
Resilience of Pinus pinea L. Trees to Drought in Central Chile Based on Tree Radial Growth Methods

 , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Dendrochronological Analyses
2.3. Climate Data
2.4. Statistical Analyses
3. Results
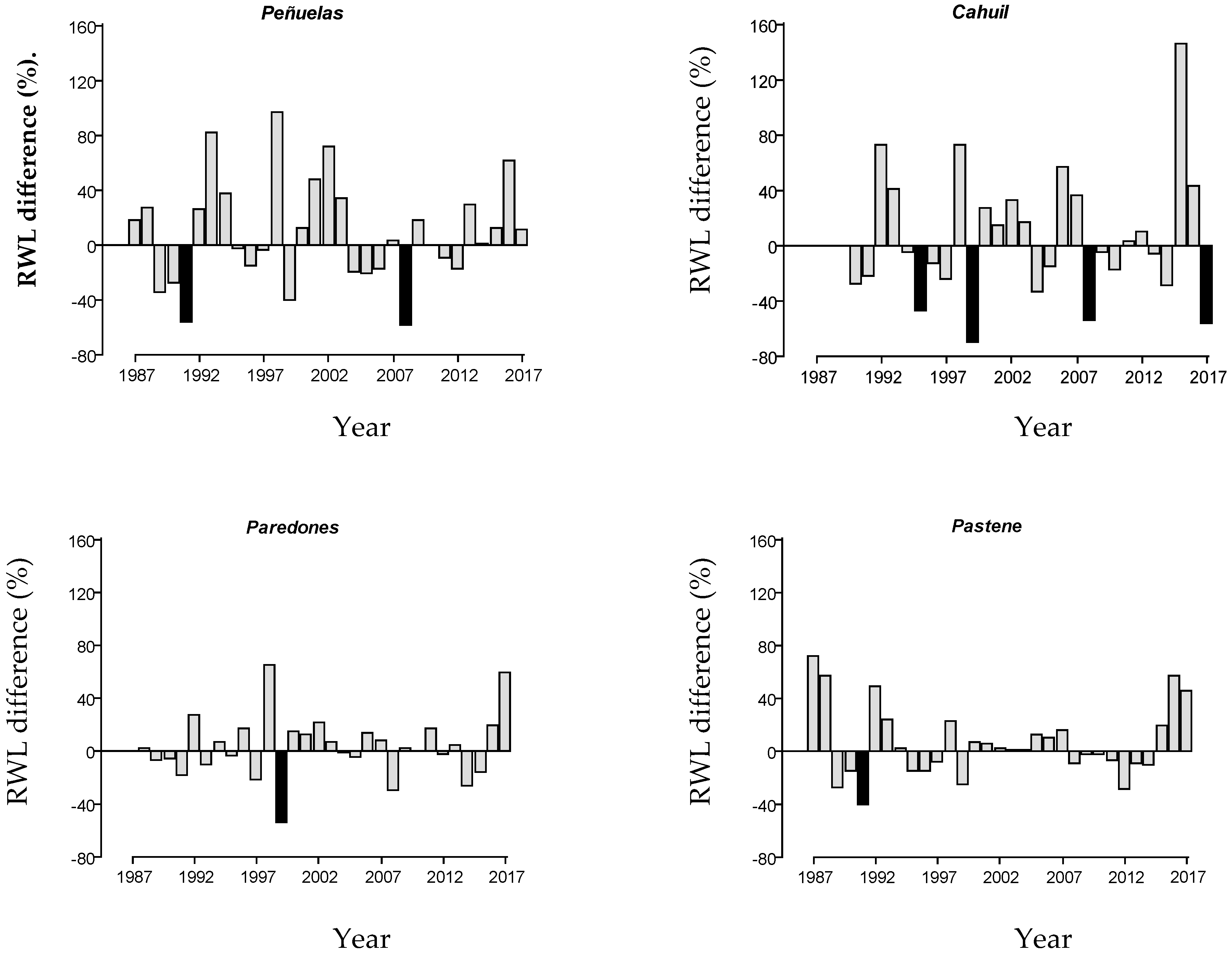
3.1. Dendrochronological Data
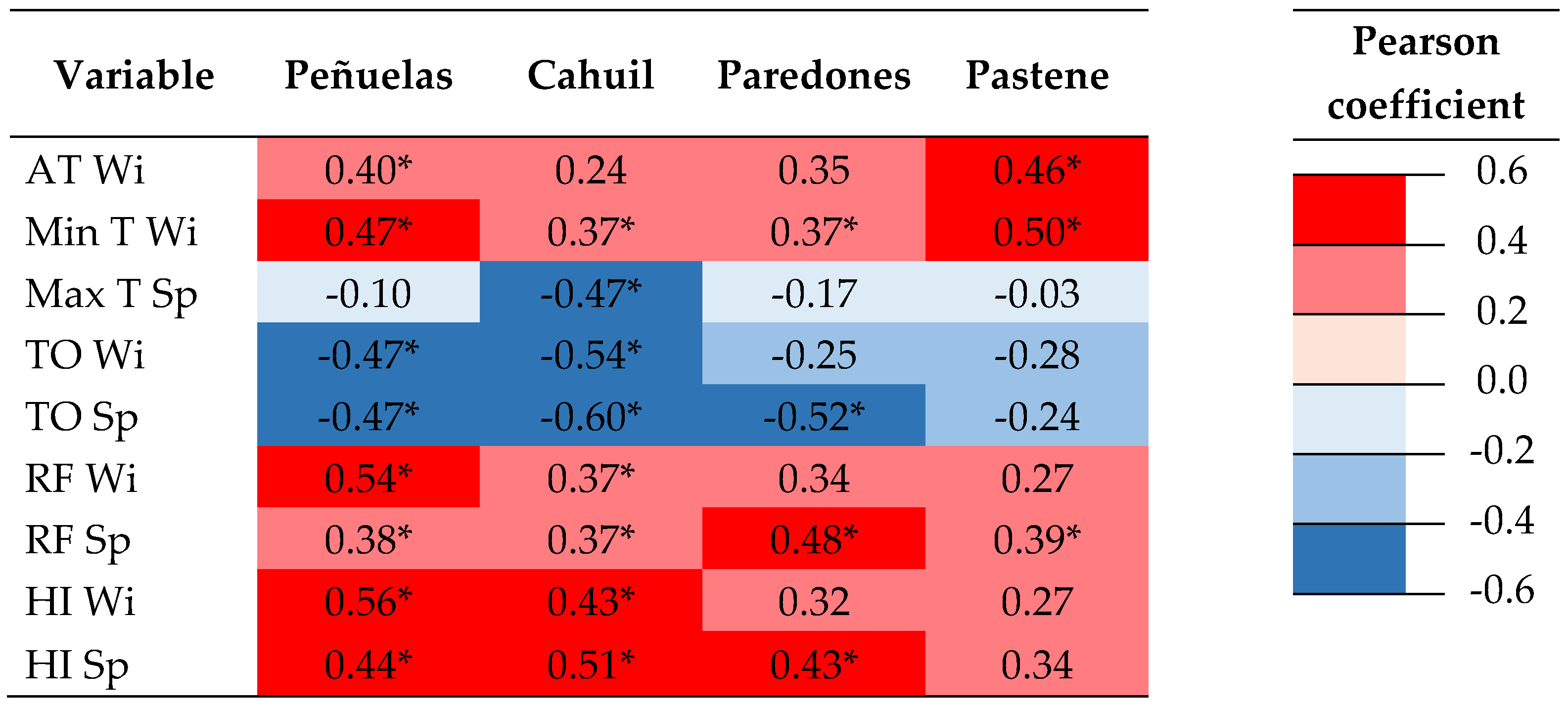
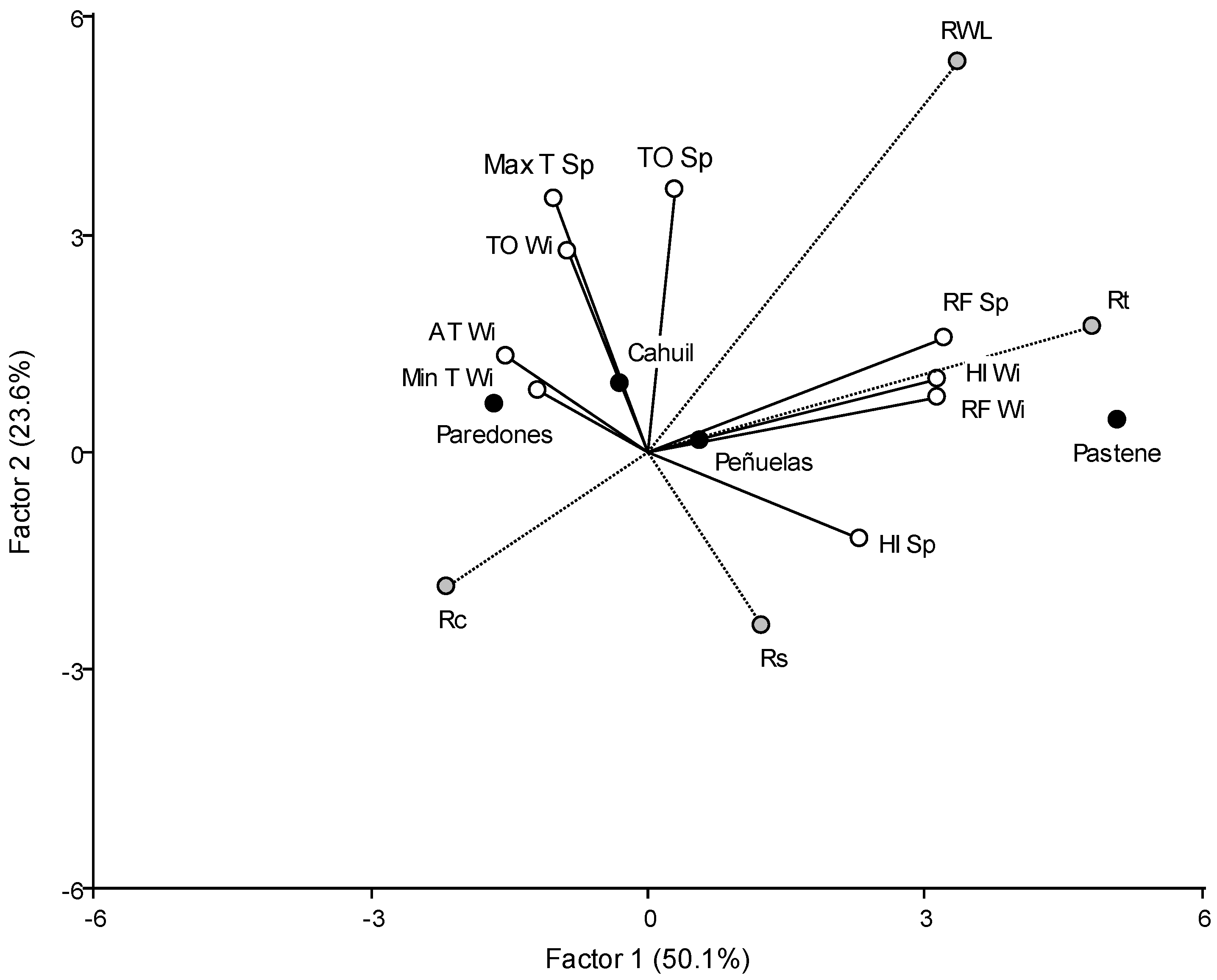
3.2. Response to Extreme Events
4. Discussion
4.1. Dendrochronological Response
4.2. Response to Droughts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cortés-Sánchez, M.; Morales-Muñiz, A.; Simón-Vallejo, M.D.; Lozano-Francisco, M.C.; Vera-Peláez, J.L.; Finlayson, C.; Rodríguez-Vidal, J.; Delgado-Huertas, A.; Jiménez-Espejo, F.J.; Martínez-Ruiz, F.; et al. Earliest Known Use of Marine Resources by Neanderthals. PLoS ONE 2011, 6, e24026. [Google Scholar] [CrossRef] [PubMed]
- Sülüsoglu, M. The Management of Villagers Owned Stone Pine (Pinus pinea L.) Plantations in Kozak Region, Turkey, a Case Study; FAO Working Paper; FAO: Rome, Italy, 2004. [Google Scholar]
- Awan, H.; Pettenella, D. Pine Nuts: A review of recent sanitary conditions and market development. Forests 2017, 8, 367. [Google Scholar] [CrossRef]
- Pinno, B.D.; Hossain, K.L.; Gooding, T.; Lieffers, V.J. Opportunities and Challenges for Intensive Silviculture in Alberta, Canada. Forests 2021, 12, 791. [Google Scholar] [CrossRef]
- Loewe, V.; Delard, C.; Balzarini, M.; Álvarez, A.; Navarro, R. Impact of climate and management variables on stone pine (Pinus pinea L.) growing in Chile. Agric. For. Meteorol. 2015, 214–215, 106–116. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2022—Impacts, Adaptation and Vulnerability; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022; pp. 3–34. [Google Scholar]
- Truettner, C.; Anderegg, W.R.L.; Biondi, F.; Koch, G.W.; Ogle, K.; Schwalm, C.; Litvak, M.E.; Shaw, J.D.; Ziaco, E. Conifer radial growth response to recent seasonal warming and drought from the southwestern USA. For. Ecol. Manag. 2018, 418, 55–62. [Google Scholar] [CrossRef]
- Trowbridge, A.M.; Adams, H.D.; Collins, A.; Dickman, L.T.; Grossiord, C.; Hofland, M.; Malone, S.; Weaver, D.K.; Sevanto, S.; Stoy, P.C.; et al. Hotter droughts alter resource allocation to chemical defenses in piñon pine. Oecologia 2021, 197, 921–938. [Google Scholar] [CrossRef]
- Garreaud, R.D.; Boisier, J.P.; Rondanelli, R.; Montecinos, A.; Sepúlveda, H.H.; Veloso-Aguila, D. The Central Chile Mega Drought (2010–2018): A climate dynamics perspective. Int. J. Climatol. 2020, 40, 421–439. [Google Scholar] [CrossRef]
- Santelices-Moya, R.; Gibson-Carpintero, S.; Cabrera-Ariza, A.; Santini-Junior, L.; Venegas-González, A. Reduced Rainfall Variability Reduces Growth of Nothofagus alessandrii Espinosa (Nothofagaceae) in the Maule Region, Chile. Forests 2022, 13, 1184. [Google Scholar] [CrossRef]
- Loewe-Muñoz, V.; del Río, R.; Delard, C.; Balzarini, M. Short-term stem diameter variations in irrigated and non-irrigated stone pine (Pinus pinea L.) trees in a xeric non-native environment. Ann. For. Sci. 2021, 78, 99. [Google Scholar] [CrossRef]
- Natalini, F.; Alejano, R.; Pardos, M.; Calama, R.; Vázquez-Piqué, J. Declining Trends in Long-Term Pinus pinea L. Growth Forecasts in Southwestern Spain. Dendrochronologia 2024, 88, 126252. [Google Scholar] [CrossRef]
- Mechergui, K.; Saleh Altamimi, A.; Jaouadi, W.; Naghmouchi, S. Climate change impacts on spatial distribution, tree-ring growth, and water use of stone pine (Pinus pinea L.) forests in the Mediterranean region and silvicultural practices to limit those impacts. iForest—Biogeosciences For. 2021, 14, 104–112. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Vergarechea, M.; Calama, R.; Pretzsch, H.; Alday, J.G.; del Río, M. Short- and long-term growth response to climate in mixed and monospecific forests of Pinus pinea and Pinus pinaster. Eur. J. For. Res. 2021, 140, 387–402. [Google Scholar] [CrossRef]
- Cutini, A. Pinus pinea L. In Pines of Silvicultural Importance; CABI, Ed.; CABI Publishing: New York, NY, USA, 2002; pp. 329–343. ISBN 0 85199 539 X. [Google Scholar]
- Perdiguero, P.; Soto, Á.; Collada, C. Comparative analysis of Pinus pinea and Pinus pinaster dehydrins under drought stress. Tree Genet. Genomes 2015, 11, 70. [Google Scholar] [CrossRef]
- Valeriano, C.; Gutiérrez, E.; Colangelo, M.; Gazol, A.; Sánchez-Salguero, R.; Tumajer, J.; Shishov, V.; Bonet, J.A.; Martínez de Aragón, J.; Ibáñez, R.; et al. Seasonal precipitation and continentality drive bimodal growth in Mediterranean forests. Dendrochronologia 2023, 78, 126057. [Google Scholar] [CrossRef]
- Balekoglu, S.; Caliskan, S.; Dirik, H.; Rosner, S. Response to drought stress differs among Pinus pinea provenances. For. Ecol. Manag. 2023, 531, e120779. [Google Scholar] [CrossRef]
- Graham, S.I.; Rokem, A.; Hille Ris Lambers, J. forestexplorR: An R package for the exploration and analysis of stem-mapped forest stand data. Ecography 2022, 2022, e06223. [Google Scholar] [CrossRef]
- Barbato, M. Construction of New Site Index Curves for the Spatial Tree Growth Model PineaFits: A Holistic Study Approach with Stem Analysis. Master’s Thesis, University of Naples, Naples, Italy, 2015; p. 61. [Google Scholar]
- Divya, K.; Kaur, S. A Study on Tree Rings: Dendrochronology using Image Processing. IOP Conf. Ser. Mater. Sci. Eng. 2021, 1022, 012115. [Google Scholar] [CrossRef]
- Newton, P.F. Wood quality attribute models and their utility when integrated into density management decision-support systems for boreal conifers. For. Ecol. Manag. 2019, 438, 267–284. [Google Scholar] [CrossRef]
- Piraino, S. Assessing Pinus pinea L. resilience to three consecutive droughts in central-western Italian Peninsula. iForest—Biogeosci. For. 2020, 13, 246–250. [Google Scholar] [CrossRef]
- Albrich, K.; Rammer, W.; Turner, M.G.; Ratajczak, Z.; Braziunas, K.H.; Hansen, W.D.; Seidl, R. Simulating forest resilience: A review. Glob. Ecol. Biogeogr. 2020, 29, 2082–2096. [Google Scholar] [CrossRef]
- Mazza, G.; Cutini, A.; Manetti, M.C. Site-specific growth responses to climate drivers of Pinus pinea L. tree rings in Italian coastal stands. Ann. For. Sci. 2014, 71, 927–936. [Google Scholar] [CrossRef]
- Smiley, T. An Introduction to Tree-Ring Dating; University of Arizona Press: Tucson, AR, USA, 1968. [Google Scholar]
- Larsson, L.; Larsson, P. CDendro and CooRecorder; Cybis Elektronik and Data AB: Saltsjöbaden, Sweden, 2018. [Google Scholar]
- Holmes, R. Computer assisted quality control in tree ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Schulman, E. Dendroclimatic Changes in Semiarid America; University of Arizona Press: Tucson, AR, USA, 1956. [Google Scholar]
- Fritts, H. Tree Rings and Climate; Academic Press: New York, NY, USA, 1976. [Google Scholar]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Muñoz-Sabater, J.; Dutra, E.; Agustí-Panareda, A.; Albergel, C.; Arduini, G.; Balsamo, G.; Boussetta, S.; Choulga, M.; Harrigan, S.; Hersbach, H.; et al. ERA5-Land: A State-of-the-Art Global Reanalysis Dataset for Land Applications. Earth Syst. Sci. Data 2021, 13, 4349–4383. [Google Scholar] [CrossRef]
- Ashaolu, E.D.; Iroye, K.A. Rainfall and Potential Evapotranspiration Patterns and Their Effects on Climatic Water Balance in the Western Lithoral Hydrological Zone of Nigeria. Ruhuna J. Sci. 2018, 9, 92. [Google Scholar] [CrossRef]
- Di Rienzo, J.; Casanoves, F.; Balzarini, M.; Gonzalez, L.; Tablada, M.; Robledo, C. InfoStat Version 2024. 2024. Available online: http://www.infostat.com.ar (accessed on 14 February 2024).
- Pardos, M.; Calama, R.; Maroschek, M.; Rammer, W.; Lexer, M.J. A model-based analysis of climate change vulnerability of Pinus pinea stands under multiobjective management in the Northern Plateau of Spain. Ann. For. Sci. 2015, 72, 1009–1021. [Google Scholar] [CrossRef]
- Calama, R.; Conde, M.; De-Dios-García, J.; Madrigal, G.; Vázquez-Piqué, J.; Gordo, F.J.; Pardos, M. Linking Climate, Annual Growth and Competition in a Mediterranean Forest: Pinus pinea in the Spanish Northern Plateau. Agric. For. Meteorol. 2019, 264, 309–321. [Google Scholar] [CrossRef]
- Sohn, J.A.; Hartig, F.; Kohler, M.; Huss, J.; Bauhus, J. Heavy and frequent thinning promotes drought adaptation in Pinus sylvestris forests. Ecol. Appl. 2016, 26, 2190–2205. [Google Scholar] [CrossRef]
- Yu Karpukhin, M.; Yussef, A.M. Electing drought-resistant Pinus pinea L. (stone pine) using dendroclimatology. IOP Conf. Ser. Earth Environ. Sci. 2021, 699, 012051. [Google Scholar] [CrossRef]
- Lebourgeois, F.; Mérian, P.; Courdier, F.; Ladier, J.; Dreyfus, P. Instability of climate signal in tree-ring width in Mediterranean mountains: A multi-species analysis. Trees 2012, 26, 715–729. [Google Scholar] [CrossRef]
- Song, Y.; Sterck, F.; Sass-Klaassen, U.; Li, C.; Poorter, L. Growth resilience of conifer species decreases with early, long-lasting and intense droughts but cannot be explained by hydraulic traits. J. Ecol. 2022, 110, 2088–2104. [Google Scholar] [CrossRef]
- Castagneri, D.; Vacchiano, G.; Hacket-Pain, A.; DeRose, R.J.; Klein, T.; Bottero, A. Meta-analysis Reveals Different Competition Effects on Tree Growth Resistance and Resilience to Drought. Ecosystems 2022, 25, 30–43. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Linares, J.C.; Fajardo, A.; Colangelo, M.; Valeriano, C.; Sánchez-Salguero, R.; Sangüesa-Barreda, G.; Granda, E.; Gimeno, T.E. Differences in temperature sensitivity and drought recovery between natural stands and plantations of conifers are species-specific. Sci. Total Environ. 2021, 796, 148930. [Google Scholar] [CrossRef]
- Aguirre, A.; Moreno-Fernández, D.; Alberdi, I.; Hernández, L.; Adame, P.; Cañellas, I.; Montes, F. Mapping forest site quality at national level. For. Ecol. Manage. 2022, 508, 120043. [Google Scholar] [CrossRef]
- Aslam, M.A.; Ahmed, M.; Hassan, F.U.; Afzal, O.; Mehmood, M.Z.; Qadir, G.; Asif, M.; Komal, S. Impact of Temperature Fluctuations on Plant Morphological and Physiological Traits. In Building Climate Resilience in Agriculture; Jatoi, W.N., Mubeen, M., Ahmad, A., Cheema, M.A., Lin, Z., Hash-mi, M.Z., Eds.; Springer: Cham, Switzerland, 2022. [Google Scholar] [CrossRef]
- Sohn, J.A.; Saha, S.; Bauhus, J. Potential of forest thinning to mitigate drought stress: A meta-analysis. For. Ecol. Manag. 2016, 380, 261–273. [Google Scholar] [CrossRef]
- Montero, G. El Pino Piñonero (Pinus pinea L.) en Andalucía: Ecología, Distribución y Selvicultura; Consejería de Medio Ambiente; Junta de Andalucía: Sevilla, Spain, 2004; ISBN 84-95785-94-3. [Google Scholar]
- Fu, Z.; Ciais, P.; Bastos, A.; Stoy, P.C.; Yang, H.; Green, J.K.; Wang, B.; Yu, K.; Huang, Y.; Knohl, A.; et al. Sensitivity of gross primary productivity to climatic drivers during the summer drought of 2018 in Europe. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190747. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; Cachinero-Vivar, A.M.; Pérez-Priego, Ó.; Aspizua Cantón, R.; Begueria, S.; Julio Camarero, J. Developing alternatives to adaptive silviculture: Thinning and tree growth resistance to drought in a Pinus species on an elevated gradient in Southern Spain. For. Ecol. Manag. 2023, 537, 120936. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Site Characteristics | Peñuelas | Cahuil | Paredones | Pastene | |
|---|---|---|---|---|---|
| Location | Latitude S | 33°11′ | 34°30′ | 34°42′ | 38°12′ |
| Longitude W | 71°29′ | 72°00′ | 71°53′ | 72°60′ | |
| Elevation (m a.s.l.) | 391 | 115 | 105 | 299 | |
| Climate † | Annual rainfall (mm) | 385 | 497 | 551 | 1072 |
| Mean temperature (°C) | 13.6 | 14.1 | 14.5 | 10.9 | |
| No. dry months †† | 9 | 8 | 8 | 6 | |
| Soil | Sand (%) | 45.3 | 43.4 | 57.1 | 40.1 |
| AWC (mm) ††† | 33.9 | 38.0 | 39.5 | 61.2 | |
| FC (cm3 cm−3) | 0.236 | 0.277 | 0.259 | 0.415 | |
| pH | 5.5 | 6.2 | 5.6 | 5.8 | |
| Texture | loam | sandy-loam | sandy clay loam | loam | |
| Stand | Density (trees ha−1) †††† | 833 | 400 | 625 | 625 |
| Age (years) †††† | 43 | 34 | 30 | 41 | |
| DBH (cm) †††† | 48.5 | 30.2 | 32.8 | 40.9 | |
| Height (m) †††† | 18.7 | 6.3 | 11.1 | 14.9 | |
| Management (age in years) | Thinned and pruned (25) | Thinned (20, 30) and pruned (30) | Thinned (15) | Thinned and pruned (20) | |
| Variables | Peñuelas | Cahuil | Paredones | Pastene |
|---|---|---|---|---|
| Mean RWL (mm) | 1.94 ± 0.19 | 4.15 ± 0.48 | 5.13 ± 0.28 | 4.87 ± 0.42 |
| SD of ring width index | 0.41 ± 0.01 | 0.41 ± 0.02 | 0.19 ± 0.01 | 0.31 ± 0.03 |
| Drought years | 1991, 2008 | 1995, 1999, 2008, 2017 | 1999 | 1991 |
| Resistance index | 0.41 ± 0.01 | 0.42 ± 0.05 | 0.45 ± 0.00 | 0.59 ± 0.00 |
| Recovery index | 2.42 ± 0.40 | 2.16 ± 0.62 | 1.94 ± 0.00 | 1.77 ± 0.00 |
| Resilience index | 0.92 ± 0.12 | 0.78 ± 0.08 | 0.93 ± 0.00 | 1.02 ± 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loewe-Muñoz, V.; Del Río, R.; Delard, C.; Cachinero-Vivar, A.M.; Camarero, J.J.; Navarro-Cerrillo, R.; Balzarini, M. Resilience of Pinus pinea L. Trees to Drought in Central Chile Based on Tree Radial Growth Methods. Forests 2024, 15, 1775. https://doi.org/10.3390/f15101775
Loewe-Muñoz V, Del Río R, Delard C, Cachinero-Vivar AM, Camarero JJ, Navarro-Cerrillo R, Balzarini M. Resilience of Pinus pinea L. Trees to Drought in Central Chile Based on Tree Radial Growth Methods. Forests. 2024; 15(10):1775. https://doi.org/10.3390/f15101775
Chicago/Turabian StyleLoewe-Muñoz, Verónica, Rodrigo Del Río, Claudia Delard, Antonio M. Cachinero-Vivar, J. Julio Camarero, Rafael Navarro-Cerrillo, and Mónica Balzarini. 2024. "Resilience of Pinus pinea L. Trees to Drought in Central Chile Based on Tree Radial Growth Methods" Forests 15, no. 10: 1775. https://doi.org/10.3390/f15101775
APA StyleLoewe-Muñoz, V., Del Río, R., Delard, C., Cachinero-Vivar, A. M., Camarero, J. J., Navarro-Cerrillo, R., & Balzarini, M. (2024). Resilience of Pinus pinea L. Trees to Drought in Central Chile Based on Tree Radial Growth Methods. Forests, 15(10), 1775. https://doi.org/10.3390/f15101775