Abstract
The Chinese red pine, Pinus tabulaeformis, is one of the most important evergreen conifer trees in China. It is widely planted in southern Gansu Province and is commonly used for garden trees, hedges, windbreaks, and soil and water conservation. However, Matsucoccus sinensis, a scale insect, has become a major pest of the P. tabulaeformis forests, and its life history and biological characteristics remain unknown. In this study, we investigated the biological characteristics, male cocoon emergence, adult mating period, and egg developmental period of M. sinensis, providing valuable insights for its prevention and control. We conducted continuous observation of the different developmental stages of M. sinensis, both in the laboratory and in P. tabulaeformis fields. The least squares method was used to calculate the egg developmental period of M. sinensis. Our results showed that the scale insect is a monophagous species with one generation occurring per year. The second-instar nymphs overwinter on the needles in the medium shell and reproduce sexually, without parthenogenesis. Male M. sinensis adults reach their peak appearance at the end of April in the study region. The peak emergence of male cocoons occurred between 2:00 AM and 4:00 AM, and the adult mating period lasted from 11:00 AM to 13:00 PM. The larvae of M. sinensis attack the needles of P. tabulaeformis. The nymphs crawled and moved to the inner base of the needles, with first- to third-instar nymphs fixing themselves to the needles to feed. The effective accumulated temperature and starting temperatures for the development of M. sinensis eggs were found to be 86.1 °C and 3.5 °C, respectively. Overall, understanding the biology and life history of M. sinensis is essential for identifying key developmental stages and determining the optimal timing for pest control, ultimately aiding in the development of targeted management strategies to protect P. tabulaeformis forests from this emerging pest.
1. Introduction
Pinus tabulaeformis is an evergreen coniferous tree species in the family Pinaceae, endemic to China, and primarily distributed in Northeast China, North China, and Northwest China [1,2]. It mainly harms Pinus tabulaeformis, Masson pine, and black pine. The insect feeds on pine needles with its piercing-sucking mouthparts leading to yellowing, severe needle mortality, and forest canopy may take on a fire-scorched appearance. Which effects significantly impair tree growth and can result in tree death [3,4,5,6,7,8]. In the Gansu province of China, it is also a dominant tree in the southern forest region, owing to its adaptability to drought and cold, robust germination capacity, well-developed root system, and crucial role in ecosystem stability [9,10,11,12]. As a key afforestation and timber species, P. tabulaeformis provides significant ecological and economic benefits, including carbon sequestration, soil and water conservation, and windbreak protection [13]. However, its long-term sustainability is increasingly threatened by insect pests, such as bark beetles, wood borers, and Matsucoccus sinensis, which has emerged as a major concern [14].
M. sinensis, also known as Sonsaucoccus sinensis, belongs to the order Hemiptera, the superfamily Coccoidea, and the family Margarodidae [15,16]. It was first reported in Zhejiang and Fujian Provinces, China, in the early 1980s [17,18], and has since been recorded in multiple provinces, including Anhui, Henan, Yunnan, Guizhou, Sichuan, Shanxi, and Gansu Provinces [19]. This species is reported to infest several pine species, including Pinus massoniana, P. yunnanensis, P. tabuliformi, and P. thunbergia. Zhang et al. [20] reported that M. sinensis is a serious pest in Henan Province and has been classified as a supplementary quarantine target for forestry pests in the region. Xiang et al. [21] found that M. sinensis causes damage to P. tabulaeformis in Diebu, Tianshui, and Longnan in the southern forest region of Gansu Province. Additionally, factors such as herbaceous ground cover, stand density, and branch height are closely associated with the severity of infestation [22]. The impact on plantations appears to be more severe than on natural forests [23].
Despite its increasing importance as a forest pest, research on M. sinensis remains extremely limited, with only a few studies available on its distribution, biology, and ecological impact. The lack of scientific literature on this species makes it difficult to develop effective management strategies. Furthermore, while M. sinensis has been reported in multiple regions, most studies focus on its occurrence in specific locations, such as Diebu, rather than addressing its broader ecological and economic significance. Understanding its life cycle, reproductive biology, and population dynamics is crucial for assessing the potential threat it poses to pine forests across China. Comparisons with other species in the genus Matsucoccus provide valuable insights into its biology. Many Matsucoccus species exhibit strong host specificity, preferring to infest particular pine species [24]. They typically have one or two generations per year, with overwintering nymphal stages and synchronized adult emergence [25]. The feeding behavior of the Matsucoccus species often leads to severe physiological stress in pine trees, causing needle chlorosis, premature needle drop, branch dieback, and, in extreme cases, tree mortality [26]. Similarly to M. josephi and M. matsumurae, which have been extensively studied due to their economic impact on pine forests in the Mediterranean and East Asia, M. sinensis may also pose a significant risk if its population continues to expand unchecked [27,28,29].
Given the limited knowledge of M. sinensis biology and its potential threat to pine ecosystems, detailed research is urgently needed to clarify its life history, morphological characteristics, and seasonal population dynamics. Therefore, the primary objective of this study was to conduct a comprehensive investigation into the life history and biological characteristics of M. sinensis in Diebu, southern Gansu. By documenting and photographing morphological changes at different developmental stages, this study aims to provide a reliable basis for the rapid and accurate identification of M. sinensis life stages, facilitate effective population monitoring, and determine the optimal timing for control measures.
2. Materials and Methods
2.1. Study Area
Our study area was located in Diebu County, between the Eastern margin of the Tibetan Plateau and the Western Qin Mountains (32°56′–34°20′ N, 102°30′–104°45′ E), at an elevation of 1900–2900 m. The mean yearly temperature, precipitation, evaporation, and frost-free period were as follows: 4.3 °C, 1023.1 mm, 918.8 mm, and 100 days, respectively. The dominant tree species in the study area were Pinus tabulaeformis, Picea asperata, and Abies faxoniana, whereas the dominant broad-leaved species were Betula albosinensis and Populus davidiana.
2.2. Life History Observation
Three plots, each 1 ha in size and containing 15-year-old P. tabulaeformis were selected as observation sites in the study area. Within each plot, three subplots of 666.7 m2 were established. In each subplot, five P. tabulaeformis trees were selected for observation using a five-point sampling method. One-year-old branches parasitized by M. sinensis on the needles were chosen as observation branches from the east, south, west, and north sides of each observation tree. From January 2021 to January 2023, we conducted continuous observations—at the beginning, middle, and end of each month—on the observation branches within the P. tabulaeformis plots. This comprehensive observation encompassed their entire life cycle, including the egg hatching period, nymph molting time, overwintering stage, pupal stage, and adult duration.
2.3. Biological Characteristics Observation
A stereomicroscope (Nikon SMZ1500, Nikon Corporation, Tokyo, Japan) was used to capture the morphological characteristics of each stage. Eggs, nymphs, pupae, and adult M. sinensis were collected from the plots. Field observations were also conducted to document the emergence rhythm and mating times of male adults, oviposition behavior of female adults, locations of cocoon formation, and pupation of 3rd-instar male nymphs. Additionally, the parasitic states and feeding damage symptoms caused by 1st- and 2nd-instar nymphs on P. tabulaeformis needles were examined.
Microscopic observations were performed using a Canon EOS 700D digital single-lens reflex (DSLR) camera (Canon Inc., Tokyo, Japan). All photographs were taken to capture damage symptoms at various scales, from all P. tabulaeformis trees to the individual branches and needles.
2.4. The Rhythm of Male Cocoon Emergence and Adult Mating
The emergence and peak emergence periods of 100 male cocoons of M. sinensis were observed both in the laboratory and in the forests of southern Gansu Province. The eclosion rhythm, mating time, and mating frequency of the female and male adults were recorded under laboratory and field conditions. Fifty female and fifty male scale insects were placed in an indoor rearing box and fixed onto P. tabulaeformis needles within the observation plot for a detailed study.
2.5. Determination of the Developmental Period of Eggs Laid by Adult Female M. sinensis
M. sinensis eggs were cultivated in a constant-temperature incubator at 70% humidity. Six temperature treatments were set up, with 30 eggs per treatment, and each treatment was replicated five times. Observations were conducted every 2 h. The temperature treatments were T1 (8 °C), T2 (10 °C), T3 (12 °C), T4 (15 °C), T5 (20 °C), and T6 (25), with the corresponding development rates denoted as V1, V2, V3, V4, V5, and V6, respectively. The following formulas were used for the calculations:
- Effective accumulated temperature = K − (T − C) N
- Development rate = V − 1/V
- Calculated Value of T = T − C + KV
The “least squares” statistical method was used to determine the values of K and C for five treatments. Subsequently, the standard error (St) of the starting point of development was calculated using the derived formula.
2.6. Data Processing and Analysis Methods
Data processing and analysis were conducted using Excel 2007 and SPSS 19.0. One-way analysis of variance (ANOVA) was used to statistically analyze the emergence period of male adults, the number of mating events at different time intervals, as well as the threshold temperature and effective accumulated temperature for egg development. Linear regression analysis was performed using Origin 9.1 to simulate the changes in the developmental rate and duration of M. sinensis eggs at different temperatures.
3. Results
3.1. Damage Characteristics
In the southern part of Gansu Province, M. sinensis only harms P. tabulaeformis, without affecting other tree species. This was first observed in the Diebu County Forest area of Gansu Province, where a pine forest damaged by M. sinensis appeared yellow-brown from a distance, resembling trees that had been burned by fire (Figure 1a–c). Newly hatched M. sinensis nymphs punctured and fed on fresh needles of P. tabulaeformis. As shown in Figure 1d, the highest feeding activity occurred during the late first and second instars (Figure 1e). The second-instar nymphs had well-developed mouthparts and the needles around the mouthparts lost their green color, forming a dry ring. This led to the gradual enlargement of the coniferous macula, causing the needles to develop yellow, withered spots, which eventually resulted in needle drop and death (Figure 1c). A significant number of coniferous needles were shed, weakening the vitality of P. tabulaeformis and bringing the tree close to death (Figure 1b). This damage severely affects tree growth, causing substantial harm to P. tabulaeformis forests.

Figure 1.
Parasitism and damage of M. sinensis on P. tabulaeformis: (a) The pine forest that has been damaged appears yellow-brown from afar, resembling the appearance of being burned by fire; (b) the pines that have been affected suffer severe needle loss, appearing dry and yellow; (c) the appearance of yellow spots, loss of green, withered, and falling P. tabulaeformis needles after being damaged; (d) first-instar nymphs feeding on the inside of the needles; and (e) the second-instar larvae feed on needles.
3.2. Morphological Characteristics and Life Cycle of M. sinensis
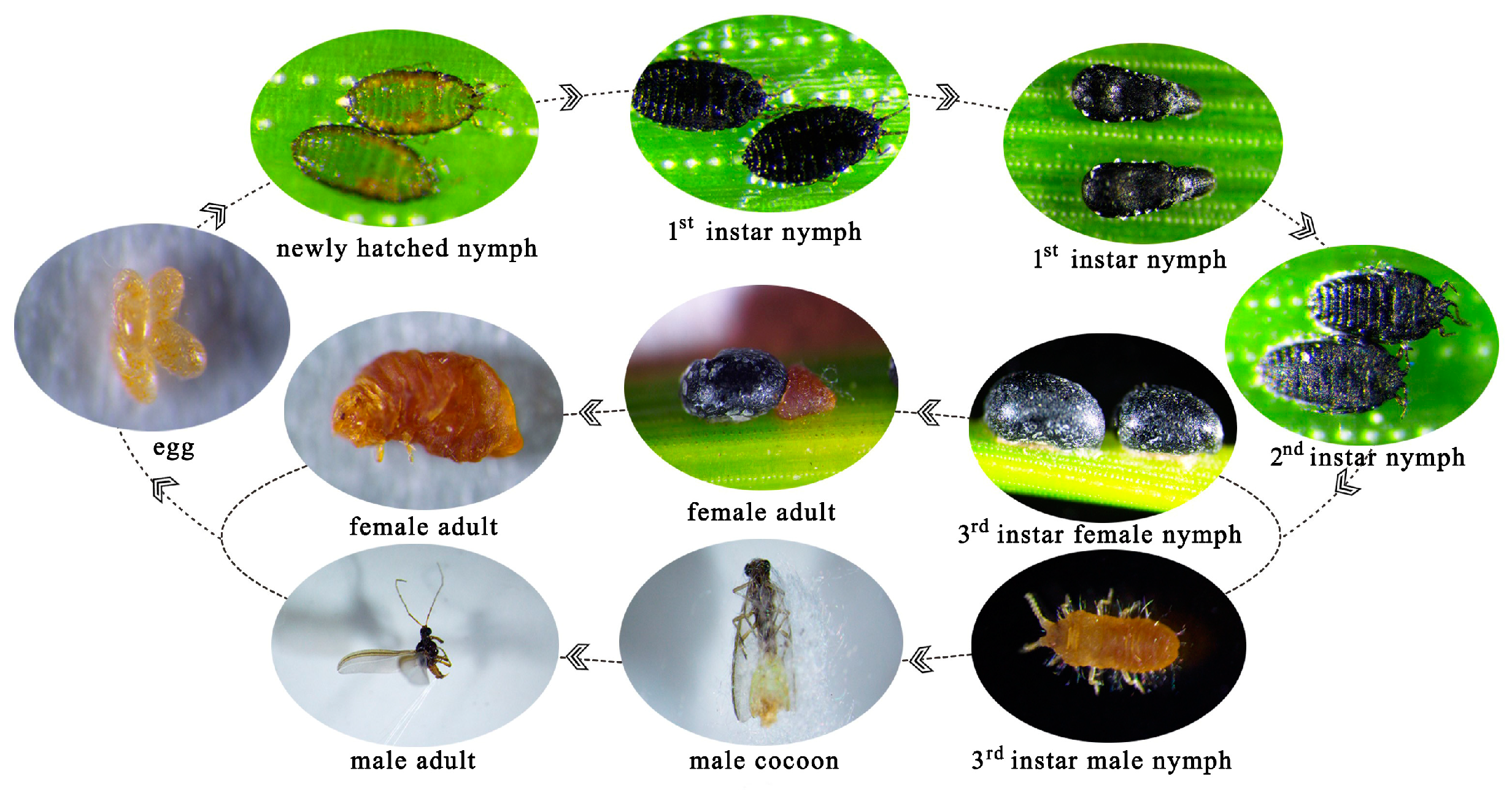
The eggs of M. sinensis are oval in shape, initially yellow in color, and later dark yellow, enclosed in a white, flocculent egg sac. Newly hatched nymphs had a nearly transparent body with fully developed appendages. The first-instar nymphs were light gray to black, with six moniliform antennae, one pair of eyes, 11 distinct body segments, and three pairs of legs (Figure 2a). During the later stages, nymphs fix themselves to the needles, and the antennae, legs, and one eye gradually disappear. The thorax and abdomen were enlarged, and the body surface adhered to a wax layer, forming a mesocell. At this point, the nymphs molted into the second instar (Figure 2a). In the second instar, the antennae, legs, and other appendages were reduced, earning the nymph the name “no-limb nymph”. The mouthparts are well developed, and the body is covered by a black hard shell resembling a black sesame seed attached to the needles. Later, male and female nymphs showed clear differentiation. The female, without limbs, has a larger, obovate body with a black shiny shell, whereas the male is smaller, oval-shaped, and also has a black shell (Figure 2a). The third-instar female nymphs grew within the shell and did not emerge (Figure 3). The third-instar male nymph is yellow, with one pair of antennae and three pairs of legs and is well developed and active in crawling (Figure 3). The adult female remains within the shell, with indistinct body segments, a pair of black monocular eyes, and degenerated antennae and legs. Male adults have filamentous antennae, a pair of compound eyes, and prominent features on both sides. The ventral tail end of the male had a hook-shaped mating device and multiple white tail filaments (Figure 2). The developmental stages of M. sinensis are shown in Figure 3.

Figure 2.
Morphological characteristics of Male M. sinensis: (a) Characteristics of the four instars of M. sinensis; (b) head, compound eye, and antennae; (c) wing and wing veins; (d) abdomen and long-wax silk.
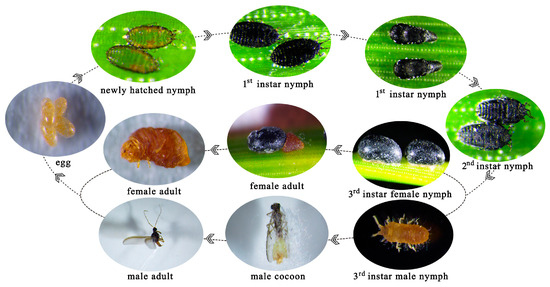
Figure 3.
Characteristics of M. sinensis instars throughout its life cycle.
3.3. Life History and Habits
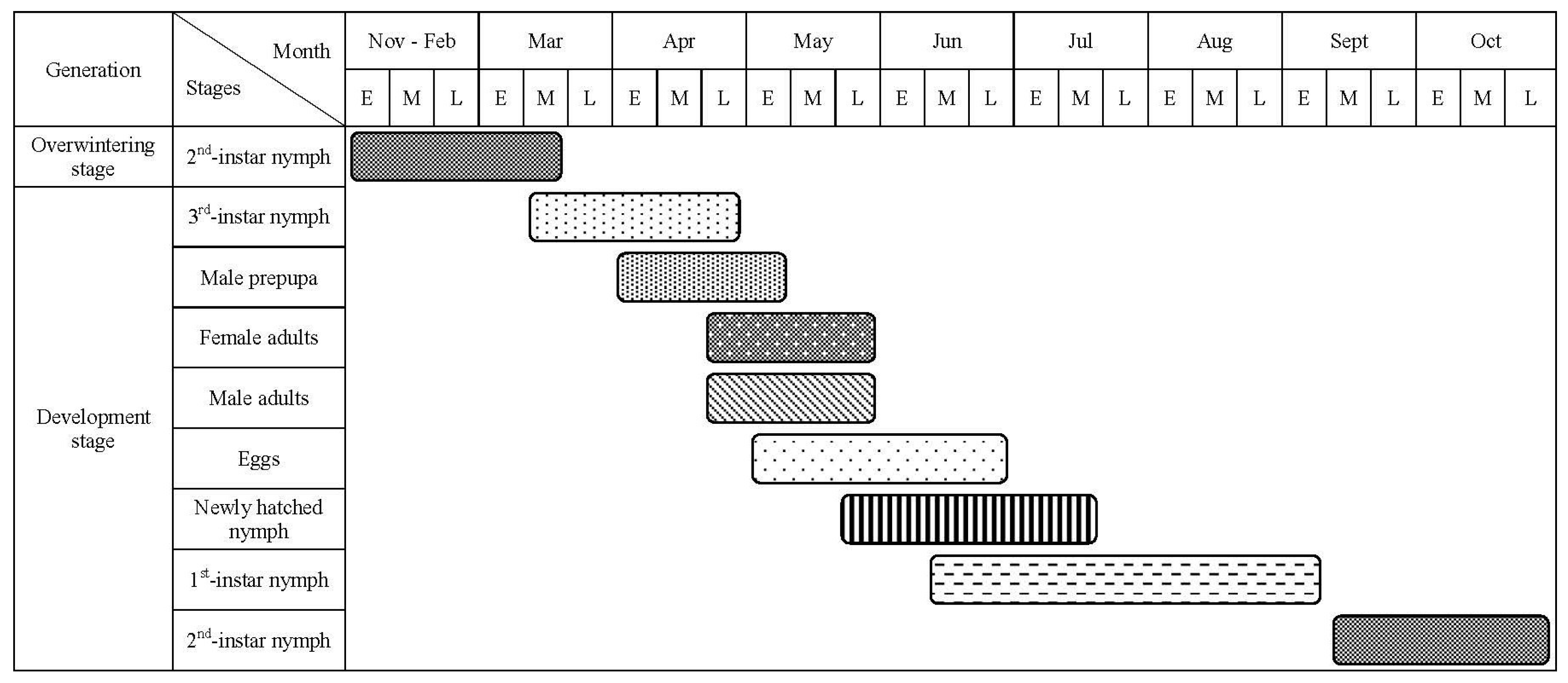
We observed that M. sinensis completes one generation over two years in southern Gansu from 2021 to 2023 (Figure 4). In mid-March, the overwintering nymphs began to move. The third-instar male nymphs exited their protective shells, crawling down branches and trunks to pupate. Male nymphs formed cocoons in the crevices of the bark at the base of the tree trunks, surrounding rocks, and fallen cones. Peak cocoon formation occurred in early April, with a large number of male pupae appearing in late April and emerging as adults in early May. Peak emergence of male adults was recorded in mid-May. The third-instar female nymphs also became active in mid-March, although morphological changes were less distinct compared to males. The peak mating between male and female adults occurred during this period. After mating, female male adults began laying eggs from early May to late July. First-hatched nymphs appeared in late May, with their diffusion peaking in late June and concluding in early August. The first-instar parasitic nymphs were present from mid-June to early September, lasting approximately three months. By mid-September, the first-instar nymphs molted into second-instar limbless nymphs. In late October, these nymphs clearly differentiated into males and females and overwintered in the second instar. Upon hatching, newly hatched nymphs produced a significant number of white wax filaments that broke off as they emerged from a round hole at the end of the scale shell of the female insect. After shedding most wax filaments, the nymphs crawled along the needles, feeding on young tissue, before settling into the gap at the base of the leaf sheath. After the dispersal of the first-instar nymphs and parasitism of the second-instar limbless nymphs, the second-instar nymphs overwintered and molted into the third-instar stage. The developmental stages of male and female nymphs were similar during the first and second instars.
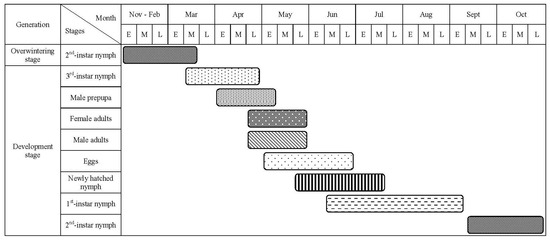
Figure 4.
Seasonal life cycle of M. sinensis in Diebu district of Gansu Province during 2021–2022 seasons. E, M, and L denote the early, middle, and late period (ten days) of a month, respectively.
3.4. Male Adult Emergence Rhythm
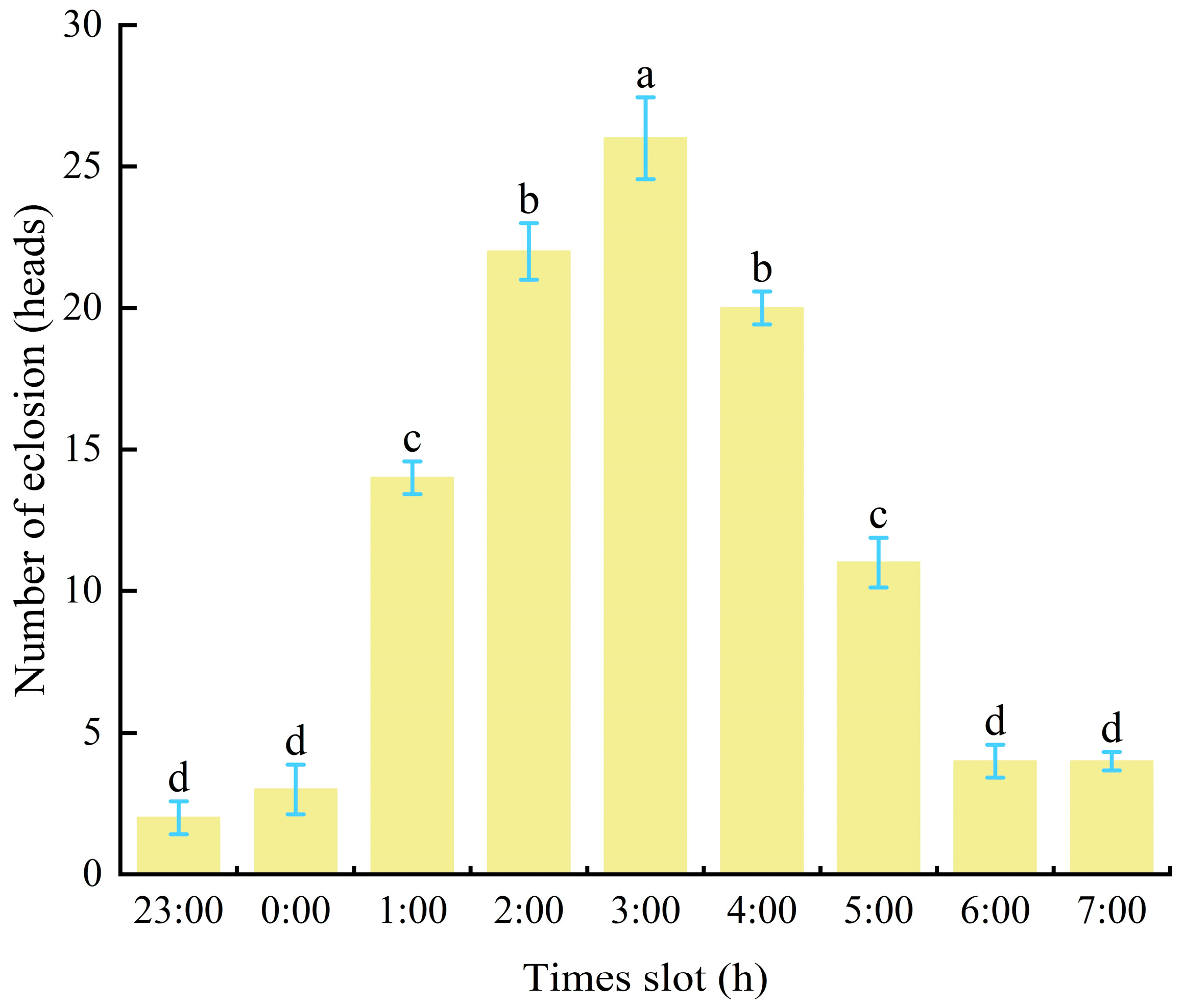
In southern Gansu forests, a large number of male cocoons appeared between April 17th and 19th, with the peak emergence of male adults occurring between April 28th and 29th. By the end of May, all adult males had emerged (Figure 5). Observations in P. tabulaeformis forests revealed that male cocoons emerged between 11:00 PM and 7:00 AM, with peak emergence occurring between 2:00 AM and 4:00 AM (Figure 5). Laboratory observations confirmed that the number of emerging male cocoons was significantly higher at 3:00 AM than at other periods (p < 0.05), with a maximum of 26 adults emerging within one hour.
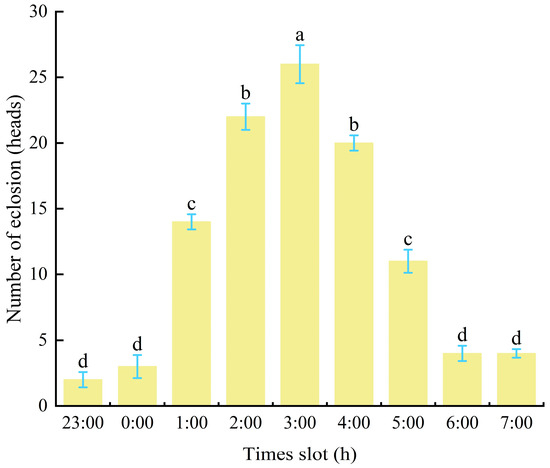
Figure 5.
Period and number of male adult emergence (individuals). Notes: Different lowercases in the bars indicated emergence signiffencantly difference in different time. Same as below.
3.5. Duration and Frequency of Adult Mating
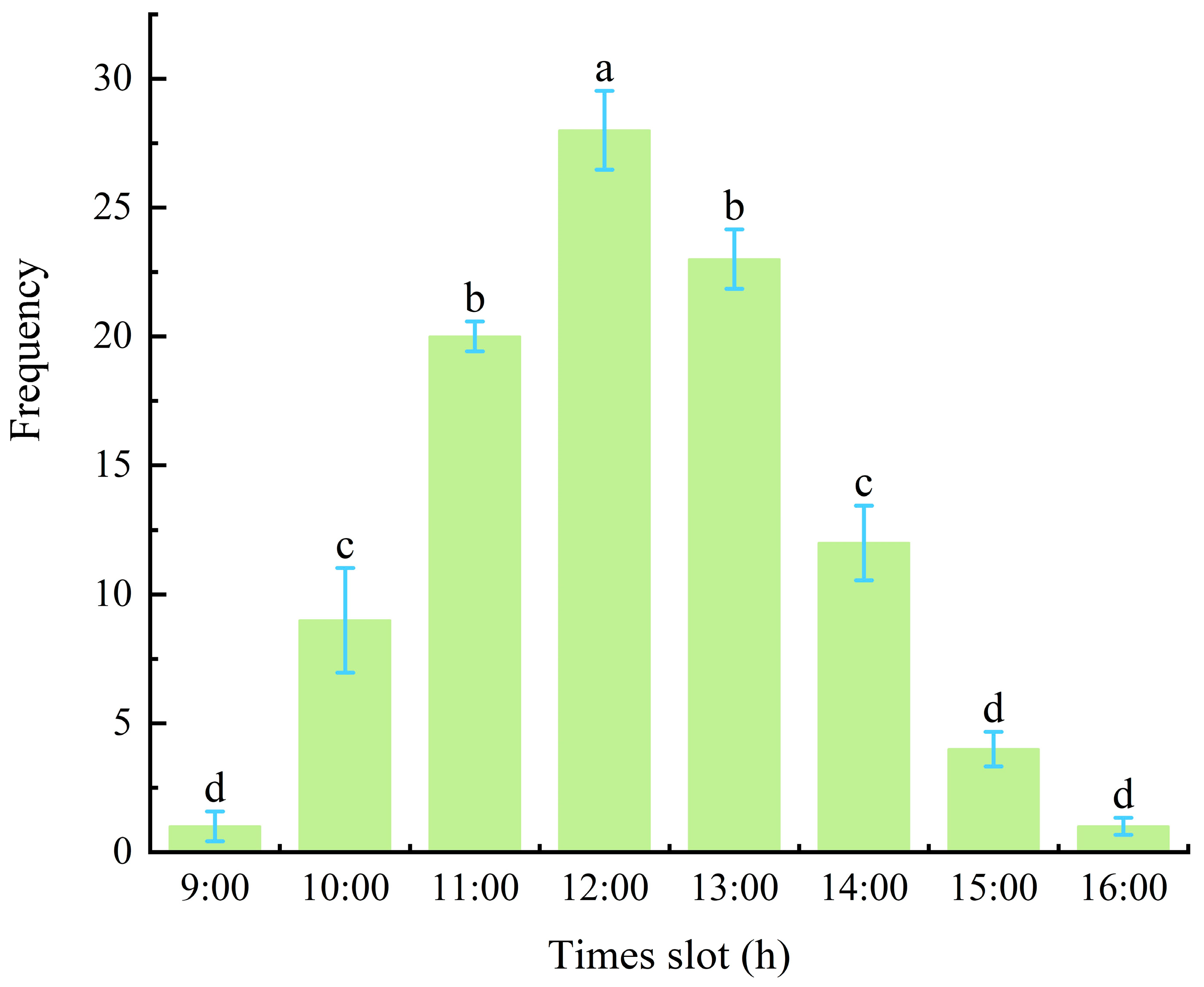
The peak mating period was at 12:00 PM, during which the highest mating frequency of 28 events per hour was recorded (Figure 6), which was significantly higher (p < 0.05) than that observed during other periods. Mating activity of M. sinensis adults was observed between 9:00 AM and 4:00 PM, and no mating behavior was recorded during the other time periods (Figure 7a). In contrast, mating frequencies at 9:00 AM and 4:00 PM were the lowest, with only two mating events per hour, which were significantly lower at other time intervals between 9:00 AM and 4:00 PM (p < 0.05).
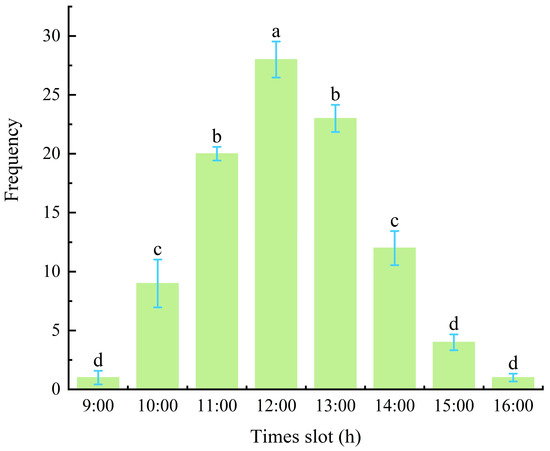
Figure 6.
The mating period and frequency of male adults (times).
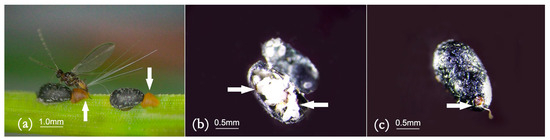
Figure 7.
Female adults of M. sinensis show tails, mates, sacs, and scale shell orifices: (a) Adult females exert their tails and mates; (b) female adults lay eggs in the scale shell and form sacs; (c) aperture of the scale shell.
3.6. Oviposition Characteristics of Female Adults
3.7. Cocoon and Pupation Sites
After reaching the third instar, male nymphs of M. sinensis cocoon and pupated in various sites, including crown depression (Figure 8a), under the bark at the base of the stem (Figure 8b), in curled dead leaves (Figure 8c), on the bark of dead trees (Figure 8d), at the base of the cones and in the phosphorous sheet (Figure 8e), and on shaded moist needles (Figure 8f). Nymphs do not cocoon or pupate on the needles or branches of P. tabulaeformis.

Figure 8.
Pupation sites of adult M. sinensis males: (a) Cone Apex Depression; (b) under the bark of a tree; (c) among the curling dead leaves; (d) on the bark of a dead tree; (e) cone base and in phosphorous tablets; (f) on the damp needles on the shaded side.
3.8. Development Rate and Duration of Eggs at Different Temperatures
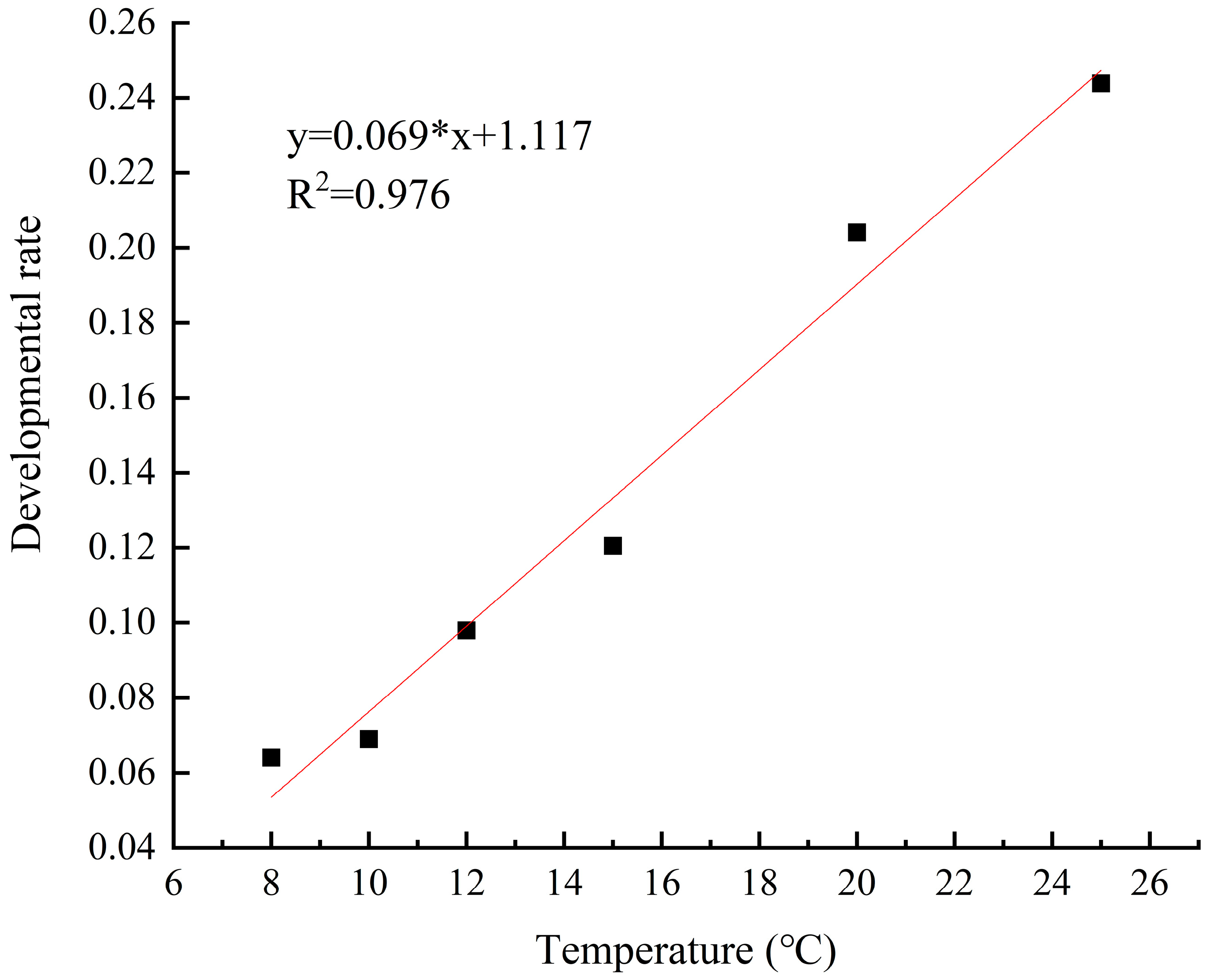
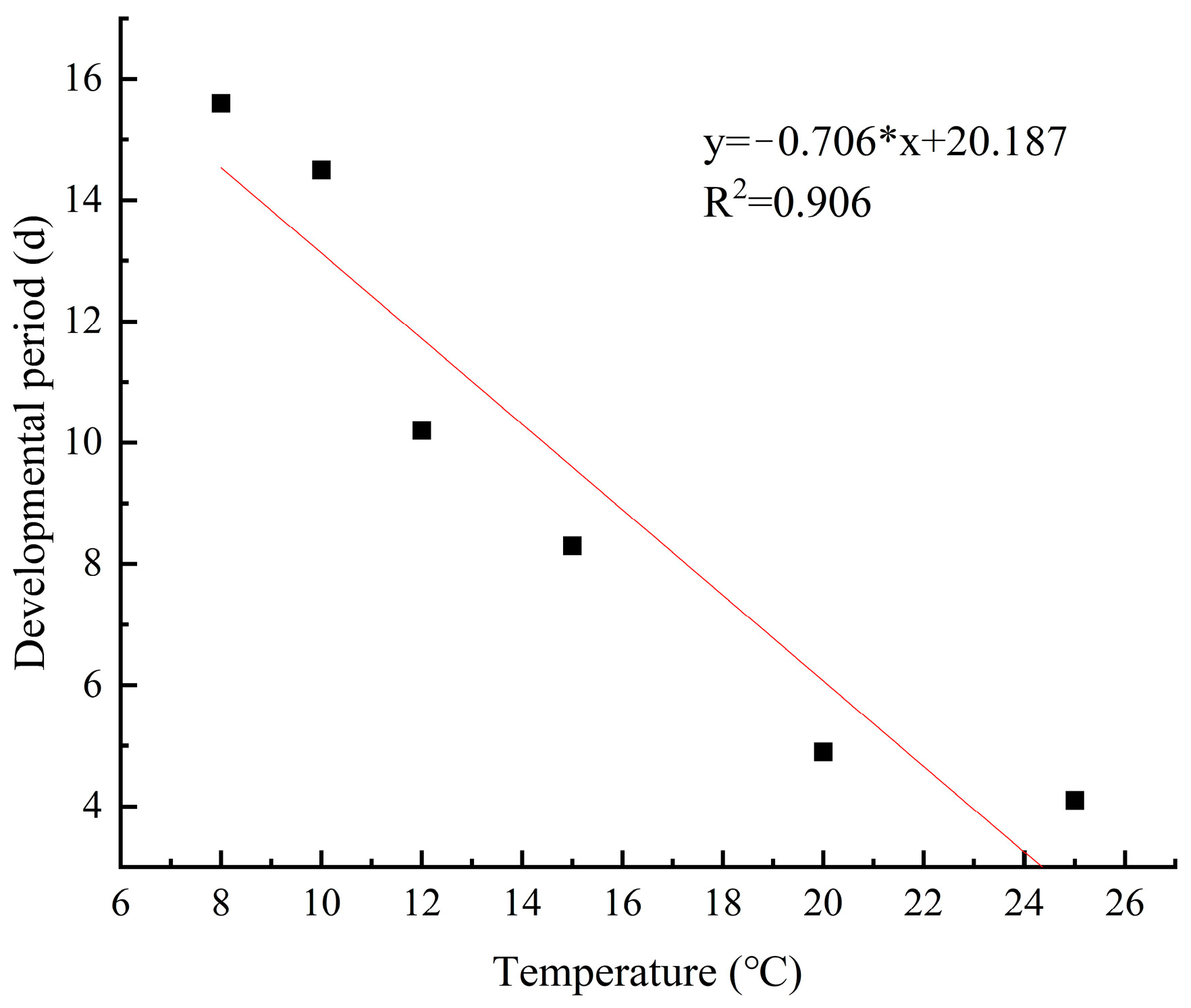
The development rate of M. sinensis eggs gradually increased and the development period gradually decreased as the temperature increased from 5 °C to 28 °C. Linear regression analysis revealed a significant linear relationship between the development rate and duration of scale eggs and temperature (R2 = 0.976; R2 = 0.906) (Figure 9 and Figure 10). At the lower temperature of 8 °C, the development rate of the eggs was 0.064, and it took 15.6 days to hatch. In contrast, at 25 °C, the development rate increased, and the development period was reduced to 4.1 days.
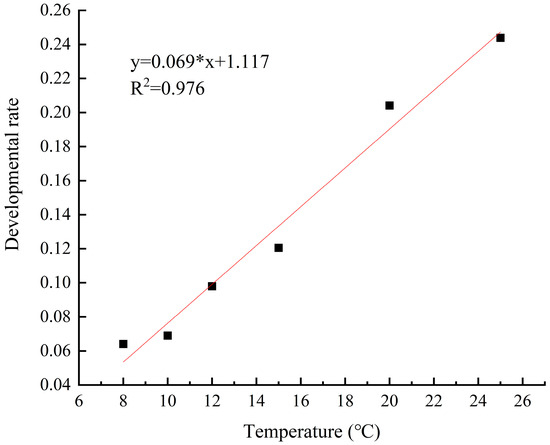
Figure 9.
Egg development rate at different temperatures.
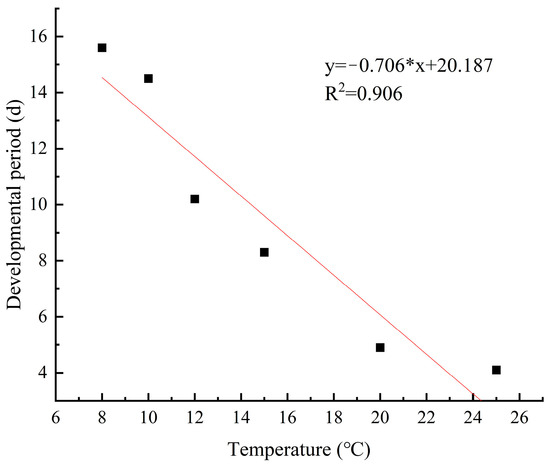
Figure 10.
Duration of Egg development at different temperatures.
3.9. Threshold and Effective Accumulated Temperature for Egg Development
The effective accumulated and threshold temperatures for the development of M. sinensis eggs were determined to be 86.1 °C (K = 86.1 °C) and 3.5 °C (C = 3.5 °C), respectively (Table 1). Therefore, the relationship between the developmental rate of the M. sinensis egg stage and temperature can be expressed by the following formula:

Table 1.
Development rate and effective accumulated temperature of M. sinensis eggs.
T′ = C + KV, T′ = 3.5 + 86.1V.
4. Discussion
The existing literature indicates that from 1980 to 2022, M. sinensis caused damage to four species of pine, including P. massoniana and P. tabuliformis, across nine provinces, including Zhejiang and Gansu [22]. To date, 113 species of pine have raised questions about whether this scale insect poses a threat to other pine species and the intrinsic factors that may limit its damage [23].
The morphological and biological characteristics of eggs, nymphs, pupae, and adults of M. sinensis have been described in previous studies [24]. In this study, we provide, for the first time, detailed information about the morphological characteristics and biological behaviors of M. sinensis at different developmental stages. These observations, which include the transparent appearance of newly hatched nymphs, the molting process of the second-instar nymph, the emergence process of female adults, the morphology of male adults, emergence, courtship and mating behavior, and the color changes in the cocoon and pupation of third-instar male nymphs, offer valuable insights. These findings provide essential information for rapid and accurate identification of the scale insect M. sinensis in the field, as well as for surveys, forecasting, and quarantine inspections.
This investigation of the life history of M. sinensis in Diebu, Gansu Province, revealed, for the first time, the peak periods of newly hatched nymphs and adult mating, providing a key window for effective pest control. Additionally, we present images of adult mating and female laying eggs within their shells for the first time. Six pupation sites for third-instar male nymphs were identified, including the tops of cones and under the bark of the branches. Notably, the male cocoons and pupation sites were dispersed and difficult to collect. Mao et al. [30] described a control method involving the collection of male cocoons in P. tabuliformis forests, although this approach remains debatable.
Indoor experiments revealed a linear relationship between the developmental rate and the duration of M. sinensis infestation at various temperatures. The effective accumulated temperature (86.1 °C) and developmental threshold temperature (3.5 °C) of the insect were determined. The hatching period of the larvae can be estimated by predicting the average temperature after egg laying. These parameters were calculated under constant indoor temperature conditions. Insect development in nature occurs under variable temperatures, and the rate of development in the field may differ from that under constant temperature conditions. Nevertheless, when combined with outdoor observations, the development rate of M. sinensis eggs provides valuable guidance for pest management.
However, there are limited reports on the application of biological control methods, such as natural enemies, sex pheromones, and entomopathogenic bacteria to manage this species of scale insect. In contrast, Matsucoccus matsumurae, an important invasive species of P. tabulaeformis in China, is controlled by natural enemies, and its sex pheromones have been documented [31].
5. Conclusions
Mstsucoccus sinensis is a monophagous insect pest that causes severe damage to multiple pine forest species across several provinces in China. It infests Pinus tabulaeformis trees, with one generation occurring per year. Infestation leads to the development of yellow spots on the needles of P. tabulaeformis, which eventually fall off. The spread of this pest is primarily facilitated by wind, rain, and the movement of newly hatched nymphs. This study documented that the second-instar nymphs of M. sinensis overwinter within their scales and cover the pine needles. Reproduction is sexual and there is no evidence of parthenogenesis. Female adults spend their entire lives within the scale covers and mate multiple times during the mating period. After emerging from the scale covers, third-instar male nymphs form cocoons at the base of tree trunks, bark crevices, or fallen cones. In the southern forests of Gansu Province, the peak occurrence of adult male and female M. sinensis occurs in early May. These findings will provide more information required for the rapid identification, monitoring, and management of M. sinensis.
Author Contributions
Conceptualization, Z.F. and D.L.; Design of the Work, Z.F., S.S., D.L. and Y.D.; Methodology, J.L., J.Y., H.Q., W.L. and D.L.; Formal Analysis, Z.F., S.S., N.S. and D.L.; Data Curation, J.Y., Y.L., W.L., W.Z. and D.L.; Resources, Z.F., S.S., J.L., J.Y., H.Q., W.L. and D.L.; Writing—Original Draft Preparation D.L., Z.F. and S.S.; Writing—Review and Editing, N.S. and Y.D.; Funding Acquisition, S.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Gansu Provincial Talent Youth Team Project in 2024 (2024RCQN02).
Data Availability Statement
The data presented in the study are deposited in the Science Data Bank, the data DOI: 10.57760/sciencedb.19867.
Acknowledgments
We are very grateful to Peng-Cheng Nie, Gansu Agricultural University, China, for his adjustments and modifications to the formatting of this manuscript and for making some valuable comments.
Conflicts of Interest
The authors declare that the research was conducted in the absence of a commercial or financial relationship that could be construed as a potential conflict of interest.
References
- Xie, Q.; Liu, Z.H.; Wang, S.H.; Li, Z.Q. Genetic diversity and phylogenetic relationships among five endemic Pinus taxa (Pinaceae) of China as revealed by SRAP Markers. Biochem. Syst. Ecol. 2015, 62, 115–120. [Google Scholar] [CrossRef]
- Li, G.; Xu, G.; Guo, K.; Du, S. Geographical boundary and climatic analysis of Pinus tabulaeformis in China: Insights on its afforestation. Ecol. Eng. 2016, 86, 75–84. [Google Scholar] [CrossRef]
- Guo, X.; Peng, C.; Li, T.; Huang, J.; Song, H.; Zhu, Q.; Wang, M. The effects of drought and re-watering on non-structural carbohydrates of Pinus tabulaeformis seedlings. Biology 2021, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xie, Y.F.; Wang, W.H.; Lv, G.Z.; Liu, B.Y. Morphological characteristics and biological properties of Pinus tabulaeformis. Mod. Agric. Sci. Technol. 2012, 16, 203. [Google Scholar]
- Fu, X.J. Occurrence and control of pests and diseases in oil pine forests in nature reserves. Mod. Rural Sci. Technol. 2025, 1, 35–36. [Google Scholar]
- Wang, W.R. Causes and Control Measures for Matsucoccus sinensis. Mod. Rural Sci. Technol. 2023, 4, 37–38. [Google Scholar]
- Zhang, X. Comprehensive analysis of the breeding and cultivation techniques and the main pest and disease control methods of Chinese red pine. Seed World 2024, 12, 192–194. [Google Scholar]
- Wang, Y. Points of cultivation and pest control technology of oil pine. World Trop. Agric. Inf. 2024, 12, 73–75. [Google Scholar]
- Che, Z.X.; Liu, X.D.; Pan, X.; Li, Y.; Jin, M.; Jing, W.M.; Wang, S.H.L.; Wang, R.X.; Zhao, W.J. The Variation characteristics of nutrients contents of main dominant tree species in Gansu Province. Ecol. Environ. Sci. 2015, 24, 237–243. [Google Scholar] [CrossRef]
- Li, D.C.; Fu, Z.L.; Liu, J.Q.; Zhang, W.Y.; Yang, J. Investigation on Matsucoccus sinensis and its natural enemies and their occurrence patterns in the southern region of Gansu Province. J. Gansu Agric. Univ. 2024, 59, 180–187. [Google Scholar] [CrossRef]
- Song, J.J.; Yang, G. Biological characteristics and life cycle of Matsucoccus sinensis in Longnan City. Shaanxi For. Sci. Technol. 2024, 52, 82–86+92. [Google Scholar] [CrossRef]
- Mo, L. Biological characteristics and control technology of Matsucoccus sinensis. Contemp. Hortic. 2023, 46, 57–58+61. [Google Scholar] [CrossRef]
- Han, C.; Li, Y.; Dong, X.; Zhao, C.; An, L. Pinus tabulaeformis forests have higher carbon sequestration potential than Larix principis-rupprechtii forests in a dryland mountain ecosystem, northwest China. Forests 2022, 13, 739. [Google Scholar] [CrossRef]
- Yan, Z.; Sun, J.; Don, O.; Zhang, Z. The red turpentine beetle, Dendroctonus valens LeConte (Scolytidae): An exotic invasive pest of pine in China. Biodivers. Conserv. 2005, 14, 1735–1760. [Google Scholar] [CrossRef]
- Liu, H.W.; Cheng, X.Q.; Kang, F.F. Changes of understory plant community in Pinus tabuliformis plantation and associated environmental explanations. Chin. J. Ecol. 2014, 33, 290–295. [Google Scholar] [CrossRef]
- Yang, P.L.; Hu, J.L.; Ren, Z.Y. On pine needle scales. Acta Entomol. Sin. 1980, 23, 42–46. [Google Scholar] [CrossRef]
- Wu, S.A. Checklist and faunistic analysis of scale insect pests (Hemi ptera: Coccoidea) in Chinese Mainland. J. Beijing For. Univ. 2009, 31, 55–63. [Google Scholar] [CrossRef]
- Li, J.Y.; Chen, W.R. Studies on Biological Characteristics and Control of Chinese Pine Shoot (Pin) Scale. Fujian For. Sci. Technol. 1987, 1, 21–29. [Google Scholar] [CrossRef]
- Cai, T.T.; Rong, C.H.; Tan, C.; Gao, J.; Liu, P.; Zhang, X.; Wang, L. Spatial Distribution Patterns and Sampling Techniques of Matsucoccus sinensis Adult. J. Southwest For. Univ. 2017, 37, 159–164. [Google Scholar] [CrossRef]
- Zhang, G.X. Relationship between occurrence of Matsucoccus sinensis and environmental factors in western Henan province. Plant Quar. 2009, 23, 11–14. [Google Scholar] [CrossRef]
- Xiang, M.; Zhang, W.Y.; Li, D.C. Risk analysis of subalpine Pinus tabulaeformis forest damaged by Matsucoccus sinensis in southern Gansu Province. Shaanxi For. Sci. Technol. 2022, 50, 61–65. [Google Scholar] [CrossRef]
- Yang, H.B. Research on integrated control technology of Matsucoccus sinensis. Seed Sci. Technol. 2019, 37, 96–97. [Google Scholar]
- Li, D.C.; Fu, Z.L.; Xu, H.X.; Cao, X.W.; Liu, J.Q. Effects of stand factors of Pinus tabulaeformis stand on damage of Matsucoccus sinensis in Bailongjiang forest area. For. Res. 2021, 34, 180–186. [Google Scholar] [CrossRef]
- Zhang, C.D.; Li, D.C.; Yang, J.; Zhang, C.A.; Ding, Y.P.; Ding, Q.D. Occurrence and harm of Matsucoccus sinensis in A’xia Provincial nature reserve of Gansu Province. Shaanxi For. Sci. Technol. 2022, 50, 40–43. [Google Scholar] [CrossRef]
- Ding, Y.P.; Zhang, C.D.; Bai, Z.L.; Hu, X.R.; Qi, W.Y.; Li, D.C. Investigation into the mortality of trees in Axia provincial nature reserve of Gansu Province. Shaanxi For. Sci. Technol. 2022, 50, 56–60. [Google Scholar] [CrossRef]
- Booth, J.M.; Gullan, P.J. Synonymy of three pestiferous Matsucoccus scale insects (Hemiptera: Coccoidea: Matsucoccidae) based on morphological and molecular evidence. Proc. Entomol. Soc. Wash. 2006, 108, 749–760. [Google Scholar]
- Foldi, I. The matsucoccidae in the mediterranean basin with a world list of species (Hemiptera: Sternorrhyncha: Coccoidea). Ann. Soc. Entomol. Fr. 2004, 40, 145–168. [Google Scholar] [CrossRef]
- Mendel, Z.; Assael, F.; Dunkelblum, E. Kairomonal attraction of predatory bugs (Heteroptera: Anthocoridae) and brown lacewings (Neuroptera: Hemerobiidae) to sex pheromones of Matsucoccus species (Hemiptera: Matsucoccidae). Biol. Control 2004, 30, 134–140. [Google Scholar] [CrossRef]
- Choi, J.; Cha, D.; Kim, D.S.; Lee, S. Review of Japanese pine bast scale, Matsucoccus matsumurae (Kuwana) (Coccomorpha: Matsucoccidae), occurring on Japanese black pine (Pinus thunbergii Parl.) and Japanese red pine (P. densiflora Siebold & Zucc.) from Korea. Forests 2019, 10, 639. [Google Scholar] [CrossRef]
- Mao, W.X.; Yuan, Q.; Fang, Q. Study on biological characteristics and experimental control of Matsucoccus sinensis in Diebu foresty of Bailongjiang. Pract. For. Technol. 2011, 6, 39–41. [Google Scholar] [CrossRef]
- Branco, M.; Franco, J.C.; Dunkelblum, E.A. Common mode of attraction of larvae and adults of insect predators to the sex pheromone of their prey (Hemiptera: Matsucoccidae). Bull. Entomol. Res. 2006, 96, 179–185. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).