Integration of Soil Moisture Factor into Light-Use Efficiency Models Improves Modeling Impact of Water Stresses on Gross Primary Production


Abstract
1. Introduction
2. Data and Methods
2.1. Flux Sites
2.2. TL-LUE Model
2.3. VPM
2.4. Model Input Data
2.5. Modeling the Impact of Soil Moisture Stress on GPP
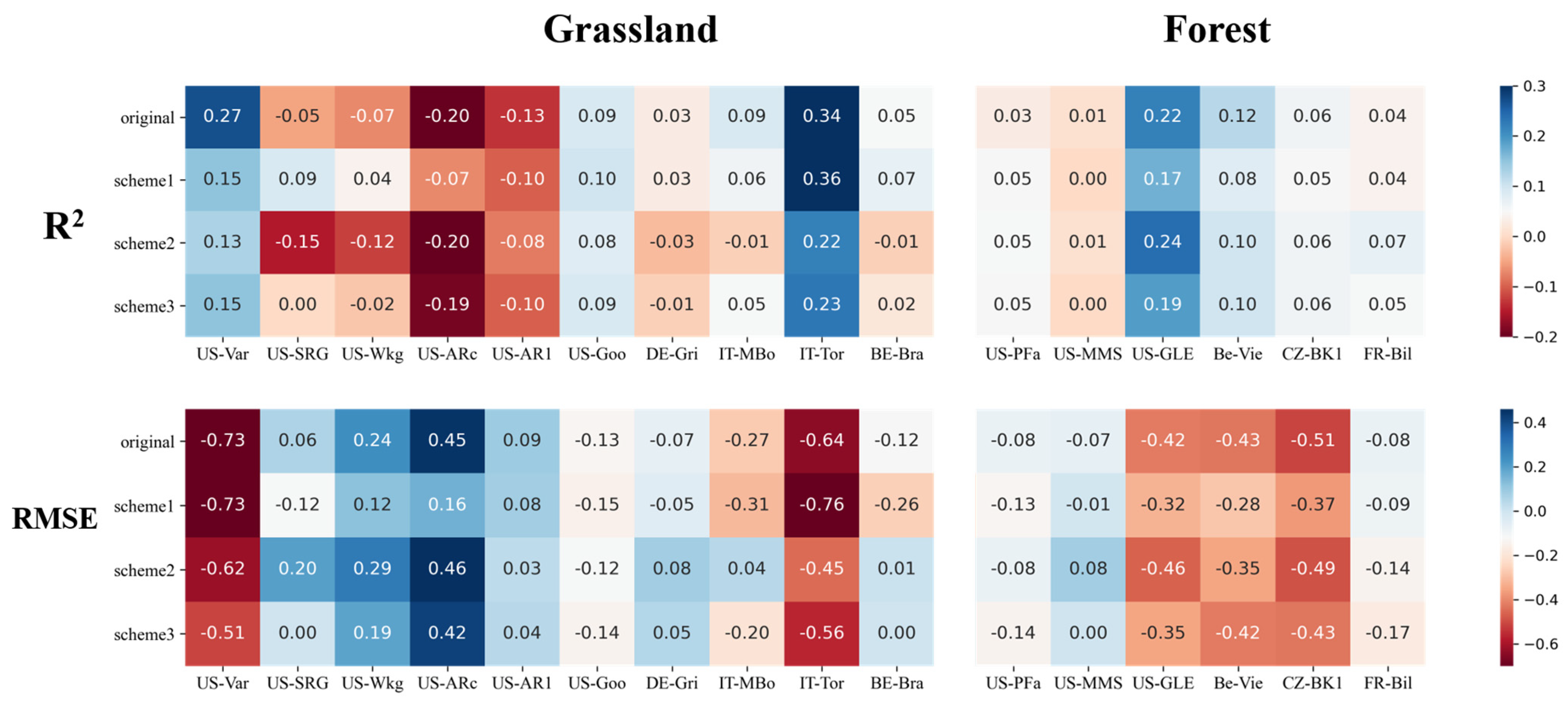
3. Results
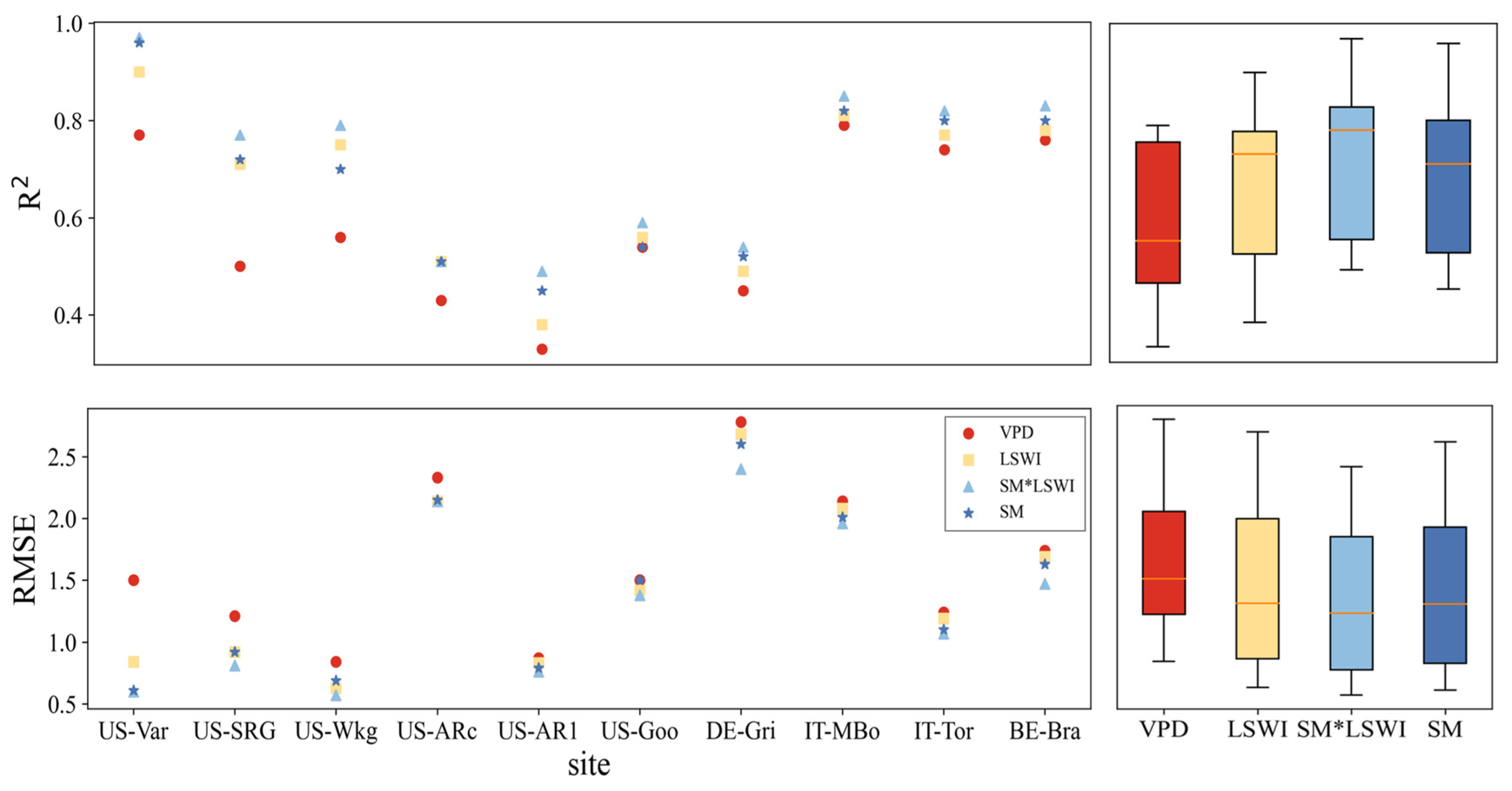
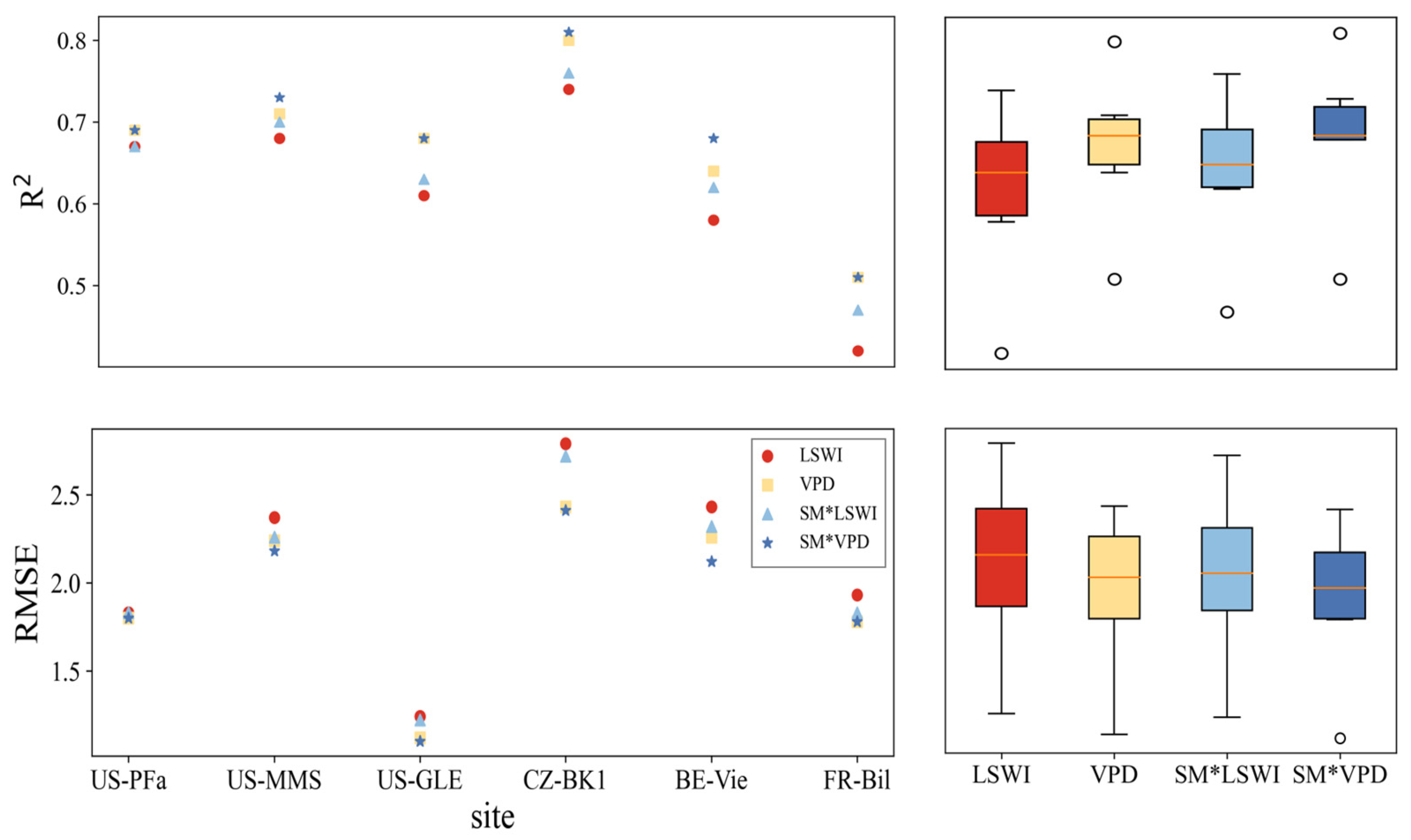
3.1. Performances of Original Models
3.2. Evaluation of Improved Models
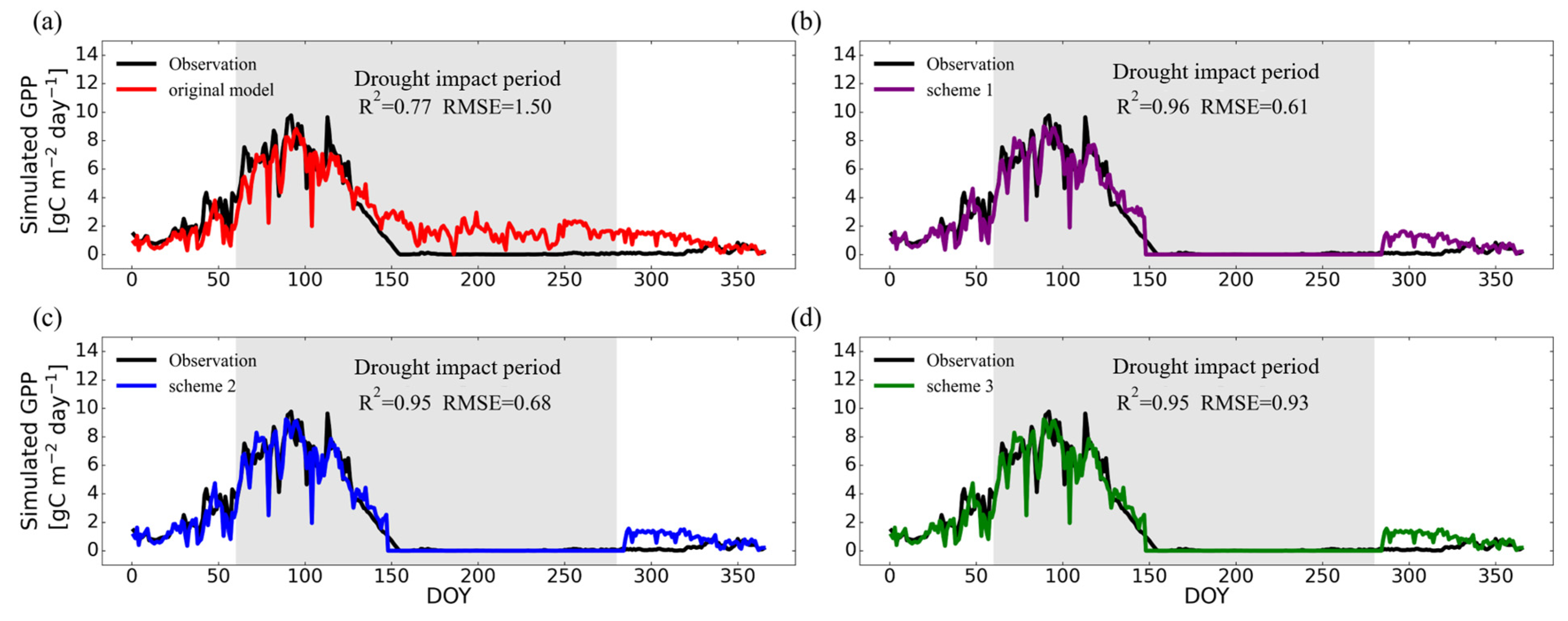
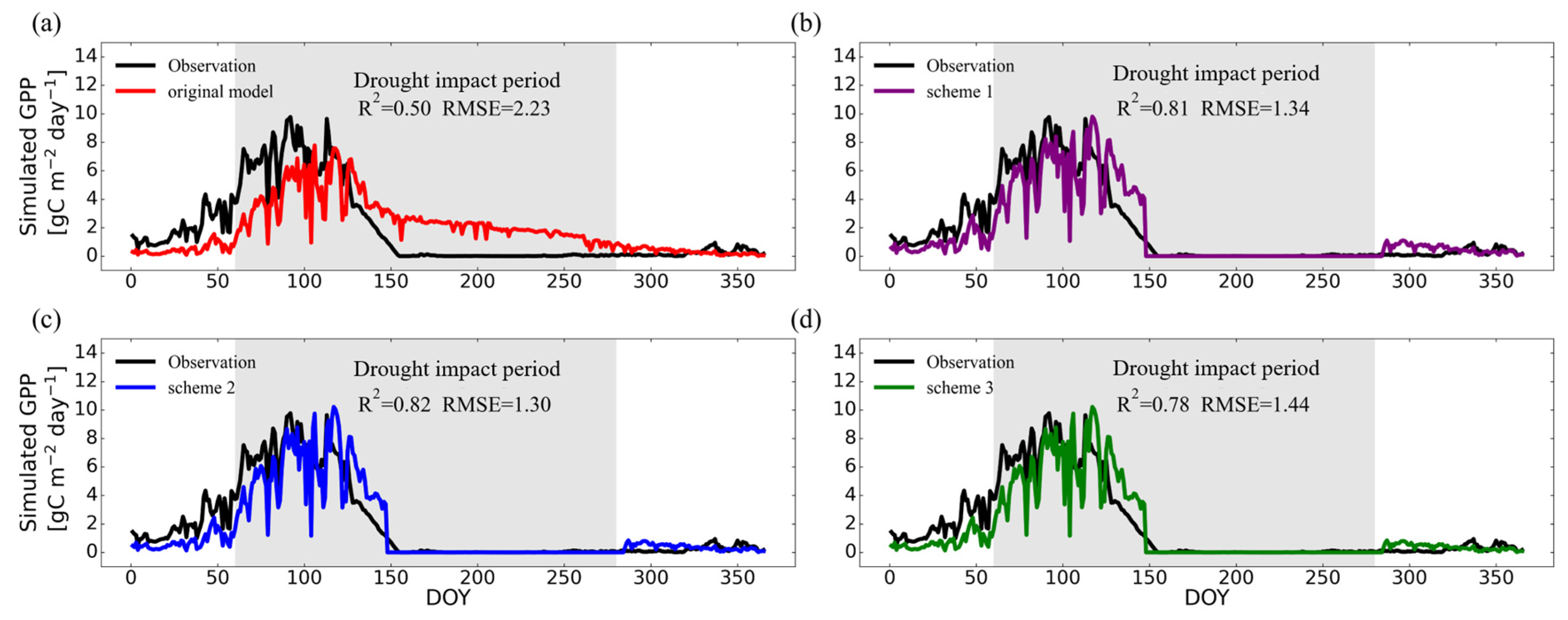
3.2.1. Temporal Variations on Typical Site
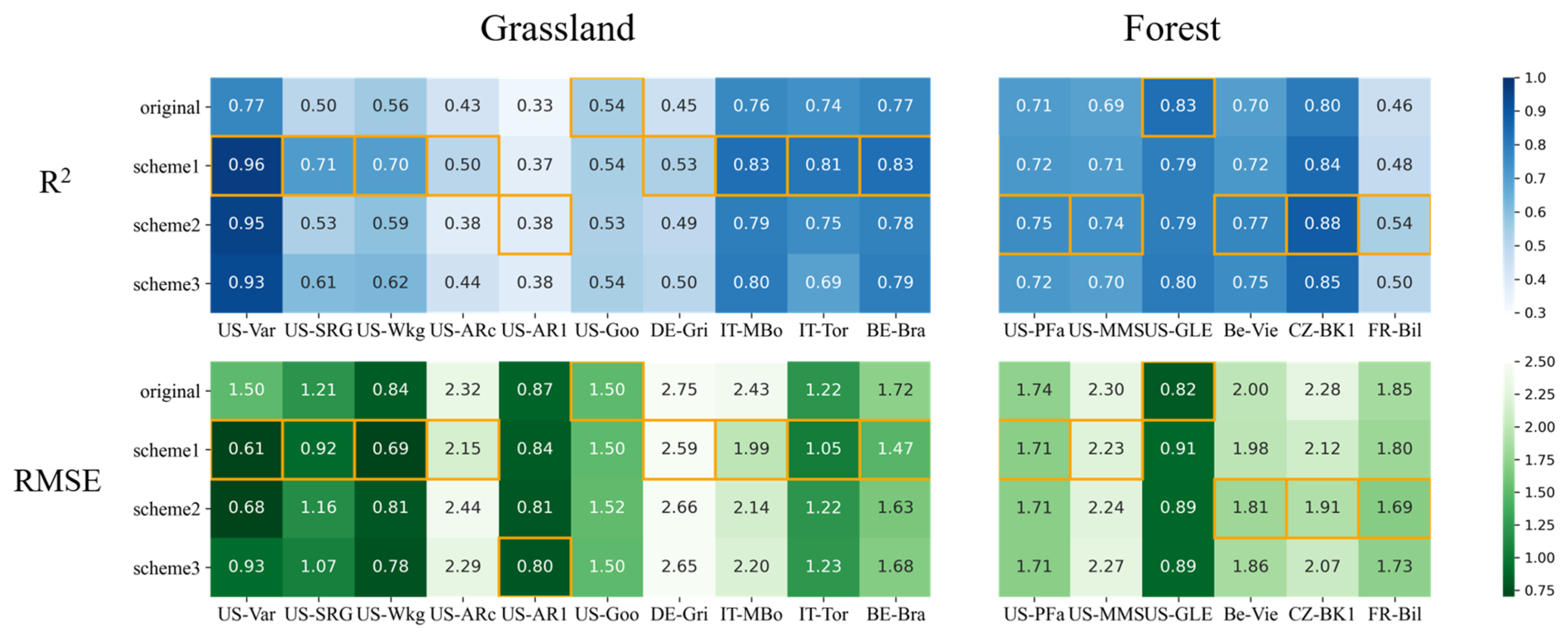
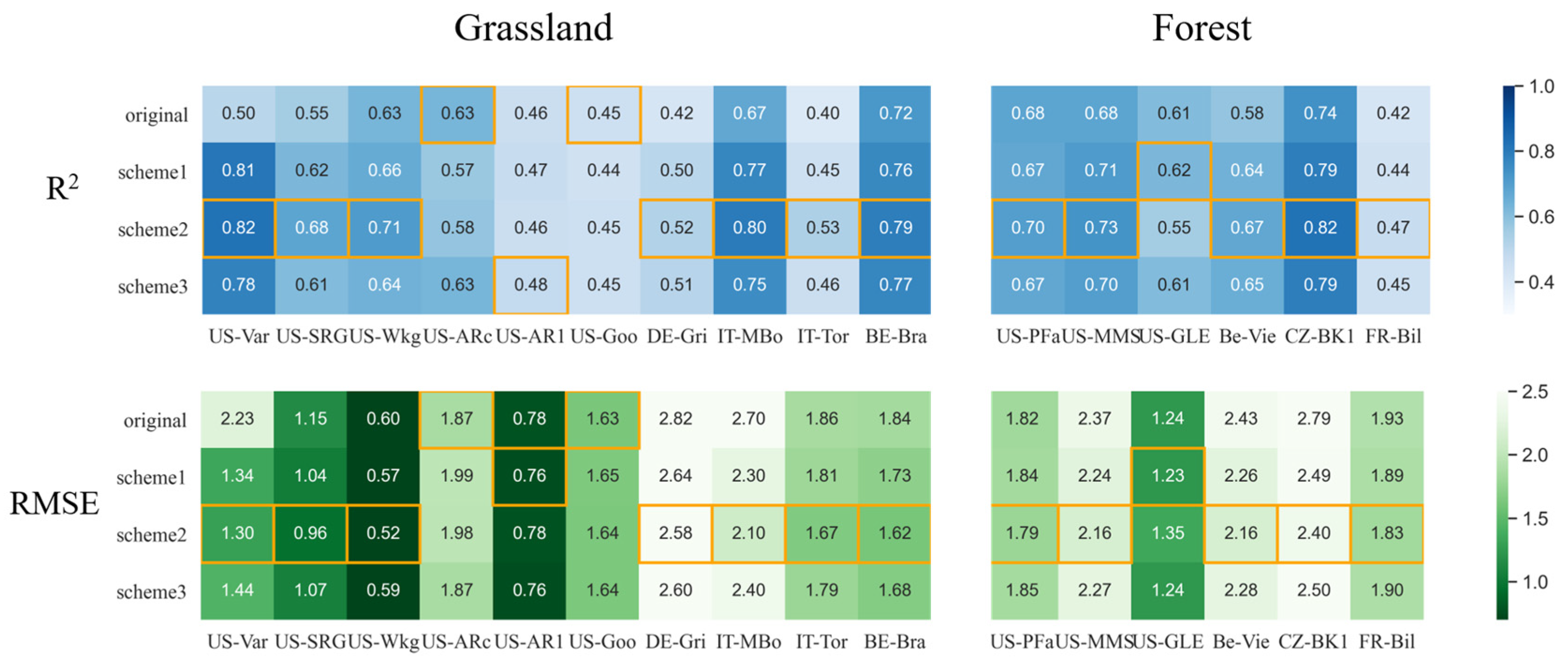
3.2.2. Improvements for Grass-Type Sites
3.2.3. Improvements for Forest Sites
3.2.4. Improvements for Non-Drought Periods
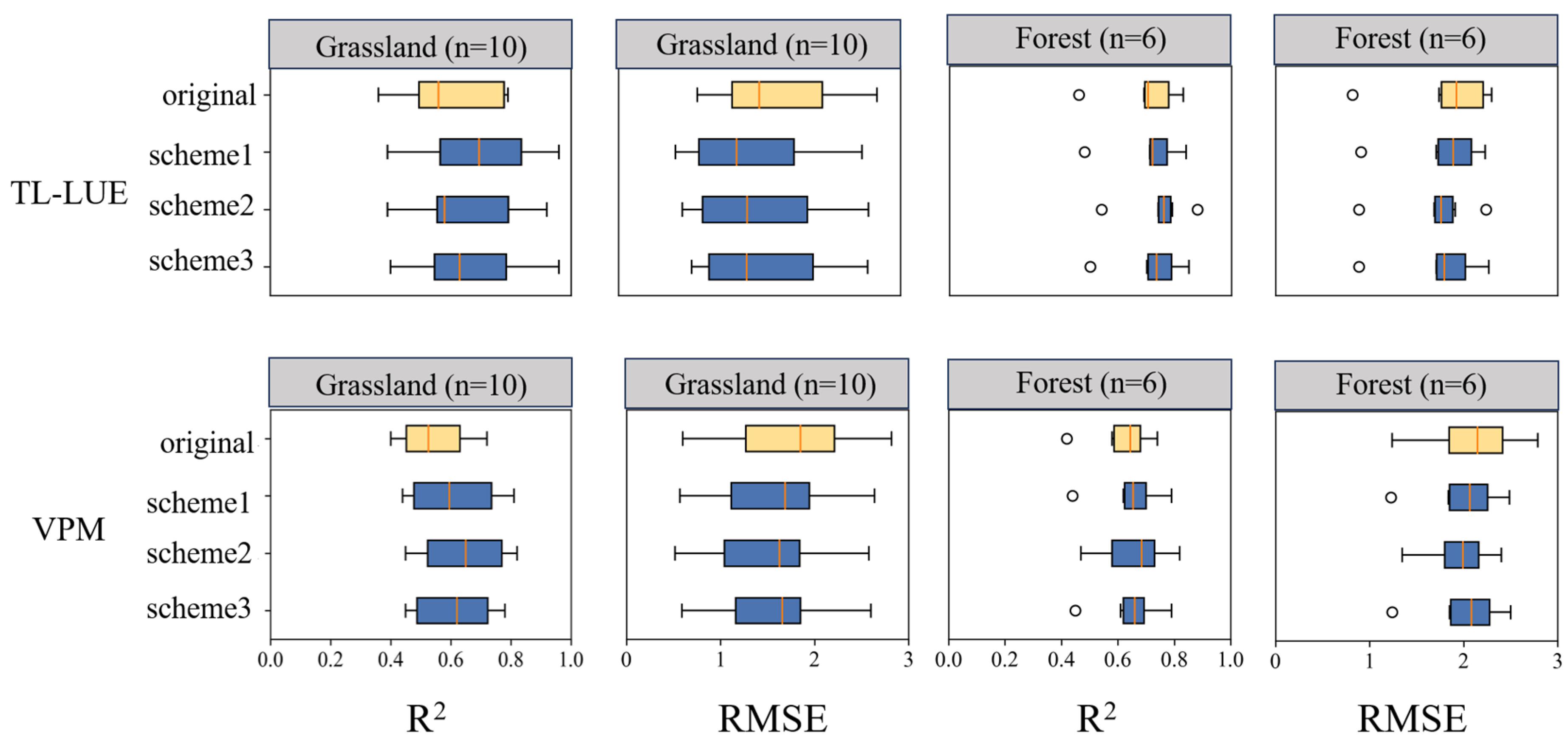
3.3. Improvement Comparison Between TL-LUE and VPM
4. Discussion
4.1. The Added Value of Incorporating Soil Moisture Information
4.2. Implications for Future LUE Model Development
4.3. Biodiversity Recovery, Ecosystem Functioning, and Sustainability
4.4. Uncertainties
5. Conclusions
- (1)
- At most grass-type sites, VPM outperformed the TL-LUE model during drought periods, which is associated with the fact that LSWI responded better to drought than VPD at grass-type ecosystems. In contrast, at forest ecosystem sites, the TL-LUE model performed better, with VPD being more representative of water stress in forest ecosystems during drought periods.
- (2)
- At grass-type sites, incorporating SM information by using a sigmoid-type SM correction function f(SM) led to clear improvements for both LUE models, in particular with the scheme multiplying f(SM) with LSWI. The TL-LUE model exhibited a maximum increase in R2 of 0.27 (+54%) and a maximum decrease in RMSE of 0.40 gCm−2 day−1 (−33%). VPM showed improvements in R2 as large as 0.32 (+64%) with a decrease in RMSE at a maximum of 0.93 gCm−2 day−1 (−41%).
- (3)
- In comparison, for forest ecosystems, it was more challenging to improve LUE models with SM under water stresses (for TL-LUE: ΔR2 = −0.04~0.08, ΔRMSE = +0.07~−0.37 gCm−2 day−1; and for VPM, ΔR2 = 0.01~0.08, ΔRMSE = −0.01~−0.39 gCm−2 day−1), since forests are less sensitive to surface SM variation. In addition, in contrast to grass-type ecosystems, using VPD as a moisture indicator in VPM can better improve forest GPP simulations under water stresses than using LSWI.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scheme 1 | Scheme 2 | Scheme 3 | |
|---|---|---|---|
| Strengths | Best performance for grass-type ecosystems. Simple and effective for capturing surface soil moisture. | Best overall performance for both grasslands and forests. Performs well during non-drought and drought periods. | Balanced improvement across ecosystems. Effective in grasslands under certain conditions. |
| Weakness | Less effective for forests (minimal improvement at forest sites). Limited in representing leaf moisture. | High data requirements (e.g., soil moisture and LSWI data). Slightly lower performance for grasslands at some sites. | Inferior to Scheme 1 and Scheme 2 at most sites. Less significant improvements during drought periods. |
| Opportunities | Can be widely applied in drought-prone grasslands. Easily extendable with more surface soil moisture data. | Promising for ecosystem-level GPP modeling in diverse regions. Can be further improved with advanced data fusion. | Useful for regions with mixed water stress impacts. Potential to refine for grassland ecosystems. |
| Threats | Performance highly dependent on surface soil moisture data quality. May not generalize to forest ecosystems. | Requires advanced remote sensing for LSWI and soil moisture integration. High computational cost for broader applications. | May fail to outperform simpler models. Less adaptable to varying climatic conditions. |
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, X.; Xiao, J. A Global, 0.05-Degree Product of Solar-Induced Chlorophyll Fluorescence Derived from OCO-2, MODIS, and Reanalysis Data. Remote Sens. 2019, 11, 517. [Google Scholar] [CrossRef]
- He, B.; Liu, J.; Guo, L.; Wu, X.; Xie, X.; Zhang, Y.; Chen, C.; Zhong, Z.; Chen, Z. Recovery of Ecosystem Carbon and Energy Fluxes From the 2003 Drought in Europe and the 2012 Drought in the United States. Geophys. Res. Lett. 2018, 45, 4879–4888. [Google Scholar] [CrossRef]
- Lv, Y.; Liu, J.; He, W.; Zhou, Y.; Tu Nguyen, N.; Bi, W.; Wei, X.; Chen, H. How Well Do Light-Use Efficiency Models Capture Large-Scale Drought Impacts on Vegetation Productivity Compared with Data-Driven Estimates? Ecol. Indic. 2023, 146, 109739. [Google Scholar] [CrossRef]
- Stocker, B.D.; Zscheischler, J.; Keenan, T.F.; Prentice, I.C.; Seneviratne, S.I.; Peñuelas, J. Drought Impacts on Terrestrial Primary Production Underestimated by Satellite Monitoring. Nat. Geosci. 2019, 12, 264–270. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, L.; Xiao, J.; Yuan, W.; Yan, M.; Li, T.; Zhang, Z. Sources of Uncertainty in Gross Primary Productivity Simulated by Light Use Efficiency Models: Model Structure, Parameters, Input Data, and Spatial Resolution. Agric. For. Meteorol. 2018, 263, 242–257. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-Wide Reduction in Primary Productivity Caused by the Heat and Drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef]
- Ghil, M.; Yiou, P.; Hallegatte, S.; Malamud, B.D.; Naveau, P.; Soloviev, A.A.; Friederichs, P.; Keilis-Borok, V.I.; Kondrashov, D.; Kossobokov, V.; et al. Extreme Events: Dynamics, Statistics and Prediction. Nonlinear Process. Geophys. 2011, 18, 295–350. [Google Scholar] [CrossRef]
- Yuan, M.; Zhu, Q.; Zhang, J.; Liu, J.; Chen, H.; Peng, C.; Li, P.; Li, M.; Wang, M.; Zhao, P. Global response of terrestrial gross primary productivity to climate extremes. Sci. Total Environ. 2021, 750, 142337. [Google Scholar] [CrossRef]
- Liu, S.; Chadwick, O.A.; Roberts, D.A.; Still, C.J. Relationships between GPP, satellite measures of greenness and canopy water content with soil moisture in Mediterranean-Climate Grassland and Oak Savanna. Appl. Environ. Soil Sci. 2011, 2011, 839028. [Google Scholar] [CrossRef]
- Valjarević, A. GIS-Based Methods for Identifying River Networks Types and Changing River Basins. Water Resour. Manag. 2024, 38, 5323–5341. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, X.; Zhou, S.; Ciais, P.; McCarthy, H.; Luo, Y. Canopy and physiological controls of GPP during drought and heat wave. Geophys. Res. Lett. 2016, 43, 3325–3333. [Google Scholar] [CrossRef]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J.; et al. Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef]
- Ji, L.; Peters, A.J. Assessing vegetation response to drought in the northern Great Plains using vegetation and drought indices. Remote Sens. Environ. 2003, 87, 85–98. [Google Scholar] [CrossRef]
- Monteith, J.L. Solar Radiation and Productivity in Tropical Ecosystems. J. Appl. Ecol. 1972, 9, 747. [Google Scholar] [CrossRef]
- Dong, J.; Xiao, X.; Wagle, P.; Zhang, G.; Zhou, Y.; Jin, C.; Torn, M.S.; Meyers, T.P.; Suyker, A.E.; Wang, J.; et al. Comparison of Four EVI-Based Models for Estimating Gross Primary Production of Maize and Soybean Croplands and Tallgrass Prairie under Severe Drought. Remote Sens. Environ. 2015, 162, 154–168. [Google Scholar] [CrossRef]
- Wu, C.; Chen, J.M.; Desai, A.R.; Hollinger, D.Y.; Arain, M.A.; Margolis, H.A.; Gough, C.M.; Staebler, R.M. Remote Sensing of Canopy Light Use Efficiency in Temperate and Boreal Forests of North America Using MODIS Imagery. Remote Sens. Environ. 2012, 118, 60–72. [Google Scholar] [CrossRef]
- Zhao, M.; Heinsch, F.A.; Nemani, R.R.; Running, S.W. Improvements of the MODIS Terrestrial Gross and Net Primary Production Global Data Set. Remote Sens. Environ. 2005, 95, 164–176. [Google Scholar] [CrossRef]
- He, W.; Ju, W.; Jiang, F.; Parazoo, N.; Gentine, P.; Wu, X.; Zhang, C.; Zhu, J.; Viovy, N.; Jain, A.K.; et al. Peak Growing Season Patterns and Climate Extremes-Driven Responses of Gross Primary Production Estimated by Satellite and Process Based Models over North America. Agric. For. Meteorol. 2021, 298–299, 108292. [Google Scholar] [CrossRef]
- Wei, X.; He, W.; Zhou, Y.; Cheng, N.; Xiao, J.; Bi, W.; Liu, Y.; Sun, S.; Ju, W. Increased Sensitivity of Global Vegetation Productivity to Drought Over the Recent Three Decades. J. Geophys. Res. Atmos. 2023, 128, e2022JD037504. [Google Scholar] [CrossRef]
- Stocker, B.D.; Zscheischler, J.; Keenan, T.F.; Prentice, I.C.; Peñuelas, J.; Seneviratne, S.I. Quantifying Soil Moisture Impacts on Light Use Efficiency across Biomes. New Phytol. 2018, 218, 1430–1449. [Google Scholar] [CrossRef]
- Green, J.K.; Seneviratne, S.I.; Berg, A.M.; Findell, K.L.; Hagemann, S.; Lawrence, D.M.; Gentine, P. Large Influence of Soil Moisture on Long-Term Terrestrial Carbon Uptake. Nature 2019, 565, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Feng, X.; Fu, B. Changes in Global Terrestrial Ecosystem Water Use Efficiency Are Closely Related to Soil Moisture. Sci. Total Environ. 2020, 698, 134165. [Google Scholar] [CrossRef] [PubMed]
- Verhoef, A.; Egea, G. Modeling Plant Transpiration under Limited Soil Water: Comparison of Different Plant and Soil Hydraulic Parameterizations and Preliminary Implications for Their Use in Land Surface Models. Agric. For. Meteorol. 2014, 191, 22–32. [Google Scholar] [CrossRef]
- Ju, W.; Chen, J.M.; Black, T.A.; Barr, A.G.; Liu, J.; Chen, B. Modelling Multi-Year Coupled Carbon and Water Fluxes in a Boreal Aspen Forest. Agric. For. Meteorol. 2006, 140, 136–151. [Google Scholar] [CrossRef]
- Yan, H.; Wang, S.; Huete, A.; Shugart, H.H. Effects of Light Component and Water Stress on Photosynthesis of Amazon Rainforests During the 2015/2016 El Niño Drought. J. Geophys. Res. Biogeosci. 2019, 124, 1574–1590. [Google Scholar] [CrossRef]
- Yuan, W.; Liu, S.; Zhou, G.; Zhou, G.; Tieszen, L.L.; Baldocchi, D.; Bernhofer, C.; Gholz, H.; Goldstein, A.H.; Goulden, M.L.; et al. Deriving a Light Use Efficiency Model from Eddy Covariance Flux Data for Predicting Daily Gross Primary Production across Biomes. Agric. For. Meteorol. 2007, 143, 189–207. [Google Scholar] [CrossRef]
- Jones, L.A.; Kimball, J.S.; Reichle, R.H.; Madani, N.; Glassy, J.; Ardizzone, J.V.; Colliander, A.; Cleverly, J.; Desai, A.R.; Eamus, D.; et al. The SMAP Level 4 Carbon Product for Monitoring Ecosystem Land–Atmosphere CO2Exchange. IEEE Trans. Geosci. Remote Sens. 2017, 55, 6517–6532. [Google Scholar] [CrossRef]
- Madani, N.; Parazoo, N.C.; Kimball, J.S.; Ballantyne, A.P.; Reichle, R.H.; Maneta, M.; Saatchi, S.; Palmer, P.I.; Liu, Z.; Tagesson, T. Recent Amplified Global Gross Primary Productivity Due to Temperature Increase Is Offset by Reduced Productivity Due to Water Constraints. AGU Adv. 2020, 1, e2020AV000180. [Google Scholar] [CrossRef]
- He, M.; Kimball, J.S.; Running, S.; Ballantyne, A.; Guan, K.; Huemmrich, F. Satellite Detection of Soil Moisture Related Water Stress Impacts on Ecosystem Productivity Using the MODIS-Based Photochemical Reflectance Index. Remote Sens. Environ. 2016, 186, 173–183. [Google Scholar] [CrossRef]
- Wang, S.; Ibrom, A.; Bauer-Gottwein, P.; Garcia, M. Incorporating Diffuse Radiation into a Light Use Efficiency and Evapotranspiration Model: An 11-Year Study in a High Latitude Deciduous Forest. Agric. For. Meteorol. 2018, 248, 479–493. [Google Scholar] [CrossRef]
- He, M.; Ju, W.; Zhou, Y.; Chen, J.; He, H.; Wang, S.; Wang, H.; Guan, D.; Yan, J.; Li, Y.; et al. Development of a Two-Leaf Light Use Efficiency Model for Improving the Calculation of Terrestrial Gross Primary Productivity. Agric. For. Meteorol. 2013, 173, 28–39. [Google Scholar] [CrossRef]
- Heiskanen, J.; Brümmer, C.; Buchmann, N.; Calfapietra, C.; Chen, H.; Gielen, B.; Gkritzalis, T.; Hammer, S.; Hartman, S.; Herbst, M.; et al. The Integrated Carbon Observation System in Europe. Bull. Am. Meteorol. Soc. 2022, 103, E855–E872. [Google Scholar] [CrossRef]
- Li, J.; Ju, W.; He, W.; Wang, H.; Zhou, Y.; Xu, M. An Algorithm Differentiating Sunlit and Shaded Leaves for Improving Canopy Conductance and Vapotranspiration Estimates. J. Geophys. Res. Biogeosci. 2019, 124, 807–824. [Google Scholar] [CrossRef]
- De Pury, D.G.G.; Farquhar, G.D. Simple scaling of photosynthesis from leaves to canopies without the errors of big-leaf models. Plant Cell Environ. 1997, 20, 537–557. [Google Scholar] [CrossRef]
- Chen, J.; Liu, J.; Cihlar, J.; Goulden, M. Daily canopy photosynthesis model through temporal and spatial scaling for remote sensing applications. Ecol. Model. 1999, 124, 99–119. [Google Scholar] [CrossRef]
- Friend, A.D. Modelling canopy CO2 fluxes: Are ‘big-leaf’ simplifications justified. Glob. Ecol. Biogeogr. 2001, 10, 603–619. [Google Scholar] [CrossRef]
- Bi, W.; He, W.; Zhou, Y.; Ju, W.; Liu, Y.; Liu, Y.; Zhang, X.; Wei, X.; Cheng, N. A Global 0.05° Dataset for Gross Primary Production of Sunlit and Shaded Vegetation Canopies from 1992 to 2020. Sci. Data 2022, 9, 213. [Google Scholar] [CrossRef]
- Keenan, T.F.; Prentice, I.C.; Canadell, J.G.; Williams, C.A.; Wang, H.; Raupach, M.; Collatz, G.J. Recent Pause in the Growth Rate of Atmospheric CO2 Due to Enhanced Terrestrial Carbon Uptake. Nat. Commun. 2016, 7, 13428. [Google Scholar] [CrossRef]
- Xiao, X.; Hollinger, D.; Aber, J.; Goltz, M.; Davidson, E.A.; Zhang, Q.; Moore, B. Satellite-Based Modeling of Gross Primary Production in an Evergreen Needleleaf Forest. Remote Sens. Environ. 2004, 89, 519–534. [Google Scholar] [CrossRef]
- Li, J.; Xiao, Z. Evaluation of the Version 5.0 Global Land Surface Satellite (GLASS) Leaf Area Index Product Derived from MODIS Data. Int. J. Remote Sens. 2020, 41, 9140–9160. [Google Scholar] [CrossRef]
- Vermote, E. MOD09A1 MODIS/Terra Surface Reflectance 8-Day L3 Global 500m SIN Grid V006. NASA EOSDIS LP DAAC. 2015. Available online: https://salsa.umd.edu/files/MOD09_UserGuide_v1.4.pdf (accessed on 19 January 2025).
- Zhang, Y.; Schaap, M.G.; Wei, Z. Development of hierarchical ensemble model and estimates of soil water retention with global coverage. Geophys. Res. Lett. 2020, 47, e2020GL088819. [Google Scholar] [CrossRef]
- Zhang, Y.; Schaap, M.G.; Zha, Y. A High-Resolution Global Map of Soil Hydraulic Properties Produced by a Hierarchical Parameterization of a Physically Based Water Retention Model. Water Resour. Res. 2018, 54, 9774–9790. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, C.; Sun, G.; Band, L.E.; Noormets, A.; Zhang, Q. Understanding Moisture Stress on Light Use Efficiency across Terrestrial Ecosystems Based on Global Flux and Remote-Sensing Data. J. Geophys. Res. Biogeosci. 2015, 120, 2053–2066. [Google Scholar] [CrossRef]
- Canadell, J.; Jackson, R.B.; Ehleringer, J.B.; Mooney, H.A.; Sala, O.E.; Schulze, E.-D. Maximum Rooting Depth of Vegetation Types at the Global Scale. Oecologia 1996, 108, 583–595. [Google Scholar] [CrossRef]
- Bayat, B.; van der Tol, C.; Yang, P.; Verhoef, W. Extending the SCOPE model to combine optical reflectance and soil moisture observations for remote sensing of ecosystem functioning under water stress conditions. Remote Sens. Environ. 2019, 221, 286–301. [Google Scholar] [CrossRef]
- Schlaepfer, D.R.; Bradford, J.B.; Lauenroth, W.K.; Munson, S.M.; Tietjen, B.; Hall, S.A.; Wilson, S.D.; Duniway, M.C.; Jia, G.; Pyke, D.A.; et al. Climate Change Reduces Extent of Temperate Drylands and Intensifies Drought in Deep Soils. Nat. Commun. 2017, 8, 14196. [Google Scholar] [CrossRef]
- Pei, Y.; Dong, J.; Zhang, Y.; Yuan, W.; Doughty, R.; Yang, J.; Zhou, D.; Zhang, L.; Xiao, X. Evolution of Light Use Efficiency Models: Improvement, Uncertainties, and Implications. Agric. For. Meteorol. 2022, 317, 108905. [Google Scholar] [CrossRef]
- Guan, X.; Chen, J.M.; Shen, H.; Xie, X.; Tan, J. Comparison of Big-Leaf and Two-Leaf Light Use Efficiency Models for GPP Simulation after Considering a Radiation Scalar. Agric. For. Meteorol. 2022, 313, 108761. [Google Scholar] [CrossRef]
- Dechant, B.; Ryu, Y.; Badgley, G.; Köhler, P.; Rascher, U.; Migliavacca, M.; Zhang, Y.; Tagliabue, G.; Guan, K.; Rossini, M.; et al. NIRVP: A Robust Structural Proxy for Sun-Induced Chlorophyll Fluorescence and Photosynthesis across Scales. Remote Sens. Environ. 2022, 268, 112763. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C.; Döll, M.; Fietz, H.-J.; Bach, T.; Kozel, U.; Meier, D.; Rahmsdorf, U. Photosynthetic Activity, Chloroplast Ultrastructure, and Leaf Characteristics of High-Light and Low-Light Plants and of Sun and Shade Leaves. Photosynth. Res. 1981, 2, 115–141. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, L.; Zhou, T.; Zhang, X. Long-term changes in the effect of drought stress on ecosystems across global drylands. Sci. China Earth Sci. 2023, 66, 146–160. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, L.; Tang, Z.; Wu, S. A synthesis of ecosystem aboveground productivity and its process variables under simulated drought stress. J. Ecol. 2019, 107, 2519–2531. [Google Scholar] [CrossRef]
- Sanaullah, M.; Rumpel, C.; Charrier, X.; Chabbi, A. How does drought stress influence the decomposition of plant litter with contrasting quality in a grassland ecosystem? Plant Soil 2012, 352, 277–288. [Google Scholar] [CrossRef]
- Yao, Y.; Liu, Y.; Zhou, S.; Song, J.; Fu, B. Soil moisture determines the recovery time of ecosystems from drought. Glob. Change Biol. 2023, 29, 3562–3574. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Guan, J.; Zheng, J.; Wang, Y.; Han, W.; Liu, Y. Cumulative effects of drought have an impact on net primary productivity stability in Central Asian grasslands. J. Environ. Manag. 2023, 344, 118734. [Google Scholar] [CrossRef]

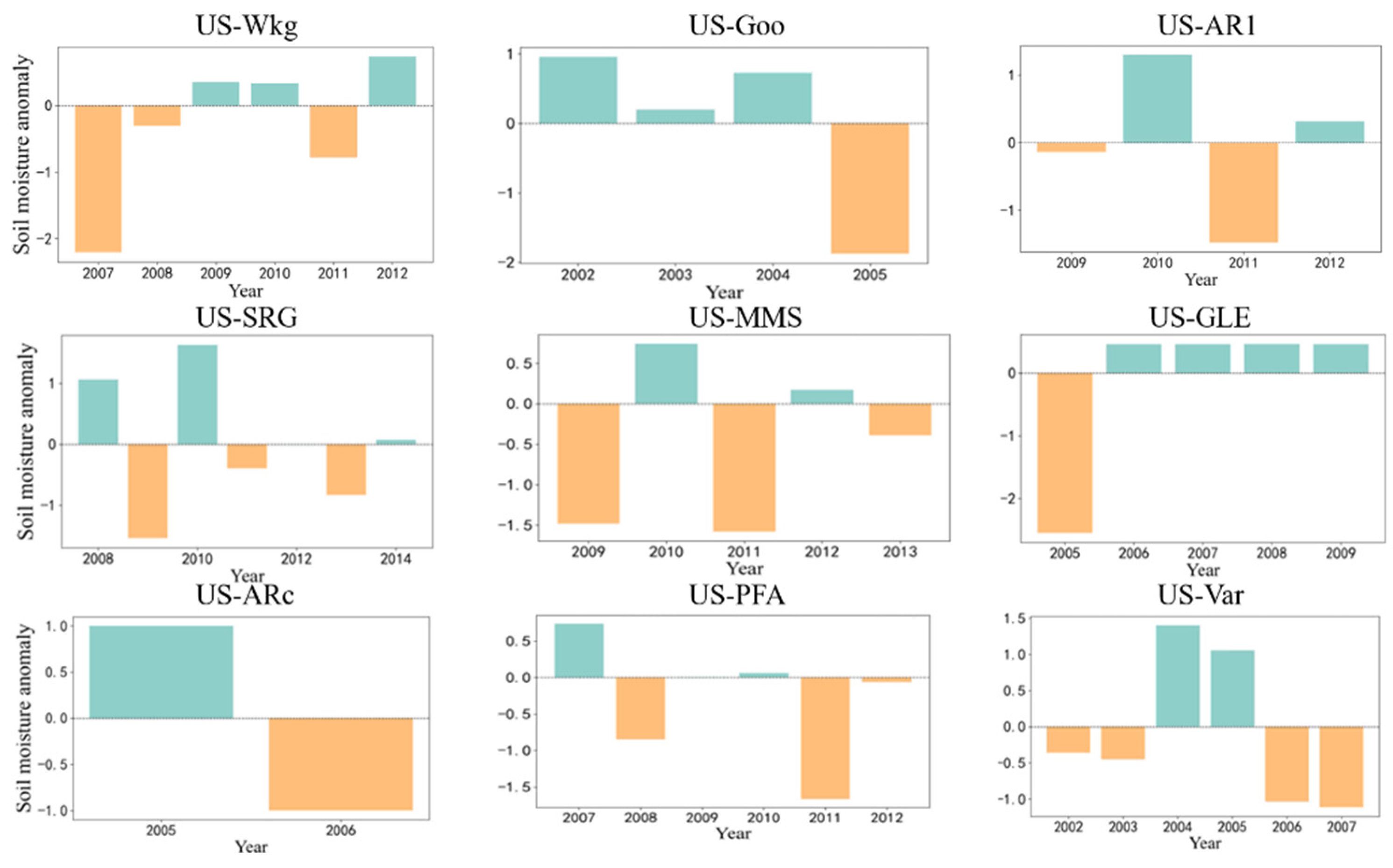

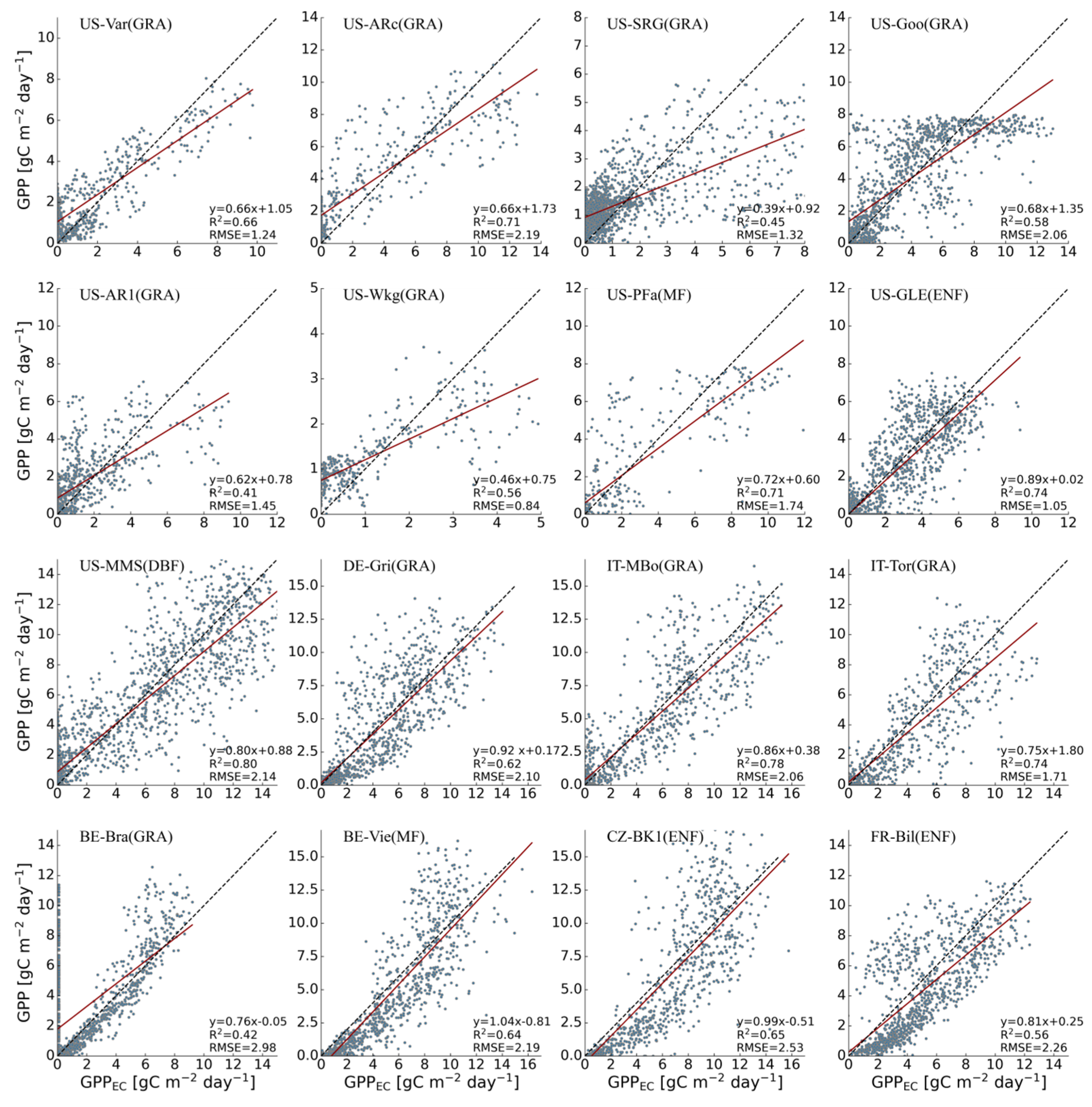
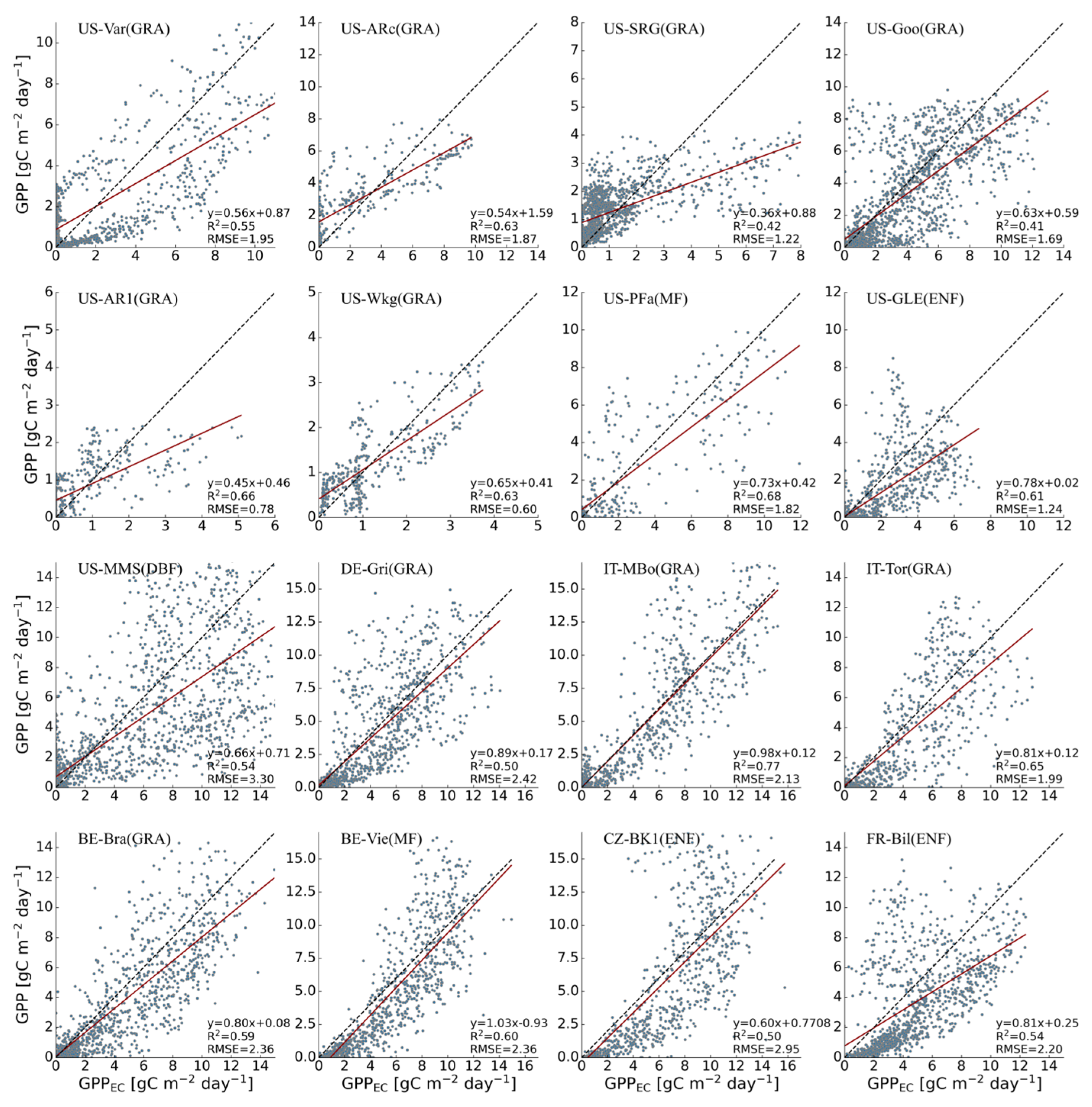
| Site-ID | Vegetation Type | Lon | Lat | WC | WP | Drought Year |
|---|---|---|---|---|---|---|
| US-Goo | GRA | 89.87° W | 34.25° N | 0.28 | 0.11 | 2005 |
| US-ARc | GRA | 98.04° W | 35.55° N | 0.29 | 0.13 | 2006 |
| US-SRG | GRA | 110.83° W | 31.79° N | 0.24 | 0.10 | 2009 |
| US-Var | GRA | 120.95° W | 38.41° N | 0.29 | 0.12 | 2007 |
| US-Wkg | GRA | 109.94° W | 31.74° N | 0.25 | 0.11 | 2007 |
| US-AR1 | GRA | 99.42° W | 36.43° N | 0.27 | 0.12 | 2011 |
| DE-Gri | GRA | 13.51° E | 50.95° N | 0.26 | 0.11 | 2018 |
| IT-MBo | GRA | 11.05° E | 46.01° N | 0.27 | 0.14 | 2019 |
| IT-Tor | GRA | 7.58° E | 45.84° N | 0.29 | 0.12 | 2017 |
| BE-Bra | GRA | 4.52° E | 51.30° N | 0.25 | 0.14 | 2020 |
| US-MMS | DBF | 86.41° W | 39.32° N | 0.32 | 0.10 | 2011 |
| US-PFa | MF | 90.27° W | 45.95° N | 0.26 | 0.09 | 2011 |
| BE-Vie | MF | 5.99° E | 50.30° N | 0.28 | 0.14 | 2013 |
| US-GLE | ENF | 106.24° W | 41.36° N | 0.31 | 0.12 | 2005 |
| CZ-BK1 | ENF | 18.54° E | 49.50° N | 0.28 | 0.15 | 2018 |
| FR-Bil | ENF | 0.96° W | 44.49° N | 0.30 | 0.10 | 2017 |
| Site | R(GPPobs-LSWI) | R(GPPobs-VPD) | R(GPPobs-SM) |
|---|---|---|---|
| US-Var | 0.88 | −0.39 | 0.40 |
| US-SRG | 0.61 | −0.02 | 0.21 |
| US-Wkg | 0.63 | 0.11 | 0.29 |
| US-ARc | 0.84 | 0.17 | 0.28 |
| US-AR1 | 0.39 | 0.19 | 0.29 |
| US-Goo | 0.75 | 0.26 | 0.05 |
| FR-TOU | 0.78 | 0.14 | 0.45 |
| DE-Gri | 0.69 | 0.18 | 0.39 |
| IT-MBo | 0.72 | 0.24 | 0.44 |
| IT-Tor | 0.79 | 0.26 | 0.51 |
| BE-Bra | 0.63 | 0.11 | 0.32 |
| US-PFa | 0.14 | 0.64 | 0.18 |
| US-MMS | 0.86 | 0.57 | 0.36 |
| BE-Vie | 0.36 | 0.67 | 0.23 |
| US-GLE | 0.23 | 0.77 | 0.08 |
| CZ-BK1 | 0.14 | 0.82 | 0.28 |
| FR-Bil | 0.24 | 0.78 | 0.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, Y.; He, W.; Liu, J.; Chen, H. Integration of Soil Moisture Factor into Light-Use Efficiency Models Improves Modeling Impact of Water Stresses on Gross Primary Production. Forests 2025, 16, 297. https://doi.org/10.3390/f16020297
Lv Y, He W, Liu J, Chen H. Integration of Soil Moisture Factor into Light-Use Efficiency Models Improves Modeling Impact of Water Stresses on Gross Primary Production. Forests. 2025; 16(2):297. https://doi.org/10.3390/f16020297
Chicago/Turabian StyleLv, Yiming, Wei He, Jinxiu Liu, and Hui Chen. 2025. "Integration of Soil Moisture Factor into Light-Use Efficiency Models Improves Modeling Impact of Water Stresses on Gross Primary Production" Forests 16, no. 2: 297. https://doi.org/10.3390/f16020297
APA StyleLv, Y., He, W., Liu, J., & Chen, H. (2025). Integration of Soil Moisture Factor into Light-Use Efficiency Models Improves Modeling Impact of Water Stresses on Gross Primary Production. Forests, 16(2), 297. https://doi.org/10.3390/f16020297