Abstract
In recent decades, large-scale forest disturbances such as windthrow and bark beetle infestations have significantly impacted Earth’s carbon balance and forest ecosystems. This impact alters soil respiration (SR), along with decreased gross primary productivity. To investigate the impact of bark beetle (Ips typographus L.) infestations in mountain spruce forests on SR, we measured SR at sites infested by bark beetles and adjacent undisturbed stands in the Tatra National Park (Slovakia) during the vegetation period (May–September) in 2016–2017 five to six years after the initial bark beetle attack. The measurements were taken along an altitudinal gradient (1100–1400 m a.s.l.). The highest rates of SR were observed during the summer months in both years (from June to August). However, yearly SR from May to September at infested sites showed significantly higher rates than uninfested ones in both years. SR showed a decreasing pattern with elevation gain in 2016 at infested sites, but this pattern was not observed in 2017. This study provides important insights into the impact of bark beetle infestations on SR and emphasizes the need for further research on the long-term effects of forest disturbances on carbon cycling. It also underscores the importance of determining the effect of different components of SR on the changed environment due to bark beetle attacks on mountain spruce forests.
1. Introduction
It is estimated that global soil carbon stock represents 1700 Gt C [1] and the contribution of forest soil ecosystems, as stated by [2], equals about half of it (861 Gt C). Top-meter soil, live biomass, deadwood, and litter each contain a percentage of the total forest C stock: 44%, 42%, 8%, and 5%, respectively. It creates an enormous carbon pool with the potential to highly increase atmospheric CO2 concentration [3] after large-scale disturbances. Carbon emission from global soil is considered the second largest carbon flux after photosynthesis, releasing 78–98 Gt C yr−1 in total [4,5]. Global forest soil carbon flux is an important contributor to global soil respiration (SR) because forests cover 26% of the Earth‘s total land area [6]. Under non-disturbance conditions, C uptake in the forest exceeds C emissions from ecosystem respiration [7] unless forest disturbance changes the balance and the forest becomes a carbon source [8,9]. Then, SR as the main component of ecosystem respiration [10] changes its rate as well.
Under climate change primarily induced by rising CO2 atmospheric concentrations [11], forests in Europe have experienced extreme heat and drought [12]. As a result, bark beetles as poikilothermic organisms have altered their behavior. They increase population size, and have extra generations per year [13]. Also, shifts of bark beetles to higher latitudes [14] and altitudes [15] have been observed. As a result, large-scale forest diebacks throughout the northern hemisphere have been caused by the European spruce bark beetle (Ips typographus L.) [16,17,18] recently.
Initially, after host tree death, the biogeochemical and biogeophysical processes (leaf area index, evapotranspiration, productivity, land surface temperature) of forest stands are altered [19,20]. Gross primary productivity (GPP) decreases as a result of tree mortality [21,22] and a reduction in the leaf area index corresponds to a contraction of GPP [19]. Fast nitrogen and carbon reduction take place in upper mineral soils, but an accumulation of soil inorganic N occurs as a result of the diminished contribution of roots, mycorrhizae, and rhizodeposition [23] which can be used by successional plant uptake [24], in the form of ammonium or nitrate [25]. The decline of fine root density increases with the level of tree mortality [26] and root biomass decomposition is accompanied by mycorrhizal fungi decomposition [27]. All of these factors contribute to changes in SR rate [28]. The decrease in SR after a disturbance event is mainly due to the reduced input of autotrophic SR [29] and nutrient losses [30]. After some time, infested trees shed their needles [20] and according to [31] approximately after 100 days spruce trees start to defoliate. Subsequently, increased solar radiation input [32,33,34] accelerates the decomposition rate of the litter as a consequence of higher temperatures [35].
Temperature is considered the most important factor influencing global SR [5,36,37]. SR positively correlates with ambient temperature [38]. With changes in temperature within different altitudinal zones, SR declines with increasing elevation [39,40]. Similarly, higher solar radiation intake on infested sites leads to rising soil and air temperatures on plots with bark beetle-infested trees [33]. SR follows seasonal dynamics of soil temperature with water surplus throughout the year [41], as it depends on both factors, with soil temperature having a more dominant effect [42,43].
Plant physiological–phenological relationships also influence SR rates, particularly the autotrophic component [44] and the successional stage after forest disturbance [45], as SR is linked to changes in phenology and photosynthesis [46]. After severe disturbance events, areas may shift to non-tree-dominated environments, with changes in plant species composition from shade-tolerant to shade-intolerant species due to altered solar conditions [32]. These plant communities, primarily grasses and perennial weeds, show increased biomass production and dispersive ability [47,48]. Over time after bark beetle attacks, grasses and perennials increase their surface coverage, leading to a greater influence on SR [49]. In the study site, four years after a windstorm, almost 100% of the area was covered by grasses and perennials [50]. The biomass production of successional grasses can exceed that of the original forest stand by a significant margin [51]. SR is also affected by Net Primary Productivity (NPP) [52,53,54]. Given that our study is on a vertical gradient on a homogenous slope, differences in physiological–phenological factors should be observed between infested and uninfested sites. Therefore, changes in SR are influenced not only by the soil temperature relationship but also by other factors [44], such as plant physiological–phenological dynamics and NPP.
There are studies that focus on SR within a vertical gradient [43,55,56] and studies that examine SR after windthrow or bark beetle attacks [22,42,49,57]. However, these studies do not address how different elevation zones of mountain spruce forests respond to disturbance events in terms of SR. Thus, in this study, we measured soil CO2 efflux for two consecutive years and investigated if there is a significant difference in SR between infested sites and uninfested sites with living trees at different elevational zones during the vegetation period. Our first hypothesis is that SR under dead trees would be lower than under uninfested trees due to a decrease in autotrophic SR (tree roots). Secondly, we predict that SR will not significantly vary across different altitudes due to minimal microclimatic differences within the measured mountain slope. Additionally, we expect the highest SR rates during summer (June to August) due to increased heterotrophic and autotrophic SR stimulated by higher soil temperatures.
2. Materials and Methods
2.1. Study Area
Our study was conducted in protected, unmanaged areas of the Tatra National Park in Slovakia. The studied site is dominated by Norway spruce (Picea abies (L.) Karst.) with a few other coniferous tree species mentioned in a previous study [58]. Since 2011, the area has been affected by an attack from the European spruce bark beetle (Ips typographus L.). Spruce forests above 1300 m in elevation are considered natural old-growth, while those below this zone are classified as semi-natural spruce forests due to frequent wind damage, resulting in a higher proportion of successional trees [58]. We measured SR five to six years after the initial bark beetle attack because SR is one of the ecological parameters measured on five-year cycles at the LTER sites (long-term ecological research) [59].
The research area was situated on the boundary between undisturbed and bark beetle-infested areas, with an elevation gradient ranging from 1100 to 1400 m above sea level between Tatranská Lomnica (820 m above sea level) and Skalnaté pleso (1754 m above sea level). The undisturbed forest in the area ranged from 100 to 165 years old, with stocking density varying from 0.4 to 0.8. A windthrow event in 2014 at 1200 m above sea level caused a decrease in stocking density to 0.4 [58]. The understory vegetation primarily consisted of acidophilic plants, particularly bilberry (Vaccinium myrtillus). Infested plots were characterized by no living trees with already shaded needles, and the understory was represented by successional grasses and perennials fully covering the soil. More detailed information about the study area can be found in the mentioned study [58].
The annual precipitation at Tatranska Lomnica in 2016 and 2017 was 922 and 934 mm, respectively, with mean annual temperatures of 6.7 and 6.6 °C during those years. These two years were wetter and warmer than the long-term average (1911–1960) of 833 mm and 4.7 °C for mean annual precipitation and air temperature, respectively. The main soil type in the studied area is an Acidic Distric Cambisol [60].
2.2. Soil Respiration Measurements
Measurements were conducted for two consecutive years, in 2016 and 2017, during the vegetation period (May–September). At each elevation zone (1100, 1200, 1300, 1400 m a.s.l.), 15 points were established for the undisturbed plots and 15 points for bark beetle infested plots using plastic collars. These collars, 3 to 5 cm high with the same diameter as the chamber, were inserted into the soil to ensure that the next measurement would be conducted at the same point. The tops of the collars were kept at the same level as the soil surface to minimize their influence on microclimatic conditions.
Measurements were taken on a biweekly basis using manual portable PP Systems, model EGM-4 (PP Systems, Amesbury, MA, USA), to measure soil CO2 fluxes. In total, 4 plots (four elevation zones) × 2 subplots (disturbed and undisturbed forest) × 15 points per each subplot = 120 measurements were conducted per sampling day. Soil temperature at a depth of 2 cm, soil moisture, and H2O flux (evaporation) were measured simultaneously.
2.3. Statistical Analysis
First, the measured data were checked for outliers. Subsequently, the data were analyzed using a three-way repeated measures analysis of variance (ANOVA), with factors including vegetation, year, month, and altitude. However, one factor had to be fixed. The statistical analysis was conducted using the R programming language (version 4.1.2, R Core Team, Vienna, Austria). The data did not follow a normal distribution based on the Shapiro–Wilk test and Q–Q plots. Thus, the aligned rank transform (ART) tool [61] was applied for nonparametric repeated measures factorial ANOVA using Holm method adjustment for post hoc pairwise comparisons, because multiple comparisons were needed.
The relationship between SR and soil temperatures was modeled using an exponential model [62,63] (Equation (1)):
where SR represents SR in g CO2 m2 h−1, ST denotes the soil temperature in °C, and a and b are the model parameters that best fit the measured values.
SR = aebST
The relationship between SR and soil moisture was analyzed using a simple linear regression model [64] (Equation (2)):
where a and b are the model coefficients and SM represents soil moisture content in %v/v.
SR = a + bSM
3. Results
SR varied throughout the vegetation period at different altitudes and months in 2016 and 2017 (Figure 1). The mean SR during the vegetation period was highest at the elevation of 1200 m.a.s.l. at both infested and uninfested sites (Figure 1 and Figure 2). We observed that SR reached its peak in the summer (July or August) at each elevation zone. Almost every month and at each elevation zone, infested sites emitted more CO2 than uninfested ones, but in many cases it was insignificantly higher. The mean SR at different elevations between infested and uninfested plots was found to be insignificant at the elevation zone of 1400 m and 1300 m in 2016 and 2017, respectively (Figure 1 and Figure 2). A faster increase in SR at infested sites towards the peak of the growing season (July–August) was observed. We also noticed a slower decrease in SR towards the end of the growing season at infested sites compared to undisturbed sites in both years. Rates of SR differed between forest statuses within a year (Figure 1). In both years, infested sites showed significantly higher yearly average values than uninfested ones. The mean annual SR (during the vegetation period) in 2016 reached 0.625 ± 0.335 g CO2 m2 h−1 and 0.428 ± 0.248 g CO2 m2 h−1 at infested and uninfested sites, respectively. In 2017, SR rates were 0.576 ± 0.275 CO2 m2 h−1 and 0.438 ± 0.249 g CO2 m2 h−1 at infested and undisturbed sites, respectively. We observed no statistical difference in SR between the two subsequent vegetation periods. The critical p-value was set at 0.05 (α = 0.05).
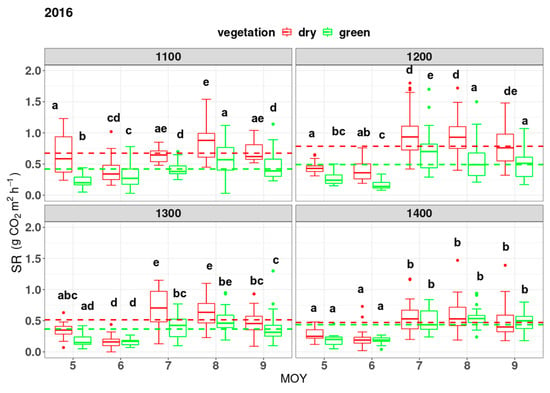
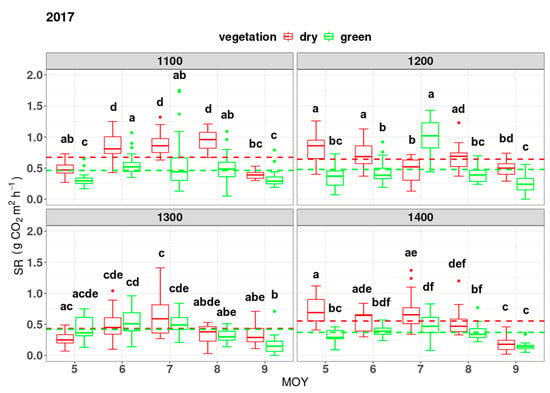
Figure 1.
Variation of soil respiration (SR) (y-axis) in g CO2 m2 h−1 within the same altitude in different months of the year (MOY) from May (5) to September (9) in both years. Letters display a significant difference with p < 0.05. Comparison is conducted within one box (elevation is in meters above sea level). Horizontal dashed lines represent the mean SR of the uninfested (green line) and infested forest sites (red line) for each elevation zone.
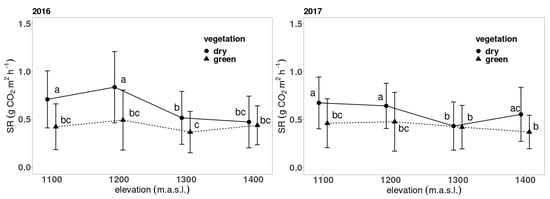
Figure 2.
Average soil respiration rates in 2016 and 2017 were compared at various altitudes and forest statuses. Soil respiration (SR) measurements were taken at different elevations in meters above sea level, with dry indicating infested forests and green representing uninfested spruce forests. Letters denote significant differences, with p < 0.05.
The monthly average fluxes from the soil surface under infested sites are higher in both years, although not always significantly (Figure 1). The average SR at undisturbed sites showed the same amount of carbon emitted throughout the vegetation period at each elevation zone, at a significant level (Figure 2). On the other hand, a higher variation of SR was observed under infested plots (Figure 2). Our results show a clear pattern of decrease or increase in soil efflux within the elevation gradient in 2017 at infested sites. However, no decline in SR with elevation was observed at undisturbed sites in both years, at a significance level of p < 0.05 (Figure 2). In 2016 at infested sites, we observed a significant decrease in SR with elevation gain.
SR in both years and types of forests did not show a high dependence on soil temperature and there were significant variations in 2016 between infected and undisturbed spruce forests (Figure 3). The disturbed forest exhibited a higher correlation between SR and temperature compared to the undisturbed forest. Additionally, the year 2016 shows a more rapid change in SR with changes in soil temperature (Figure 3). In 2017, a minimal difference in the dependence between SR and temperature was observed between disturbed and undisturbed forests. The correlation slope showed almost identical values. We also noticed a decrease in soil temperature between the highest and the lowest elevation zone in both years and forest conditions. The highest soil temperatures were measured during the summer months (from June to August) in both years and no correlation between soil moisture and respiration was observed. As optimal values of soil moisture were reached, it did not significantly impact SR rates. Additionally, we found a higher variance in moisture content at infested sites. The difference in moisture content between infected and uninfected sites was statistically significant. Disturbed sites covered with grasses and perennials had higher soil moisture content in both years (Table 1).
Figure 3.
Relationship between soil respiration and soil temperature. Relationship is modelled using Equation (1) (solid line). SR refers to soil respiration.

Table 1.
Comparison of soil moisture and temperature between dry (infested) and green (uninfested) spruce forest in different years. The critical p-value is α = 0.05. Mean and standard deviation (sd) are reported in degrees Celsius (°C) for soil temperature and as a percentage by volume (%v/v) for soil moisture. The symbols used to denote statistical significance are as follows: *** = p close to 0, * = p close to 0.01, ns = non-significant.
We observed significantly higher mean annual soil moisture and temperature at infested sites than at uninfested sites. The increase in soil temperature at infested sites within two years of the experiment was also observed. Similarly, significantly higher values for soil moisture were observed at infested sites. Some differences in soil moisture between the years were measured, but they were at the limit of significance (Table 1).
4. Discussion
In our experiment, we found that SR was significantly higher in the infested forest throughout both years. Changes in SR rate after disturbance events are not consistent throughout the published research papers [22,30,42,57,65,66,67]. In the girdling experiment in boreal Scots pine (Pinus sylvestris L.) forest, SR decreased by approximately 50% relative to ungirdled sites within one to two months [65], where forest mycorrhizae alone contribute to one-third of dissolved organic matter in forest soils, together with associated roots, contributing to 50% of dissolved organic matter [27]. However, a recent meta-analysis by [68] concluded that microbial, root, and mycorrhizal respiration contribute 57%, 28%, and 15%, respectively, to total SR. Soil fluxes decline as a consequence of altering key factors and nutrients [30] controlling SR rate.
Ref. [69] mentioned that up to 3 years after lodgepole pine (Pinus contorta Dougl. ex Loud.) infestation by mountain pine beetle, most needles remained on the trees. Therefore, no additional needle litter is added from dead pines to increase SR rate during this period of infestation. After a pulse of dead needles, SR almost fully recovered, lasting for up to 2 years and then followed by a decline again [22]. We suggest that needlefall and debris input from dying and dead spruce trees increased heterotrophic respiration, which compensates for the loss of autotrophic respiration. Since our study was conducted 5 to 6 years after the initial infestation by bark beetles, the results are quite similar to those of [22,49] with the difference that SR not only equaled undisturbed plots but exceeded them. This contradicts our initial hypothesis that SR will be higher at uninfested sites. Nonetheless, in our study, infested sites were covered by grasses and perennials with significant biomass productivity [51,70], which can increase autotrophic SR rates originating from the rhizosphere [71]. This phenomenon can also increase SR rates, in addition to hypothesized increased heterotrophic respiration due to higher mean soil temperatures [72]. The observed faster growth of SR towards the peak of the growing season (July and August) followed by a slower decrease in SR rates towards the end of the growing season at disturbed sites can be attributed to the gradual growth of perennials and grasses. This can be explained by the physiological–phenological development of a variety of plant species occupying research plots. Firstly, spring species start to grow (April–May), followed by summer species (May–June) and autumn species (June–August), creating a gradual and steady supply of autotrophic SR. Therefore, after disturbance, subsequent root biomass production of understory vegetation (grasses and perennials) [51,57,70] can account for increased SR within disturbed stands. Seven years after the disturbance, in terms of net primary productivity, successional understory vegetation acquires values that are only three times lower than forest stands before the disturbance event [73]. Nevertheless, developed understory vegetation combined with increased heterotrophic respiration caused by higher nutrient content [74,75] in the soil can compensate for severe tree death following a bark beetle attack.
Ref. [57] did not observe any significant changes in SR at a stand level from July to September over a period of 5 years between live lodgepole pine sites and bark beetle-infested sites. This phenomenon is attributed to surviving trees, understory vegetation, and the nutrient pulse from needlefall as noted by [74]. The decline in autotrophic respiration is offset by higher heterotrophic respiration induced by increased soil temperature, as discussed by [35,76]. Nonetheless, it is suggested that if mortality reaches 100%, total SR decreases to one-third of that in uninfested sites as stated by [57]. A similar pattern to that observed by [57] has been seen in fir-spruce forests by [42], attributed to root-respiring carbohydrates after tree death or a decrease in autotrophic respiration being replaced by heterotrophic respiration from dead roots and foliage. Dying roots and mycorrhizae release stored carbon for 2–3 years after disturbance, as reported by [77]. Additionally, an increase in soil temperature and soil moisture has been observed in infested ponderosa pine (Pinus ponderosa Laws.) forests by [33]. Ref. [49] confirmed that up to six years after disturbance, SR does not decrease at windthrow-disturbed sites. This is due to the substitution of decreased autotrophic SR with increasing heterotrophic SR supported by disturbance-induced alteration of soil temperature. In contrast, Ref. [30] concluded that the rapid decline of dissolved organic carbon, organic nitrogen, and phosphorus is accompanied by a decrease in SR after trees dieback, but after 4 years nutrients begin to recover due to litter mineralization. The post-disturbance chronosequence is an important factor influencing SR rate after bark beetle infestation. These varying results can be attributed to different mortality rates, gap size formation, and pre-existing understory vegetation.
As the temperature is considered the most important factor influencing global SR rates [5,36,37], with studies showing a positive correlation between SR and ambient temperature [71], we hypothesize that peak SR rates occur during the peak growing season in July and August, when temperatures are highest. This is likely due to the close relationship between SR and both air and soil temperature [78], as well as the increased contribution of heterotrophic respiration [79]. As temperatures decrease across different altitudinal zones [39,40], consequently, SR declines with increasing elevation [43,80]. Our results demonstrate a clear pattern of either decrease or increase in soil efflux along the elevation gradient in 2017 at infested sites, consistent with [56]. We observed a significant decrease in SR with elevation gain at infested sites in 2016, but no significant decline of SR with elevation had been observed at undisturbed sites in both years. This partially rejects our hypothesis that there will be no significant change in SR rates with increasing elevation.
The study of [75] conducted in the same vertical gradient shows that soil nutrient content increases with elevation and is statistically higher at infested sites. However, this finding contradicts the decreasing pattern of SR observed at infested sites in our study, as nutrient content plays a crucial role in supporting soil microorganisms that drive heterotrophic SR [30,81,82,83]. On the other hand, Ref. [75] only analyzed nutrient content for the year 2016. Nevertheless, we can suppose that increased nutrient availability of macroelements, increases autotrophic SR more than the heterotrophic component [84], but it can increase SR significantly because SR positively correlates with macroelement addition and increased temperature [81]. Also, like our study, Ref. [57] found no correlation between SR and moisture. We suppose that it can be supported by optimal moisture conditions during the studied period.
5. Conclusions
In this paper, we revealed that SR was significantly higher at sites with dead spruce trees in both years. SR did not significantly differ between the elevation zones in the undisturbed forest. On the other hand, a decrease in SR with elevation was observed at the sites with dead spruce trees in 2016. This highlights that different elevation zones respond differently to bark beetle attacks in terms of SR rates in mountain spruce forest. We suggest that it would be reasonable to partition SR into heterotrophic and autotrophic components to determine which component increases more due to increased soil temperature, moisture, and the productivity of successional vegetation. As our study was conducted only for two consecutive years, five to six years after the initial bark beetle attack, we propose long-term monitoring of SR at these sites to catch the dynamics of soil efflux in the later stages of succession.
Author Contributions
Conceptualization, P.F. and P.F.J.; methodology, P.F., P.F.J. and J.T.; software, P.F.J. and J.T.; validation, P.F. and P.F.J.; formal analysis, P.F.; investigation, P.F. and P.F.J.; resources, P.F.; data curation, P.F.J.; writing—original draft preparation, J.T. and M.K.; writing—review and editing, P.F.; visualization, J.T. and M.K.; supervision, P.F.J.; project administration, P.F.; funding acquisition, P.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Slovak Research and Development Agency, grant numbers APVV 17-0644 and APVV 21-0412.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank Ľuboš Slameň for assistance with field measurements. We also thank Richard Wylly for English revision and proofreading of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Canadell, J.G.; Monteiro, P.M.S.; Costa, M.H.; Cotrim da Cunha, L.; Cox, P.M.; Eliseev, A.V.; Henson, S.; Ishii, M.; Jaccard, S.; Koven, C.; et al. 2021: Global Carbon and other Biogeochemical Cycles and Feedbacks. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2023; pp. 673–816. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, M.U.F. Will Changes in Soil Organic Carbon Act as a Positive or Negative Feedback on Global Warming? Biogeochemistry 2000, 48, 21–51. [Google Scholar] [CrossRef]
- Hashimoto, S. A New Estimation of Global Soil Greenhouse Gas Fluxes Using a Simple Data-Oriented Model. PLoS ONE 2012, 7, e41962. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.; Thomson, A. Temperature-Associated Increases in the Global Soil Respiration Record. Nature 2010, 464, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, H.; Spooner, F.; Roser, M. Forests and Deforestation. Our World Data 2023. Available online: https://ourworldindata.org/forests-and-deforestation (accessed on 28 October 2023).
- Randerson, J.T.; Chapin, F.S.; Harden, J.W.; Neff, J.C.; Harmon, M.E. Net Ecosystem Production: A Comprehensive Measure of Net Carbon Accumulation by Ecosystems. Ecol. Appl. 2002, 12, 937–947. [Google Scholar] [CrossRef]
- Lindauer, M.; Schmid, H.P.; Grote, R.; Mauder, M.; Steinbrecher, R.; Wolpert, B. Net Ecosystem Exchange over a Non-Cleared Wind-Throw-Disturbed Upland Spruce Forest—Measurements and Simulations. Agric. For. Meteorol. 2014, 197, 219–234. [Google Scholar] [CrossRef]
- Yamanoi, K.; Mizoguchi, Y.; Utsugi, H. Effects of a Windthrow Disturbance on the Carbon Balance of a Broadleaf Deciduous Forest in Hokkaido, Japan. Biogeosciences 2015, 12, 6837–6851. [Google Scholar] [CrossRef]
- Hermle, S.; Lavigne, M.B.; Bernier, P.Y.; Bergeron, O.; Pare, D. Component Respiration, Ecosystem Respiration and Net Primary Production of a Mature Black Spruce Forest in Northern Quebec. Tree Physiol. 2010, 30, 527–540. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 3–32. [Google Scholar] [CrossRef]
- Pörtner, H.-O.; Roberts, D.C.; Adams, H.; Adelekan, I.; Adler, C.; Adrian, R.; Aldunce, P.; Ali, E.; Begum, R.A.; Bednar-Friedl, B.; et al. Technical Summary. In Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M.M.B., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., et al., Eds.; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Jakoby, O.; Lischke, H.; Wermelinger, B. Climate Change Alters Elevational Phenology Patterns of the European Spruce Bark Beetle (Ips Typographus). Glob. Chang. Biol. 2019, 25, 4048–4063. [Google Scholar] [CrossRef]
- Romashkin, I.; Neuvonen, S.; Tikkanen, O. Northward Shift in Temperature Sum Isoclines May Favour Ips typographus Outbreaks in European Russia. Agric. For. Entomol. 2020, 22, 238–249. [Google Scholar] [CrossRef]
- Marini, L.; Ayres, M.P.; Battisti, A.; Faccoli, M. Climate Affects Severity and Altitudinal Distribution of Outbreaks in an Eruptive Bark Beetle. Clim. Chang. 2012, 115, 327–341. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.-J.; Rammer, W.; Verkerk, P.J. Increasing Forest Disturbances in Europe and Their Impact on Carbon Storage. Nat. Clim. Chang. 2014, 4, 806–810. [Google Scholar] [CrossRef]
- Hlásny, T.; König, L.; Krokene, P.; Lindner, M.; Montagné-Huck, C.; Müller, J.; Qin, H.; Raffa, K.F.; Schelhaas, M.-J.; Svoboda, M.; et al. Bark Beetle Outbreaks in Europe: State of Knowledge and Ways Forward for Management. Curr. For. Rep. 2021, 7, 138–165. [Google Scholar] [CrossRef]
- Trubin, A.; Mezei, P.; Zabihi, K.; Surový, P.; Jakuš, R. Northernmost European Spruce Bark Beetle Ips typographus Outbreak: Modelling Tree Mortality Using Remote Sensing and Climate Data. For. Ecol. Manag. 2022, 505, 119829. [Google Scholar] [CrossRef]
- Bright, B.C.; Hicke, J.A.; Meddens, A.J.H. Effects of Bark Beetle-caused Tree Mortality on Biogeochemical and Biogeophysical MODIS Products. J. Geophys. Res. Biogeosciences 2013, 118, 974–982. [Google Scholar] [CrossRef]
- Edburg, S.L.; Hicke, J.A.; Brooks, P.D.; Pendall, E.G.; Ewers, B.E.; Norton, U.; Gochis, D.; Gutmann, E.D.; Meddens, A.J. Cascading Impacts of Bark Beetle-caused Tree Mortality on Coupled Biogeophysical and Biogeochemical Processes. Front. Ecol. Environ. 2012, 10, 416–424. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain Pine Beetle and Forest Carbon Feedback to Climate Change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef]
- Moore, D.J.P.; Trahan, N.A.; Wilkes, P.; Quaife, T.; Stephens, B.B.; Elder, K.; Desai, A.R.; Negron, J.; Monson, R.K. Persistent Reduced Ecosystem Respiration after Insect Disturbance in High Elevation Forests. Ecol. Lett. 2013, 16, 731–737. [Google Scholar] [CrossRef]
- Xiong, Y.; D’Atri, J.J.; Fu, S.; Xia, H.; Seastedt, T.R. Rapid Soil Organic Matter Loss from Forest Dieback in a Subalpine Coniferous Ecosystem. Soil Biol. Biochem. 2011, 43, 2450–2456. [Google Scholar] [CrossRef]
- Hu, Y.-H.; Zhang, X.-Y.; Zhang, K.; Song, M.-H.; Gao, J.-Q.; Dorodnikov, M.; Soromotin, A.; Kuzyakov, Y. Tussock Microhabitats Increase Nitrogen Uptake by Plants in an Alpine Wetland. Plant Soil 2021, 466, 569–580. [Google Scholar] [CrossRef]
- Powlson, D.S.; Addiscott, T.M. Nitrogen in Soils|Nitrates. In Encyclopedia of Soils in the Environment; Elsevier: Amsterdam, The Netherlands, 2005; pp. 21–31. [Google Scholar] [CrossRef]
- Cigan, P.W.; Karst, J.; Cahill, J.F.; Sywenky, A.N.; Pec, G.J.; Erbilgin, N. Influence of Bark Beetle Outbreaks on Nutrient Cycling in Native Pine Stands in Western Canada. Plant Soil 2015, 390, 29–47. [Google Scholar] [CrossRef]
- Högberg, M.N.; Högberg, P. Extramatrical Ectomycorrhizal Mycelium Contributes One-third of Microbial Biomass and Produces, Together with Associated Roots, Half the Dissolved Organic Carbon in a Forest Soil. New Phytol. 2002, 154, 791–795. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, X. Soil Respiration and the Environment; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Kobler, J.; Jandl, R.; Dirnböck, T.; Mirtl, M.; Schindlbacher, A. Effects of Stand Patchiness Due to Windthrow and Bark Beetle Abatement Measures on Soil CO2 Efflux and Net Ecosystem Productivity of a Managed Temperate Mountain Forest. Eur. J. For. Res. 2015, 134, 683–692. [Google Scholar] [CrossRef]
- Trahan, N.A.; Dynes, E.L.; Pugh, E.; Moore, D.J.P.; Monson, R.K. Changes in Soil Biogeochemistry Following Disturbance by Girdling and Mountain Pine Beetles in Subalpine Forests. Oecologia 2015, 177, 981–995. [Google Scholar] [CrossRef]
- Jaakkola, E.; Gärtner, A.; Jönsson, A.M.; Ljung, K.; Olsson, P.-O.; Holst, T. Spruce Bark Beetle (Ips typographus) Infestation Cause up to 700 Times Higher Bark BVOC Emission Rates from Norway Spruce (Picea Abies); preprint; Biodiversity and Ecosystem Function. Terrestrial 2022, 7, 1–32. [Google Scholar] [CrossRef]
- Swanson, M.E.; Franklin, J.F.; Beschta, R.L.; Crisafulli, C.M.; DellaSala, D.A.; Hutto, R.L.; Lindenmayer, D.B.; Swanson, F.J. The Forgotten Stage of Forest Succession: Early-successional Ecosystems on Forest Sites. Front. Ecol. Environ. 2011, 9, 117–125. [Google Scholar] [CrossRef]
- Morehouse, K.; Johns, T.; Kaye, J.; Kaye, M. Carbon and Nitrogen Cycling Immediately Following Bark Beetle Outbreaks in Southwestern Ponderosa Pine Forests. For. Ecol. Manag. 2008, 255, 2698–2708. [Google Scholar] [CrossRef]
- Reed, D.E.; Ewers, B.E.; Pendall, E. Impact of Mountain Pine Beetle Induced Mortality on Forest Carbon and Water Fluxes. Environ. Res. Lett. 2014, 9, 105004. [Google Scholar] [CrossRef]
- Mayer, M.; Matthews, B.; Rosinger, C.; Sandén, H.; Godbold, D.L.; Katzensteiner, K. Tree Regeneration Retards Decomposition in a Temperate Mountain Soil after Forest Gap Disturbance. Soil Biol. Biochem. 2017, 115, 490–498. [Google Scholar] [CrossRef]
- Hursh, A.; Ballantyne, A.; Cooper, L.; Maneta, M.; Kimball, J.; Watts, J. The Sensitivity of Soil Respiration to Soil Temperature, Moisture, and Carbon Supply at the Global Scale. Glob. Chang. Biol. 2017, 23, 2090–2103. [Google Scholar] [CrossRef]
- Chen, S.; Huang, Y.; Zou, J.; Shen, Q.; Hu, Z.; Qin, Y.; Chen, H.; Pan, G. Modeling Interannual Variability of Global Soil Respiration from Climate and Soil Properties. Agric. For. Meteorol. 2010, 150, 590–605. [Google Scholar] [CrossRef]
- Tang, J.; Bradford, M.A.; Carey, J.; Crowther, T.W.; Machmuller, M.B.; Mohan, J.E.; Todd-Brown, K. Temperature Sensitivity of Soil Carbon. In Ecosystem Consequences of Soil Warming; Elsevier: Amsterdam, The Netherlands, 2019; pp. 175–208. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, M.; Kang, M.; Xing, K.; Wang, Y.; Xue, F.; Chen, C. Diurnal and Seasonal Variation of the Elevation Gradient of Air Temperature in the Northern Flank of the Western Qinling Mountain Range, China. J. Mt. Sci. 2017, 14, 94–105. [Google Scholar] [CrossRef]
- Chen, B.-X.; Sun, Y.-F.; Zhang, H.-B.; Han, Z.-H.; Wang, J.-S.; Li, Y.-K.; Yang, X.-L. Temperature Change along Elevation and Its Effect on the Alpine Timberline Tree Growth in the Southeast of the Tibetan Plateau. Adv. Clim. Change Res. 2018, 9, 185–191. [Google Scholar] [CrossRef]
- Makita, N.; Kosugi, Y.; Sakabe, A.; Kanazawa, A.; Ohkubo, S.; Tani, M. Seasonal and Diurnal Patterns of Soil Respiration in an Evergreen Coniferous Forest: Evidence from Six Years of Observation with Automatic Chambers. PLoS ONE 2018, 13, e0192622. [Google Scholar] [CrossRef] [PubMed]
- Speckman, H.N.; Frank, J.M.; Bradford, J.B.; Miles, B.L.; Massman, W.J.; Parton, W.J.; Ryan, M.G. Forest Ecosystem Respiration Estimated from Eddy Covariance and Chamber Measurements under High Turbulence and Substantial Tree Mortality from Bark Beetles. Glob. Chang. Biol. 2015, 21, 708–721. [Google Scholar] [CrossRef]
- Badraghi, A.; Ventura, M.; Polo, A.; Borruso, L.; Giammarchi, F.; Montagnani, L. Soil Respiration Variation along an Altitudinal Gradient in the Italian Alps: Disentangling Forest Structure and Temperature Effects. PLoS ONE 2021, 16, e0247893. [Google Scholar] [CrossRef]
- DeForest, J.L.; Noormets, A.; McNulty, S.G.; Sun, G.; Tenney, G.; Chen, J. Phenophases Alter the Soil Respiration–Temperature Relationship in an Oak-Dominated Forest. Int. J. Biometeorol. 2006, 51, 135–144. [Google Scholar] [CrossRef]
- Mathes, K.; Schriefer, T. Soil Respiration during Secondary Succession: Influence of Temperature and Moisture. Soil Biol. Biochem. 1985, 17, 205–211. [Google Scholar] [CrossRef]
- Davidson, E.A.; Holbrook, N.M. Is Temporal Variation of Soil Respiration Linked to the Phenology of Photosynthesis? In Phenology of Ecosystem Processes; Noormets, A., Ed.; Springer New York: New York, NY, USA, 2009; pp. 187–199. [Google Scholar] [CrossRef]
- Prach, K.; Pyšek, P. How Do Species Dominating in Succession Differ from Others? J. Veg. Sci. 1999, 10, 383–392. [Google Scholar] [CrossRef]
- Sedláková, I.; Fiala, K. Ecological Problems of Degradation of Alluvial Meadows Due to Expanding Calamagrostis Epigejos. Ekol. Bratisl. 2001, 20, 226–233. [Google Scholar]
- Mayer, M.; Matthews, B.; Schindlbacher, A.; Katzensteiner, K. Soil CO2 Efflux from Mountainous Windthrow Areas: Dynamics over 12 Years Post-Disturbance. Biogeosciences 2014, 11, 6081–6093. [Google Scholar] [CrossRef]
- Budzáková, M.; Galvánek, D.; Littera, P.; Šibík, J. The Wind and Fire Disturbance in Central European Mountain Spruce Forests: The Regeneration after Four Years. Acta Soc. Bot. Pol. 2013, 81, 13–24. [Google Scholar] [CrossRef]
- Pyšek, P. What Do We Know about Calamagrostis Villosa? A Review of the Species Behaviour in Secondary Habitats. Preslia 1993, 65, 1–20. [Google Scholar]
- Chang, S.X.; Shi, Z.; Thomas, B.R. Soil Respiration and Net Ecosystem Productivity in a Chronosequence of Hybrid Poplar Plantations1. Can. J. Soil Sci. 2020, 100, 488–502. [Google Scholar] [CrossRef]
- Macdonald, C.A.; Anderson, I.C.; Khachane, A.; Singh, B.P.; Barton, C.V.M.; Duursma, R.A.; Ellsworth, D.S.; Singh, B.K. Plant Productivity Is a Key Driver of Soil Respiration Response to Climate Change in a Nutrient-Limited Soil. Basic Appl. Ecol. 2021, 50, 155–168. [Google Scholar] [CrossRef]
- Caprez, R.; Niklaus, P.A.; Körner, C. Forest Soil Respiration Reflects Plant Productivity across a Temperature Gradient in the Alps. Oecologia 2012, 170, 1143–1154. [Google Scholar] [CrossRef]
- Rodeghiero, M.; Cescatti, A. Main Determinants of Forest Soil Respiration along an Elevation/Temperature Gradient in the Italian Alps. Glob. Chang. Biol. 2005, 11, 1024–1041. [Google Scholar] [CrossRef]
- Zheng, X.; Zhao, C.; Peng, S.; Jian, S.; Liang, B.; Wang, X.; Yang, S.; Wang, C.; Peng, H.; Wang, Y. Soil CO2 Efflux along an Elevation Gradient in Qinghai Spruce Forests in the Upper Reaches of the Heihe River, Northwest China. Environ. Earth Sci. 2014, 71, 2065–2076. [Google Scholar] [CrossRef]
- Borkhuu, B.; Peckham, S.D.; Ewers, B.E.; Norton, U.; Pendall, E. Does Soil Respiration Decline Following Bark Beetle Induced Forest Mortality? Evidence from a Lodgepole Pine Forest. Agric. For. Meteorol. 2015, 214, 201–207. [Google Scholar] [CrossRef]
- Jamnická, G.; Konôpková, A.; Fleischer, P.; Kurjak, D.; Petrík, P.; Petek, A.; Húdoková, H.; Fleischer, P.; Homolová, Z.; Ježík, M.; et al. Physiological Vitality of Norway Spruce (Picea Abies L.) Stands along an Altitudinal Gradient in Tatra National Park. Cent. Eur. For. J. 2020, 66, 227–242. [Google Scholar] [CrossRef]
- Fleischer, P.; Homolová, Z. Tatry Ako Objekt Dlhodobého Ekologického Výskumu Prírodných Disturbancií. Životné Prostr. 2016, 50, 1–4. [Google Scholar]
- Fleischer, P.; Homolová, Z. Long-Term Research on Ecological Condition in the Larch-Spruce Forest in High Tatras after Natural Disturbances. Lesn. Čas. For. J. 2011, 57, 237–250. [Google Scholar]
- Elkin, L.A.; Kay, M.; Higgins, J.J.; Wobbrock, J.O. An Aligned Rank Transform Procedure for Multifactor Contrast Tests. In Proceedings of the 34th Annual ACM Symposium on User Interface Software and Technology, Virtual, 10–14 October 2021; pp. 754–768. [Google Scholar] [CrossRef]
- Lei, N.; Wang, H.; Zhang, Y.; Chen, T. Components of Respiration and Their Temperature Sensitivity in Four Reconstructed Soils. Sci. Rep. 2022, 12, 6107. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Sha, L.; Wu, C.; Tan, Z.; Song, Q.; Liu, Y.; Dong, L. Effects of Continuous Drought Stress on Soil Respiration in a Tropical Rainforest in Southwest China. Plant Soil 2015, 394, 343–353. [Google Scholar] [CrossRef]
- Tang, X.; Zhou, G.; Liu, S.; Zhang, D.; Liu, S.; Li, J.; Zhou, C. Dependence of Soil Respiration on Soil Temperature and Soil Moisture in Successional Forests in Southern China. J. Integr. Plant Biol. 2006, 48, 654–663. [Google Scholar] [CrossRef]
- Högberg, P.; Nordgren, A.; Buchmann, N.; Taylor, A.F.; Ekblad, A.; Högberg, M.N.; Nyberg, G.; Ottosson-Löfvenius, M.; Read, D.J. Large-Scale Forest Girdling Shows That Current Photosynthesis Drives Soil Respiration. Nature 2001, 411, 789–792. [Google Scholar] [CrossRef]
- Levy-Varon, J.H.; Schuster, W.S.F.; Griffin, K.L. Rapid Rebound of Soil Respiration Following Partial Stand Disturbance by Tree Girdling in a Temperate Deciduous Forest. Oecologia 2014, 174, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Brouillard, B.M.; Mikkelson, K.M.; Bokman, C.M.; Berryman, E.M.; Sharp, J.O. Extent of Localized Tree Mortality Influences Soil Biogeochemical Response in a Beetle-Infested Coniferous Forest. Soil Biol. Biochem. 2017, 114, 309–318. [Google Scholar] [CrossRef]
- Han, M.; Feng, J.; Chen, Y.; Sun, L.; Fu, L.; Zhu, B. Mycorrhizal Mycelial Respiration: A Substantial Component of Soil Respired CO2. Soil Biol. Biochem. 2021, 163, 108454. [Google Scholar] [CrossRef]
- Klutsch, J.G.; Negrón, J.F.; Costello, S.L.; Rhoades, C.C.; West, D.R.; Popp, J.; Caissie, R. Stand Characteristics and Downed Woody Debris Accumulations Associated with a Mountain Pine Beetle (Dendroctonus Ponderosae Hopkins) Outbreak in Colorado. For. Ecol. Manag. 2009, 258, 641–649. [Google Scholar] [CrossRef]
- Whitney, G.G. The Productivity and Carbohydrate Economy of a Developing Stand of Rubus Idaeus. Can. J. Bot. 1982, 60, 2697–2703. [Google Scholar] [CrossRef]
- Tang, X.; Pei, X.; Lei, N.; Luo, X.; Liu, L.; Shi, L.; Chen, G.; Liang, J. Global Patterns of Soil Autotrophic Respiration and Its Relation to Climate, Soil and Vegetation Characteristics. Geoderma 2020, 369, 114339. [Google Scholar] [CrossRef]
- Zeeshan, M.; Wenjun, Z.; Chuansheng, W.; Yan, L.; Azeez, P.A.; Qinghai, S.; Yuntong, L.; Yiping, Z.; Zhiyun, L.; Liqing, S. Soil Heterotrophic Respiration in Response to Rising Temperature and Moisture along an Altitudinal Gradient in a Subtropical Forest Ecosystem, Southwest China. Sci. Total Environ. 2022, 816, 151643. [Google Scholar] [CrossRef] [PubMed]
- Yazaki, T.; Hirano, T.; Sano, T. Biomass Accumulation and Net Primary Production during the Early Stage of Secondary Succession after a Severe Forest Disturbance in Northern Japan. Forests 2016, 7, 287. [Google Scholar] [CrossRef]
- Norton, U.; Ewers, B.E.; Borkhuu, B.; Brown, N.R.; Pendall, E. Soil Nitrogen Five Years after Bark Beetle Infestation in Lodgepole Pine Forests. Soil Sci. Soc. Am. J. 2015, 79, 282–293. [Google Scholar] [CrossRef]
- Kubov, M.; Janík, R.; Tomes, J.; Schieber, B. Nutrient Content in Biomass of Bilberry (Vaccinium Myrtillus L.) in Different Habitats of Protected Areas of Inner Western Carpathians. J. For. Sci. 2024; 70, in press. [Google Scholar]
- Mayer, M.; Sandén, H.; Rewald, B.; Godbold, D.; Katzensteiner, K. Increase in Heterotrophic Soil Respiration by Temperature Drives Decline in Soil Organic Carbon Stocks after Forest Windthrow in a Mountainous Ecosystem. Funct. Ecol. 2017, 31, 1163–1172. [Google Scholar] [CrossRef]
- Maurer, G.E.; Chan, A.M.; Trahan, N.A.; Moore, D.J.P.; Bowling, D.R. Carbon Isotopic Composition of Forest Soil Respiration in the Decade Following Bark Beetle and Stem Girdling Disturbances in the Rocky Mountains. Plant Cell Environ. 2016, 39, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.D.; Natali, S.M.; Minions, C.; Risk, D.; Arndt, K.; Zona, D.; Euskirchen, E.S.; Rocha, A.V.; Sonnentag, O.; Helbig, M.; et al. Soil Respiration Strongly Offsets Carbon Uptake in Alaska and Northwest Canada. Environ. Res. Lett. 2021, 16, 084051. [Google Scholar] [CrossRef]
- Ananyeva, N.D.; Sushko, S.V.; Ivashchenko, K.V.; Vasenev, V.I. Soil Microbial Respiration in Subtaiga and Forest-Steppe Ecosystems of European Russia: Field and Laboratory Approaches. Eurasian Soil Sci. 2020, 53, 1492–1501. [Google Scholar] [CrossRef]
- Luo, S.; Liu, G.; Li, Z.; Hu, C.; Gong, L.; Wang, M.; Hu, H. Soil Respiration along an Altitudinal Gradient in a Subalpine Secondary Forest in China. IForest Biogeosci. For. 2015, 8, 526–532. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, Y.; Zhang, Y.; Cui, Y.; Wu, J.; Zhang, W.; Wang, D.; Zou, J. Interactions between Nitrogen and Phosphorus in Modulating Soil Respiration: A Meta-Analysis. Sci. Total Environ. 2023, 905, 167346. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Shang, H. Contribution of Soil Respiration to the Global Carbon Equation. J. Plant Physiol. 2016, 203, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Kuzyakov, Y. Sources of CO2 Efflux from Soil and Review of Partitioning Methods. Soil Biol. Biochem. 2006, 38, 425–448. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Wang, J.; Chen, W.; Tian, D.; Niu, S. Different Responses of Soil Respiration and Its Components to Nitrogen and Phosphorus Addition in a Subtropical Secondary Forest. For. Ecosyst. 2021, 8, 37. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).