensure a high level of genetic diversity in future Swiss stone pine populations [24,25,26,64].
Significant results for the majority of the analyzed wood traits (except for LWP), particularly for the provenances of Alps origin, were noted in the Cugir trial compared to the Cârlibaba trial. A similar result was noted for the phenotypic traits [45]. The significant differences between the two sites’ conditions (Table 1) were related to the air temperature (1.34 °C higher in Cugir) and the geographic coordinates (2 degrees latitude and 1.5 degrees longitude lower in Cugir). The genetic component seems to play an important role in ring width, as some provenances performed well in both trials (particularly, 12—Bois des Ayes (French Alps)—and 4—Călimani (Eastern Carpathians)); yet, others also exhibited poor outcomes in both trials (8—Grachen Wallis (Swiss Alps)—11—Imst–Roppen (Austrian Alps)—and 6—Pietrosul (Eastern Carpathians)). However, a need for caution in transferring forest reproductive materials is highlighted by the contrasting results registered for the provenances 1—Gemenele—and 5—Valea Lalei (Table 3). In addition, the French provenance cannot be recommended for the Sothern Carpathian region due to the low LWP result registered in the Cugir trial, which is in complete contrast with the results obtained in the Cârlibaba trial (Figure 2). The Carpathian provenances were superior in the Cârlibaba trial for all of the analyzed traits and also for LW and LWP in the Cugir trial. In contrast, the Alps provenances performed better for RW and EW in the Cugir trial. The average RWIs of the two trials, 1.003 mm/year in Cârlibaba and 0.996 mm/year in Cugir, with slightly better results for the Carpathian provenances (by 0.3% in Cârlibaba and by 0.7% in Cugir), are in accordance with previous findings [2,15,39,65,66].
The growth dynamics of the 5-year intervals followed the trend of hard-growing species, reaching the highest RW in the 2009–2013 interval in the Cugir trial and even later (2014–2018) in the Cârlibaba trial, without significant differences between the two groups (Figure S1). Specifically, RW decreased significantly in the last period, with values of 47% and 37% in Cugir and Cârlibaba, respectively, of those registered in the previous five-year interval. In addition, for LWP, the twelve provenances registered the highest values in the first and the last 5-year periods in both trials (Figure S1). The French provenance, highlighted for its LWP in the Cârlibaba trial, showed a higher LWP in all 5-year periods; in comparison, in the Cugir trial, the LWP was reduced in all periods. Continued global warming and increasingly intense climatic perturbations require tree species to continually adapt [15,17,22]. In our study, although the climatic conditions over the last two decades have influenced the provenances tested in the Southern and Eastern Carpathians trials to differing degrees, there remain similarities. Thus, the precipitation rate of the previous year positively influenced the RWI (statistically significant in the Cugir trial only; r = 0.47 *; insignificant in the Cârlibaba trial, r = 0.31), in agreement with the results of previous studies [15,35,37,39,67]. A surprising result of the present study was the negative influence of the summer month temperature (June, July, and August) on the LWP, particularly in the Cârlibaba trial (r = −0.46 *) and for the Carpathian provenances (r = −0.55 **); in comparison, for the Alp’s provenances, the influence was statistically insignificant (r = −0.26). These findings could be the result of the exponential increase in temperatures over the last two decades, in addition to the gradually extended periods of heatwaves and drought in the summer months. In our study, the mean annual temperature increased by 1.4 °C from the 2003–2007 period to the 2019–2023 one [47]. The authors of previous studies [2,68,69], regarding RW and June–July temperatures, concluded that Swiss stone pine growth is controlled by the summer temperature; additionally, it was determined that the temperature during a short 14-day period (from the middle of June to the start of July) influenced the RW [66]. However, D’Arrigo et al. [70] and Carella [65] found in their studies that an increase in summer temperature could have a negative effect on trees growing at high altitudes because of amplified drought stress, in accordance with our findings. In both of ours trials, the temperature in May positively influenced the RWI (significantly in the Cârlibaba trial only, r = 0.47 *), in agreement with the results of Carrer et al. [35], and the precipitation rate in August had a favorable effect on the LWP (significant for the Carpathian provenances in the Cârlibaba trial (r = 0.51 *) and for Alps provenances in the Cugir trial (r = 0.56 **)). As a result of a possible extension of the growing season, a negative influence of temperature and a positive influence of precipitation, in October of the current year, on the RWI was determined for the first time.
In the Cugir trial, the low QST among the provenances indicated the necessity of promoting individual selection for LWP; in comparison, in the Cârlibaba trial, provenance selection (best three, 25%) for RW could be performed. In addition, the low-to-medium QST values suggest reduced genetic diversity, requiring a combination of the best-performing provenances from numerous studies in future afforestation programs. A high level of genetic diversity is considered an essential component of biodiversity [28,71]. In the same trials, 47%–56% lower QST values were registered for phenotypic traits, such as tree height, breast height diameter, tree volume, and branch diameter [45], suggesting that wood traits are under stronger genetic control.
5. Conclusions
Moderate genetic differentiation among provenances and a large number of provenances (12) from the entire European distribution of Swiss stone pine (from the Alps and the Carpathians, covering four countries) could ensure the success of breeding programs.
Twenty-seven years after the trials were established, we have successfully managed to select the 25% best-performing provenances based on ring width traits and recommend them for use in the provenance region where they have performed, in the two trials. The highly significant influences of the testing site and the different reactions of the provenances require extreme caution in future afforestation efforts regarding the transfer of forest reproductive materials. It must be noted that the breeding strategies applied in the two trials differed: individual selection in Cugir and provenance selection in Cârlibaba.
The significantly superior resilience and resistance recorded in the Cârlibaba trial are noteworthy, in addition to the significant recovery capacity of all provenances in both trials. However, the low resistance noted in both trials, particularly in the Cugir trial (0.76, well below the critical threshold of 1.0), is worrying in the context of the ever-greater anticipated impact of global warming.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f16030391/s1, Figure S1: Five-year dynamic of ring width (RW) and latewood proportion (LW); Table S1: ANOVA for the wood traits, by location and over locations.
Author Contributions
Conceptualization, M.B. and E.B.; methodology, M.B.; software, D.P. and E.B.; validation, M.B., D.P. and E.B.; formal analysis, M.B. and E.B.; investigation, M.B.; resources, M.B.; data curation, M.B., D.P. and E.B.; writing—original draft preparation, M.B.; writing—review and editing, M.B. and E.B.; visualization, D.P. and E.B.; supervision, M.B.; project administration, M.B.; funding acquisition, M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This paper was funded within the PN-IV-P8-8.1-PRE-HE-ORG-2024-0187 project (contract 71PHE/2024) financed by the Romanian CNCS/CCCDI-UEFISCDI.
Data Availability Statement
Data can be obtained from the authors.
Acknowledgments
We wish to express our gratitude to the anonymous reviewers for their important contribution in the manuscript improvement.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Esper, J.; Klippel, L.; Krusic, P.; Konter, O.; Raible, C.; Xoplaki, E.; Luterbacher, J.; Büntgen, U. Eastern Mediterranean summer temperatures since 730 CE from Mt. Smolikas tree-ring densities. Clim. Dyn. 2020, 54, 1367–1382. [Google Scholar] [CrossRef]
- Izworska, K.; Muter, E.; Matulewski, P.; Zielonka, T. Tree rings as an ecological indicator of the reaction of Swiss stone pine (Pinus cembra L.) to climate change and disturbance regime in the extreme environment of cliff forests. Ecol. Indic. 2023, 148, 110102. [Google Scholar] [CrossRef]
- Popa, I.; Kern, Z. Long-term summer temperature reconstruction inferred from tree-ring records from the Eastern Carpathians. Clim. Dyn. 2009, 32, 1107–1117. [Google Scholar] [CrossRef]
- Oberhuber, W.; Bendler, U.; Gamper, V.; Geier, J.; Hölzl, A.; Kofler, W.; Krismer, H.; Waldboth, B.; Wieser, G. Growth trends of coniferous species along elevational transects in the central European alps indicate decreasing sensitivity to climate warming. Forests 2020, 11, 132. [Google Scholar] [CrossRef]
- Larysch, E.; Stangler, D.F.; Nazari, M.; Seifert, T.; Kahle, H.P. Xylem phenology and growth response of European beech, silver fir and scots pine along an elevational gradient during the extreme drought year 2018. Forests 2021, 12, 75. [Google Scholar] [CrossRef]
- Izworska, K.; Muter, E.; Fleischer, P.; Zielonka, T. Delay of growth release after a windthrow event and climate response in a light-demanding species (European larch Larix decidua Mill.). Trees 2022, 36, 427–438. [Google Scholar] [CrossRef]
- Nabuurs, G.; Lindner, M.; Verkerk, H.; Gunia, K.; Deda, P.; Michalak, R.; Grassi, G. First signs of carbon sink saturation in European forest biomass. Nat. Clim. Chang. 2013, 3, 792–796. [Google Scholar] [CrossRef]
- Liang, J.; Crowther, T.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.; McGuire, A.D.; Bozzato, F.; Pretzsch, H.; et al. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, 196. [Google Scholar] [CrossRef] [PubMed]
- Ammer, C. Diversity and forest productivity in a changing climate. New Phytol. 2019, 22, 50–66. [Google Scholar] [CrossRef]
- Bastin, J.F.; de Haulleville, T.; Maniatis, D.; Marchi, G.; Massaccesi, E.; Mollicone, D.; Papa, C.; Pregagno, C. Tree Restoration Potential in the European Union; FAO: Rome, Italy; European Commission Directorate General for Environment (DG ENV): Brussels, Belgium, 2020. [Google Scholar] [CrossRef]
- Chakraborty, D.; Ciceu, A.; Ballian, D.; Benito Garzón, M.; Bolte, A.; Bozic, G.; Buchacher, R.; Čepl, J.; Cremer, E.; Ducousso, A.; et al. Assisted tree migration can preserve the European forest carbon sink under climate change. Nat. Clim. Chang. 2024, 14, 845–852. [Google Scholar] [CrossRef]
- Spathelf, P.; Stanturf, J.; Kleine, M.; Jandl, R.; Chiatante, D.; Bolte, A. Adaptive measures: Integrating adaptive forest management and forest landscape restoration. Ann. For. Sci. 2018, 75, 55. [Google Scholar] [CrossRef]
- Jandl, R.; Spathelf, P.; Bolte, A.; Prescott, C.E. Forest adaptation to climate change-is non-management an option? Ann. For. Sci. 2019, 76, 48. [Google Scholar] [CrossRef]
- Li, M.-H.; Yang, J. Effects of microsite on growth of Pinus cembra in the subalpine zone of the Austrian Alps. Ann. For. Sci. 2004, 61, 319–325. [Google Scholar] [CrossRef]
- Leonelli, G.; Pelfini, M.; Battipaglia, G.; Cherubini, P. Site-aspect influence on climate sensitivity over time of a high-altitude Pinus cembra tree-ring network. Clim. Change 2009, 96, 185–201. [Google Scholar] [CrossRef]
- Blada, I.; Popescu, F. Diallel crossing in Pinus cembra: Age trends in genetic parameters and genetic gain for height. Silvae Genet. 2012, 61, 66–79. [Google Scholar] [CrossRef]
- Mosca, E.; Gugerli, F.; Eckert, A.J.; Neale, D.B. Signatures of natural selection on Pinus cembra and P. mugo along elevational gradients in the Alps. Tree Genet. Genomes 2016, 12, 6. [Google Scholar] [CrossRef]
- Neuschulz, E.L.; Merges, D.; Bollmann, K.; Gugerli, F.; Böhning-Gaese, K. Biotic interactions and seed deposition rather than abiotic factors determine recruitment at elevational range limits of an alpine tree. J. Ecol. 2018, 106, 948–959. [Google Scholar] [CrossRef]
- Tóth, E.G.; Tremblay, F.; Housset, J.M.; Bergeron, Y.; Carcaillet, C. Geographic isolation and climatic variability contribute to genetic differentiation in fragmented populations of the long-lived subalpine conifer Pinus cembra L. in the western Alps. BMC Evol. Biol. 2019, 19, 190. [Google Scholar] [CrossRef] [PubMed]
- Zięba, A.; Różański, W.; Bukowski, M.; Ciesielska, B.; Szwagrzyk, J. Distribution and habitat conditions of Pinus cembra forests in the Tatra Mountains. Dendrobiology 2019, 81, 86–96. [Google Scholar] [CrossRef]
- Dauphin, B.; Rellstab, C.; Schmid, M.; Zoller, S.; Karger, D.N.; Brodbeck, S.; Guillaume, F.; Gugerli, F. Genomic vulnerability to rapid climate warming in a tree species with a long generation time. Glob. Chang. Biol. 2021, 27, 1181–1195. [Google Scholar] [CrossRef] [PubMed]
- Gugerli, F.; Brodbeck, S.; Bebi, P.; Bollmann, K.; Dauphin, B.; Gossner, M.; Krumm, F.; Peter, M.; Queloz, V.; Reiss, G.; et al. Swiss stone pine—Portrait of a mountain forest tree. Fact Sheet 2022, 72, 16. [Google Scholar] [CrossRef]
- Gugerli, F.; Brodbeck, S.; Lendvay, B.; Dauphin, B.; Bagnoli, F.; van der Knaap, W.O.; Tinner, W.; Höhn, M.; Vendramin, G.G.; Morales-Molino, C.; et al. A range-wide postglacial history of Swiss stone pine based on molecular markers and palaeoecological evidence. J. Biogeogr. 2023, 50, 1049–1062. [Google Scholar] [CrossRef]
- Belokon, M.M.; Belokon, Y.S.; Politov, D.V.; Altukhov, Y.P. Allozyme polymorphism of Swiss stone pine Pinus cembra L. in mountain populations of the Alps and the Eastern Carpathians. Russ. J. Genet. 2005, 41, 1268–1280. [Google Scholar] [CrossRef]
- Höhn, M.; Gugerli, F.; Abran, P.; Bisztray, G.; Buonamici, A.; Cseke, K.; Hufnagel, L.; Sebastiani, F.; Quintela-Sabaris, S.; Vendramin, G.G. Variation in the chloroplast DNA of Swiss stone pine (Pinus cembra L) reflects contrasting postglacial history of populations from the Carpathians and the Alps. J. Biogeogr. 2009, 36, 1798–1806. [Google Scholar] [CrossRef]
- Salzer, K.; Gugerli, F. Reduced fitness at early life stages in peripheral versus core populations of Swiss stone pine (Pinus cembra) is not reflected by levels of inbreeding in seed families. Alp. Bot. 2012, 122, 75–85. [Google Scholar] [CrossRef]
- Saura, M.; Perez-Figueroa, A.; Fernandez, J.; Toro, M.; Caballero, A. Preserving population allele frequencies in ex situ conservation programs. Conserv. Biol. 2008, 22, 1277–1287. [Google Scholar] [CrossRef]
- Lendvay, B.; Höhn, M.; Brodbeck, S.; Mîndrescu, M.; Gugerli, F. Genetic structure in Pinus cembra from the Carpathian Mountains inferred from nuclear and chloroplast microsatellites confirms post-glacial range contraction and identifies introduced individuals. Tree Genet. Genomes 2014, 10, 1419–1433. [Google Scholar] [CrossRef]
- Wojnicka-Półtorak, A.; Celiński, K.; Chudzińska, E.; Prus-Głowacki, W.; Niemtur, S. Genetic resources of Pinus cembra L. Marginal populations from the Tatra mountains: Implications for conservation. Biochem. Genet. 2015, 5, 49–61. [Google Scholar] [CrossRef]
- Sonnenwyl, V.; Dauphin, B.; Fragnière, Y.; Clément, B.; Grünig, S.; Brodbeck, S.; Parisod, C.; Kozlowski, G.; Gugerli, F. Genetic underpinning of historical afforestation with allochthonous Pinus cembra in the northwestern Swiss Alps. Alp. Bot. 2024, 134, 1–13. [Google Scholar] [CrossRef]
- Zhang, Z. Tree-rings, a key ecological indicator of environment and climate change. Ecol. Indic. 2015, 51, 107–116. [Google Scholar] [CrossRef]
- Camarero, J.J.; Linares, J.C.; Sangüesa-Barreda, G.; Sánchez-Salguero, R.; Gazol, A.; Navarro-Cerrillo, R.M.; Carreira, J.A. The multiple causes of forest decline in Spain: Drought, historical logging, competition and biotic stressors. In Dendroecology; Amoroso, M., Daniels, L., Baker, P., Camarero, J.J., Eds.; Springer: New York, NY, USA, 2017; pp. 307–323. [Google Scholar] [CrossRef]
- Altman, J. Tree-ring-based disturbance reconstruction in interdisciplinary research: Current state and future directions. Dendrochronologia 2020, 62, 125733. [Google Scholar] [CrossRef]
- Carrer, M.; Urbinati, C. Age-dependent tree-ring growth responses to climate in Larix decidua and Pinus cembra. Ecology 2004, 85, 730–740. [Google Scholar] [CrossRef]
- Carrer, M.; Nola, P.; Eduard, J.L.; Motta, R.; Urbinati, C. Regional variability of climate–growth relationships in Pinus cembra high elevation forests in the Alps. J. Ecol. 2007, 95, 1072–1083. [Google Scholar] [CrossRef]
- Carrer, M.; Unterholzner, L.; Castagneri, D. Wood anatomical traits highlight complex temperature influence on Pinus cembra at high elevation in the Eastern Alps. Int. J. Biometeorol. 2018, 62, 1745–1753. [Google Scholar] [CrossRef] [PubMed]
- Saulnier, M.; Edouard, J.L.; Corona, C.; Guibal, F. Climate/growth relationships in a Pinus cembra high-elevation network in the Southern French Alps. Ann. For. Sci. 2011, 68, 189–200. [Google Scholar] [CrossRef]
- Popa, I.; Nechita, C.; Hofgaard, A. Stand structure, recruitment and growth dynamics in mixed subalpine spruce and Swiss stone pine forests in the Eastern Carpathians. Sci. Total Environ. 2017, 598, 1050–1057. [Google Scholar] [CrossRef]
- Știrbu, M.-I.; Roibu, C.-C.; Carrer, M.; Mursa, A.; Unterholzner, L.; Prendin, A. Contrasting climate sensitivity of Pinus cembra tree-ring traits in the Carpathians. Front. Plant Sci. 2022, 13, 855003. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Saez, J.; Corona, C.; Von Arx, G.; Fonti, P.; Slamova, L.; Stoffel, M. Tree-ring anatomy of Pinus cembra trees opens new avenues for climate reconstructions in the European Alps. Sci. Total Environ. 2023, 855, 158605. [Google Scholar] [CrossRef]
- Blada, I. Cercetări Asupra Rezistenței Genetice la Cronartium Ribicola a unor Hibrizi interspecifici (Research on the Genetic Resistance to Cronartium ribicola of Some Interspecific Hybrids). Ph.D. Thesis, Academy of Agricultural and Forestry Sciences, Bucharest, Romania, 1987; 146p. (In Romania). [Google Scholar]
- Blada, I. Memoriu de Activitate 1961–2007 (Activity Memory 1961–2007); Silvică Publishing House: Bucharest, Romania, 2019; 51p. (In Romania) [Google Scholar]
- Budeanu, M.; Popescu, F.; Besliu, E.; Apostol, E.N. Adaptability of Swiss stone pine (Pinus cembra) in two different environmental conditions of Romanian Carpathians. Appl. Sci. 2024, 14, 7428. [Google Scholar] [CrossRef]
- Blada, I.; Popescu, F. Swiss stone pine provenance experiment in Romania. II. Variation in growth and branching traits to age 14. Silvae Genet. 2007, 56, 148–158. [Google Scholar] [CrossRef]
- Budeanu, M.; Popescu, F.; Apostol, E.N.; Pleșca, I.M.; Besliu, E. Adaptability of twelve European provenances of Pinus cembra in two different branches of the Carpathians. Silvae Genet. 2025; under review. [Google Scholar]
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological maps for the main European woody species. Data Brief 2017, 12, 662–666. [Google Scholar] [CrossRef] [PubMed]
- B4EST. Adaptive Breeding for Better Forests. Climate Database. Available online: http://www.b4est.eu/ (accessed on 1 November 2024).
- Cybis Elektronik & Data AB. Cybis Dendrochronology and History; Cybis Electronic and Data: Saltsjöbaden, Sweden, 2020; Available online: www.cybis.se (accessed on 3 December 2024).
- Popa, I. Fundamente Metodologice și Aplicații de Dendrocronologie (Methodological Fundamentals and Applications of Dendrochronology); Editura Silvică: Câmpulung Moldovenesc, Romania, 2004; p. 200. [Google Scholar]
- Yamaguchi, D.K.; Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology, Applications in the Environmental Sciences. Arct. Alp. Res. 1991, 23, 120. [Google Scholar] [CrossRef]
- Leca, Ş. Creșterea Arborilor și Arboretelor în Sistemul de Monitorizare Forestieră Intensivă (Trees and Stands Growth in Intensive Forest Monitoring System). Ph.D. Thesis, Transylvania University, Brasov, Romania, 2014. [Google Scholar]
- Nechita, C.; Popa, I.; Eggertsson, Ó. Climate response of oak (Quercus spp.), an evidence of a bioclimatic boundary induced by the Carpathians. Sci. Total Environ. 2017, 599–600, 1598–1607. [Google Scholar] [CrossRef]
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; Mérian, P.; Qeadan, F.; Zang, C.; Pucha-Cofrep, D.; Wernicke, J. dplR: Dendrochronology Program Library in R; R Package Version 1.6.9; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.r-project.org/ (accessed on 5 December 2024).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://www.r-project.org/ (accessed on 5 December 2024).
- Nanson, A. Génétique et Amélioration Des. Arbres Forestières (Genetic and Forest Trees Breeding); Les Presses Agronomique de Gembloux: Gembloux, Belgium, 2004; p. 712. (In French) [Google Scholar]
- Morgan, T.J.; Evans, M.A.; Garland, T.; Swallow, J.G.; Carter, P.A. Molecular and quantitative genetic divergence among populations of house mice with known evolutionary histories. Heredity 2005, 94, 518–525. [Google Scholar] [CrossRef]
- Zang, C.; Biondi, F. treeclim: An R package for the numerical calibration of proxy-climate relationships. Ecography 2015, 38, 431–436. [Google Scholar] [CrossRef]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter-specific facilitation. Plant. Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Budeanu, M.; Apostol, E.N.; Besliu, E.; Crișan, V.E.; Petritan, A.M. Phenotypic Variability and Differences in the Drought Response of Norway Spruce Pendula and Pyramidalis Half-Sib Families. Forests 2021, 12, 947. [Google Scholar] [CrossRef]
- Van Der Maaten-Theunissen, M.; Van Der Maaten, E.; Bouriaud, O. pointRes: An R package to analyze pointer years and components of resilience. Dendrochronologia 2015, 35, 34–38. [Google Scholar] [CrossRef]
- StatSoft Inc. Statistica 10.0; StatSoft Inc.: Tulsa, OK, USA, 2010. [Google Scholar]
- Budeanu, M.; Apostol, E.N.; Popescu, F.; Postolache, D.; Ioniţă, L. Testing of the narrow crowned Norway spruce ideotype (Picea abies f. pendula) and the hybrids with normal crown form (pyramidalis) in multisite comparative trials. Sci. Total Environ. 2019, 689, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Dzialuk, A.; Chybicki, I.; Gout, R.; Maczka, T.; Fleischer, P.; Konrad, H.; Curtu, A.L.; Şofletea, N.; Valadon, A. No reduction in genetic diversity of Swiss stone pine (Pinus cembra L.) in Tatra Mountains despite high fragmentation and small population size. Conserv. Genet. 2014, 15, 1433–1445. [Google Scholar] [CrossRef]
- Carella, A. 500 Years of Forest Growth Dynamics in the High Mountain Valley of Avers, Switzerland. Master’s Thesis, ETH Zurich, Zurich, Switzerland, 2022. [Google Scholar] [CrossRef]
- Izworska, K.; Zielonka, T.; Matulewski, P.; Muter, E. Daily Climatic Data Better Explain the Radial Growth of Swiss Stone Pine (Pinus cembra L.) in High-Elevation Cliffs in the Carpathians. Forests 2023, 14, 1411. [Google Scholar] [CrossRef]
- Oberhuber, W.; Kofler, W.; Pfeifer, K.; Seeber, A.; Gruber, A.; Wieser, G. Long-term changes in tree–ring–climate relationships at Mt. Patscherkofel (Tyrol, Austria) since the mid-1980s. Trees 2008, 22, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Vittoz, P.; Rulence, B.; Largey, T.; Freléchoux, F. Effects of climate and land-use change on the establishment and growth of Cembran Pine (Pinus cembra L.) over the altitudinal treeline ecotone in the central Swiss alps. Arct. Antarct. Alp. Res. 2008, 40, 225–232. [Google Scholar] [CrossRef]
- Housset, J.M.; Tóth, G.T.; Girardin, M.P.; Tremblay, F.; Motta, R.; Bergeron, Y.; Carcaillet, C. Tree–rings, genetics and the environment: Complex interactions at the rear edge of species distribution range. Dendrochronologia 2021, 69, 125863. [Google Scholar] [CrossRef]
- D’Arrigo, R.; Wilson, R.; Liepert, B.; Cherubini, P. On the ‘divergence problem’ in northern forests: A review of the tree ring evidence and possible causes. Global Planet. Chang. 2008, 60, 289–305. [Google Scholar] [CrossRef]
- Șofletea, N.; Curtu, A.L.; Daia, M.L.; Budeanu, M. The Dynamics and Variability of Radial Growth in Provenance Trials of Norway Spruce (Picea abies (L.) Karst.) Within and Beyond the Hot Margins of its Natural Range. Not. Bot. Horti Agrobot. 2015, 43, 265–271. [Google Scholar] [CrossRef][Green Version]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).