Elevational Gradients of Soil Nematode Communities in Subtropical Forest Ecosystems


 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Soil Sampling and Measurements
2.3. Measurements of Soil Properties
2.4. Statistical Analysis
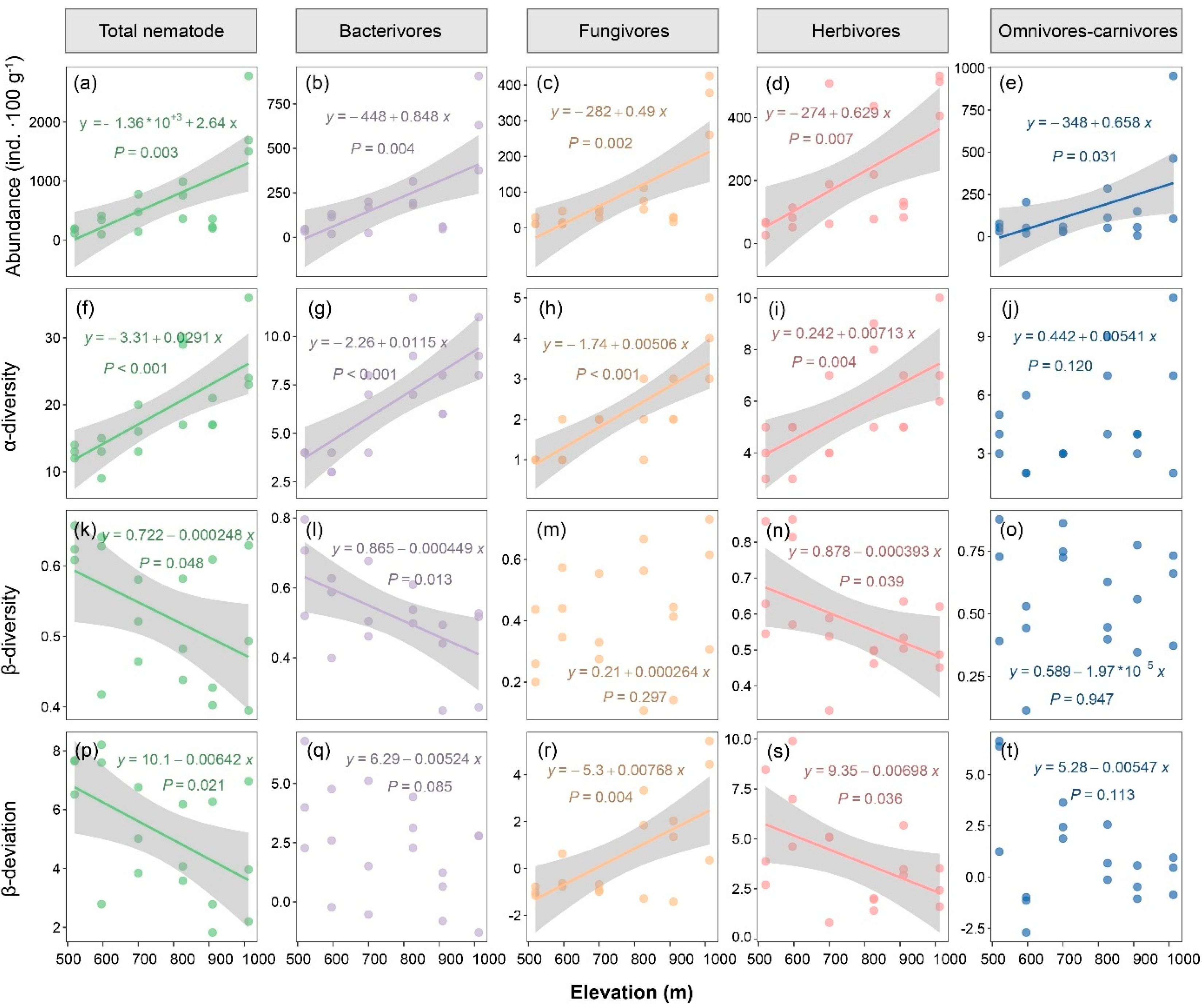
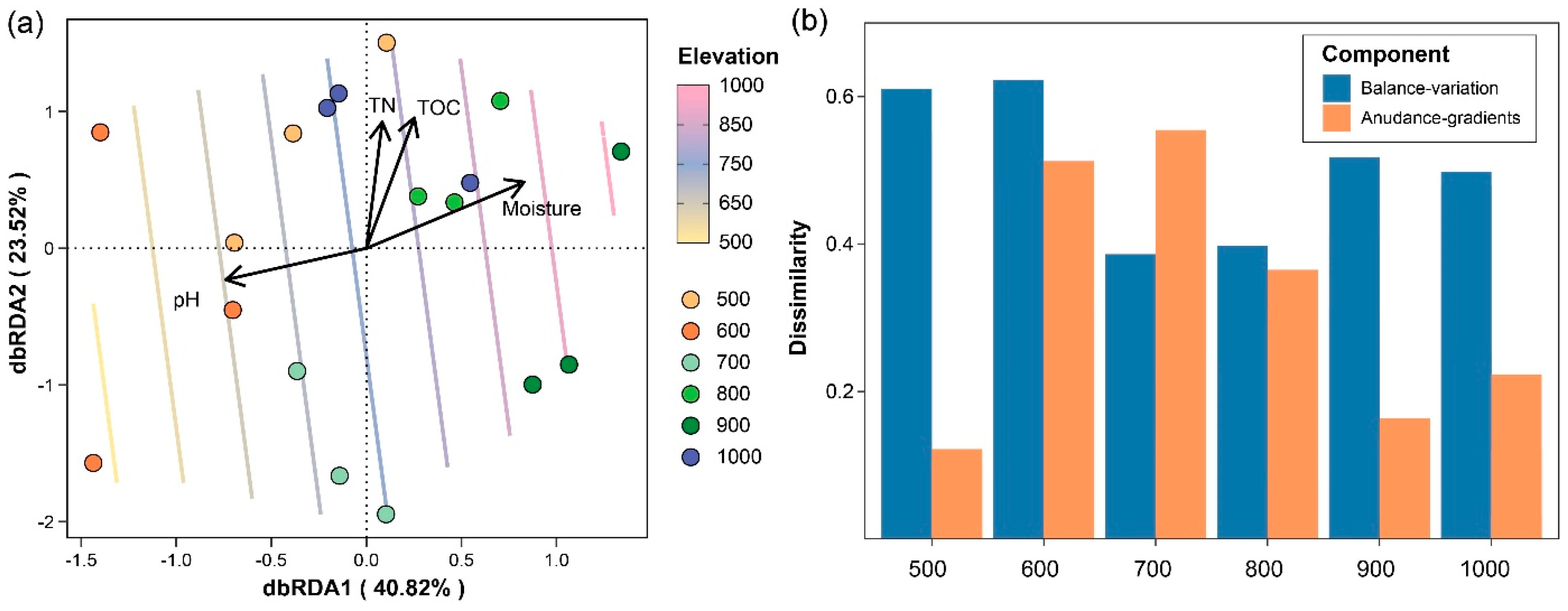
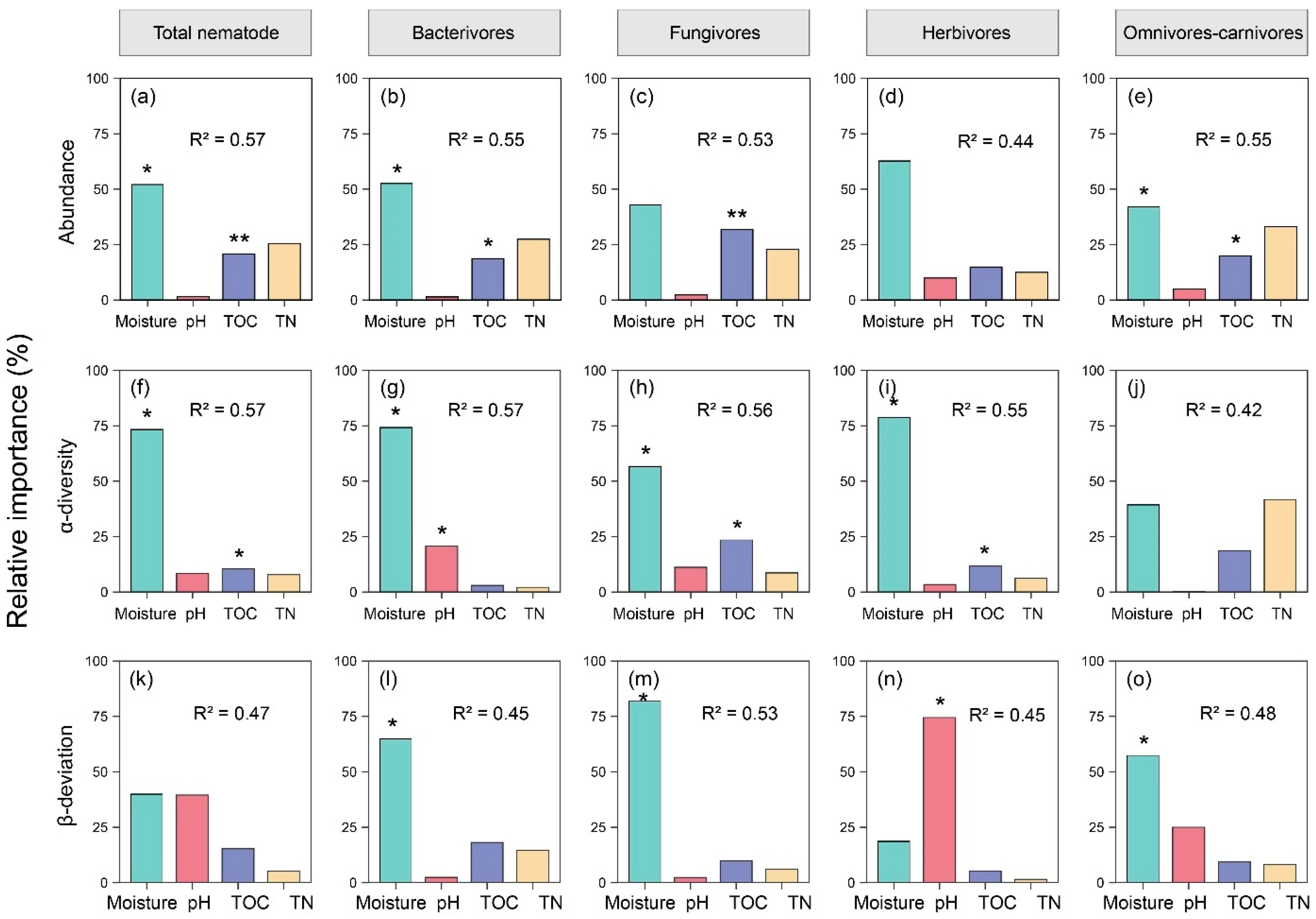
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bardgett, R.D.; Van Der Putten, W.H. Belowground Biodiversity and Ecosystem Functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Wall, D.H.; Bardgett, R.D.; Kelly, E. Biodiversity in the Dark. Nat. Geosci. 2010, 3, 297–298. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Reich, P.B.; Trivedi, C.; Eldridge, D.J.; Abades, S.; Alfaro, F.D.; Bastida, F.; Berhe, A.A.; Cutler, N.A.; Gallardo, A.; et al. Multiple Elements of Soil Biodiversity Drive Ecosystem Functions across Biomes. Nat. Ecol. Evol. 2020, 4, 210–220. [Google Scholar] [CrossRef]
- Wagg, C.; Bender, S.F.; Widmer, F.; Van Der Heijden, M.G.A. Soil Biodiversity and Soil Community Composition Determine Ecosystem Multifunctionality. Proc. Natl. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef]
- Wu, L.; Chen, H.; Chen, D.; Wang, S.; Wu, Y.; Wang, B.; Liu, S.; Yue, L.; Yu, J.; Bai, Y. Soil Biota Diversity and Plant Diversity Both Contributed to Ecosystem Stability in Grasslands. Ecol. Lett. 2023, 26, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Jackson, R.B. The Diversity and Biogeography of Soil Bacterial Communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef]
- Hanson, C.A.; Fuhrman, J.A.; Horner-Devine, M.C.; Martiny, J.B.H. Beyond Biogeographic Patterns: Processes Shaping the Microbial Landscape. Nat. Rev. Microbiol. 2012, 10, 497–506. [Google Scholar] [CrossRef]
- Shen, C.; Liang, W.; Shi, Y.; Lin, X.; Zhang, H.; Wu, X.; Xie, G.; Chain, P.; Grogan, P.; Chu, H. Contrasting Elevational Diversity Patterns between Eukaryotic Soil Microbes and Plants. Ecology 2014, 95, 3190–3202. [Google Scholar] [CrossRef]
- Li, X.; Liu, Z.; Zhang, C.; Zheng, L.; Li, H. Altitudinal Variation in Soil Nematode Communities in an Alpine Mountain Region of the Eastern Tibetan Plateau. Eur. J. Soil Biol. 2024, 121, 103617. [Google Scholar] [CrossRef]
- Bokhorst, S.; (Ciska) Veen, G.F.; Sundqvist, M.; De Long, J.R.; Kardol, P.; Wardle, D.A. Contrasting Responses of Springtails and Mites to Elevation and Vegetation Type in the Sub-Arctic. Pedobiologia 2018, 67, 57–64. [Google Scholar] [CrossRef]
- Bryant, J.A.; Lamanna, C.; Morlon, H.; Kerkhoff, A.J.; Enquist, B.J.; Green, J.L. Microbes on Mountainsides: Contrasting Elevational Patterns of Bacterial and Plant Diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 11505–11511. [Google Scholar] [CrossRef]
- Ma, L.; Liu, L.; Lu, Y.; Chen, L.; Zhang, Z.; Zhang, H.; Wang, X.; Shu, L.; Yang, Q.; Song, Q. When Microclimates Meet Soil Microbes: Temperature Controls Soil Microbial Diversity along an Elevational Gradient in Subtropical Forests. Soil Biol. Biochem. 2022, 166, 108566. [Google Scholar] [CrossRef]
- Song, L.; Yang, T.; Xia, S.; Yin, Z.; Liu, X.; Li, S.; Sun, R.; Gao, H.; Chu, H.; Ma, C. Soil Depth Exerts Stronger Impact on Bacterial Community than Elevation in Subtropical Forests of Huangshan Mountain. Sci. Total Environ. 2022, 852, 158438. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Si, G.; Wang, J.; Luo, T.; Zhang, G. Bacterial Community in Alpine Grasslands along an Altitudinal Gradient on the Tibetan Plateau. FEMS Microbiol. Ecol. 2014, 87, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Mori, A.S.; Isbell, F.; Seidl, R. β-Diversity, Community Assembly, and Ecosystem Functioning. Trends Ecol. Evol. 2018, 33, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Omidipour, R.; Tahmasebi, P.; Faal Faizabadi, M.; Faramarzi, M.; Ebrahimi, A. Does β Diversity Predict Ecosystem Productivity Better than Species Diversity? Ecol. Indic. 2021, 122, 107212. [Google Scholar] [CrossRef]
- Wang, S.; Loreau, M. Biodiversity and Ecosystem Stability across Scales in Metacommunities. Ecol. Lett. 2016, 19, 510–518. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning Abundance-based Multiple-site Dissimilarity into Components: Balanced Variation in Abundance and Abundance Gradients. Methods Ecol. Evol. 2017, 8, 799–808. [Google Scholar] [CrossRef]
- Li, S.; Wang, P.; Chen, Y.; Wilson, M.C.; Yang, X.; Ma, C.; Lu, J.; Chen, X.; Wu, J.; Shu, W.; et al. Island Biogeography of Soil Bacteria and Fungi: Similar Patterns, but Different Mechanisms. ISME J. 2020, 14, 1886–1896. [Google Scholar] [CrossRef]
- van den Hoogen, J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.A.; de Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.S.; et al. Soil Nematode Abundance and Functional Group Composition at a Global Scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef]
- Yeates, G.W.; Bongers, T.; De Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding Habits in Soil Nematode Families and Genera-An Outline for Soil Ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- de Vries, F.T.; Thébault, E.; Liiri, M.; Birkhofer, K.; Tsiafouli, M.A.; Bjørnlund, L.; Jørgensen, H.B.; Brady, M.V.; Christensen, S.; De Ruiter, P.C.; et al. Soil Food Web Properties Explain Ecosystem Services across European Land Use Systems. Proc. Natl. Acad. Sci. USA 2013, 110, 14296–14301. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Wang, Z.; Liu, T.; Kardol, P.; Ma, C.; Hu, Y.; Cui, Y.; Zhao, C.; Zhang, W.; Guo, D. Drivers of Nematode Diversity in Forest Soils across Climatic Zones. Proc. R. Soc. B 2023, 290, 20230107. [Google Scholar] [CrossRef]
- Li, X.; Liu, T.; Li, H.; Geisen, S.; Hu, F.; Liu, M. Management Effects on Soil Nematode Abundance Differ among Functional Groups and Land-Use Types at a Global Scale. J. Anim. Ecol. 2022, 91, 1770–1780. [Google Scholar] [CrossRef]
- Griffiths, B.S. Microbial-Feeding Nematodes and Protozoa in Soil: Their Effectson Microbial Activity and Nitrogen Mineralization in Decomposition Hotspots and the Rhizosphere. Plant Soil 1994, 164, 25–33. [Google Scholar] [CrossRef]
- Bonkowski, M.; Villenave, C.; Griffiths, B. Rhizosphere Fauna: The Functional and Structural Diversity of Intimate Interactions of Soil Fauna with Plant Roots. Plant Soil 2009, 321, 213–233. [Google Scholar] [CrossRef]
- Topalović, O.; Geisen, S. Nematodes as Suppressors and Facilitators of Plant Performance. New Phytol. 2023, 238, 2305–2312. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Cook, R.; Yeates, G.W.; Denton, C.S. The Influence of Nematodes on Below-Ground Processes in Grassland Ecosystems. Plant Soil 1999, 212, 23–33. [Google Scholar] [CrossRef]
- Chen, D.; Lan, Z.; Hu, S.; Bai, Y. Effects of Nitrogen Enrichment on Belowground Communities in Grassland: Relative Role of Soil Nitrogen Availability vs. Soil Acidification. Soil Biol. Biochem. 2015, 89, 99–108. [Google Scholar] [CrossRef]
- Vandegehuchte, M.L.; Sylvain, Z.A.; Reichmann, L.G.; De Tomasel, C.M.; Nielsen, U.N.; Wall, D.H.; Sala, O.E. Responses of a Desert Nematode Community to Changes in Water Availability. Ecosphere 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Xiao, H.; Wang, B.; Lu, S.; Chen, D.; Wu, Y.; Zhu, Y.; Hu, S.; Bai, Y. Soil Acidification Reduces the Effects of Short-Term Nutrient Enrichment on Plant and Soil Biota and Their Interactions in Grasslands. Glob. Chang. Biol. 2020, 26, 4626–4637. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Cheng, J.; Chu, P.; Hu, S.; Xie, Y.; Tuvshintogtokh, I.; Bai, Y. Regional-Scale Patterns of Soil Microbes and Nematodes across Grasslands on the Mongolian Plateau: Relationships with Climate, Soil, and Plants. Ecography 2015, 38, 622–631. [Google Scholar] [CrossRef]
- Lu, L.; Li, G.; He, N.; Li, H.; Liu, T.; Li, X.; Whalen, J.K.; Geisen, S.; Liu, M. Drought Shifts Soil Nematodes to Smaller Size across Biological Scales. Soil Biol. Biochem. 2023, 184, 109099. [Google Scholar] [CrossRef]
- Steffan, S.A.; Chikaraishi, Y.; Currie, C.R.; Horn, H.; Gaines-Day, H.R.; Pauli, J.N.; Zalapa, J.E.; Ohkouchi, N. Microbes Are Trophic Analogs of Animals. Proc. Natl. Acad. Sci. USA 2015, 112, 15119–15124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liang, W.-J.J.; Zhang, X.-K.K.; Liang, W.-J.J. Soil Nematode Abundance and Diversity in Different Forest Types at Changbai Mountain, China. Zool. Stud. 2012, 51, 619–626. [Google Scholar]
- Kergunteuil, A.; Campos-Herrera, R.; Sánchez-Moreno, S.; Vittoz, P.; Rasmann, S. The Abundance, Diversity, and Metabolic Footprint of Soil Nematodes Is Highest in High Elevation Alpine Grasslands. Front. Ecol. Evol. 2016, 4, 84. [Google Scholar] [CrossRef]
- Kouser, Y.; Shah, A.A.; Rasmann, S. The Functional Role and Diversity of Soil Nematodes Are Stronger at High Elevation in the Lesser Himalayan Mountain Ranges. Ecol. Evol. 2021, 11, 13793–13804. [Google Scholar] [CrossRef]
- Devetter, M.; Háněl, L.; Řeháková, K.; Doležal, J. Diversity and Feeding Strategies of Soil Microfauna along Elevation Gradients in Himalayan Cold Deserts. PLoS ONE 2017, 12, e0187646. [Google Scholar] [CrossRef]
- Pitteloud, C.; Descombes, P.; Sànchez-Moreno, S.; Kergunteuil, A.; Ibanez, S.; Rasmann, S.; Pellissier, L. Contrasting Responses of Above- and below-Ground Herbivore Communities along Elevation. Oecologia 2020, 194, 515–528. [Google Scholar] [CrossRef]
- Liu, W.; Liu, L.; Yang, X.; Deng, M.; Wang, Z.; Wang, P.; Yang, S.; Li, P.; Peng, Z.; Yang, L.; et al. Long-Term Nitrogen Input Alters Plant and Soil Bacterial, but Not Fungal Beta Diversity in a Semiarid Grassland. Glob. Chang. Biol. 2021, 27, 3939–3950. [Google Scholar] [CrossRef]
- González, G.; García, E.; Cruz, V.; Borges, S.; Zalamea, M.; Rivera, M.M. Earthworm Communities along an Elevation Gradient in Northeastern Puerto Rico. Eur. J. Soil Biol. 2007, 43, S24–S32. [Google Scholar] [CrossRef]
- Decaëns, T. Macroecological Patterns in Soil Communities. Glob. Ecol. Biogeogr. 2010, 19, 287–302. [Google Scholar] [CrossRef]
- Liu, D.; Liu, D.; Yu, H.; Wu, H. Strong Variations and Shifting Mechanisms of Altitudal Diversity and Abundance Patterns in Soil Oribatid Mites (Acari: Oribatida) on the Changbai Mountain, China. Appl. Soil Ecol. 2023, 186, 104808. [Google Scholar] [CrossRef]
- Xie, Z.; Sun, X.; Lux, J.; Chen, T.; Potapov, M.; Wu, D.; Scheu, S. Drivers of Collembola Assemblages along an Altitudinal Gradient in Northeast China. Ecol. Evol. 2022, 12, e8559. [Google Scholar] [CrossRef] [PubMed]
- Dong, K.; Moroenyane, I.; Tripathi, B.; Kerfahi, D.; Takahashi, K.; Yamamoto, N.; An, C.; Cho, H.; Adams, J. Soil Nematodes Show a Mid-Elevation Diversity Maximum and Elevational Zonation on Mt. Norikura, Japan. Sci. Rep. 2017, 7, 3028. [Google Scholar] [CrossRef]
- Xiong, D.; Wei, C.Z.; Wubs, E.R.J.; Veen, G.F.; Liang, W.; Wang, X.; Li, Q.; Van der Putten, W.H.; Han, X. Nonlinear Responses of Soil Nematode Community Composition to Increasing Aridity. Glob. Ecol. Biogeogr. 2020, 29, 117–126. [Google Scholar] [CrossRef]
- Soininen, J.; Heino, J.; Wang, J. A Meta-analysis of Nestedness and Turnover Components of Beta Diversity across Organisms and Ecosystems. Glob. Ecol. Biogeogr. 2018, 27, 96–109. [Google Scholar] [CrossRef]
- Adams, R.I.; Miletto, M.; Taylor, J.W.; Bruns, T.D. Dispersal in Microbes: Fungi in Indoor Air Are Dominated by Outdoor Air and Show Dispersal Limitation at Short Distances. ISME J. 2013, 7, 1262–1273. [Google Scholar] [CrossRef]
- Peay, K.G.; Garbelotto, M.; Bruns, T.D. Evidence of Dispersal Limitation in Soil Microorganisms: Isolation Reduces Species Richness on Mycorrhizal Tree Islands. Ecology 2010, 91, 3631–3640. [Google Scholar] [CrossRef]
- de Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S.; Hallin, S.; Kaisermann, A.; Keith, A.M.; Kretzschmar, M.; et al. Soil Bacterial Networks Are Less Stable under Drought than Fungal Networks. Nat. Commun. 2018, 9, 3033. [Google Scholar] [CrossRef]
- Yang, T.; Tedersoo, L.; Liu, X.; Gao, G.; Dong, K.; Adams, J.M.; Chu, H. Fungi Stabilize Multi-kingdom Community in a High Elevation Timberline Ecosystem. iMeta 2022, 1, e49. [Google Scholar] [CrossRef] [PubMed]
- Xiong, D.; Wei, C.; Wang, X.; Xiaotao, L.; Fang, S.; Li, Y.; Wang, X.; Liang, W.; Han, X.; Bezemer, T.M.; et al. Spatial Patterns and Ecological Drivers of Soil Nematode β-Diversity in Natural Grasslands Vary among Vegetation Types and Trophic Position. J. Anim. Ecol. 2021, 90, 1367–1378. [Google Scholar] [CrossRef]
- de Vries, F.T.; Liiri, M.E.; Bjørnlund, L.; Bowker, M.A.; Christensen, S.; Setälä, H.M.; Bardgett, R.D. Land Use Alters the Resistance and Resilience of Soil Food Webs to Drought. Nat. Clim. Chang. 2012, 2, 276–280. [Google Scholar] [CrossRef]
- Hu, Z.; Chen, C.; Chen, X.; Yao, J.; Jiang, L.; Liu, M. Home-Field Advantage in Soil Respiration and Its Resilience to Drying and Rewetting Cycles. Sci. Total Environ. 2021, 750, 141736. [Google Scholar] [CrossRef] [PubMed]
- Rahbek, C. The Elevational Gradient of Species Richness: A Uniform Pattern? Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Thakur, M.P.; Geisen, S. Trophic Regulations of the Soil Microbiome. Trends Microbiol. 2019, 27, 771–780. [Google Scholar] [CrossRef]
- Jiang, Y.; Luan, L.; Hu, K.; Liu, M.; Chen, Z.; Geisen, S.; Chen, X.; Li, H.; Xu, Q.; Bonkowski, M.; et al. Trophic Interactions as Determinants of the Arbuscular Mycorrhizal Fungal Community with Cascading Plant-Promoting Consequences. Microbiome 2020, 8, 142. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Elevation (m) | pH | Moisture (%) | TN (g/kg) | TOC (g/kg) |
|---|---|---|---|---|
| 500 | 4.6 ± 0.3 bc | 22.0 ± 10.2 b | 2.4 ± 0.5 ab | 37.4 ± 1.9 ab |
| 600 | 5.0 ± 0.1 a | 32.6 ± 2.7 b | 2.6 ± 1.0 ab | 36.9 ± 13.5 ab |
| 700 | 4.8 ± 0.3 ab | 35.1 ± 5.8 b | 1.5 ± 0.6 b | 26.4 ± 5.3 b |
| 800 | 4.5 ± 0.1 bc | 50.6 ± 2.3 a | 2.5 ± 0.4 ab | 34.4 ± 9.8 b |
| 900 | 4.2 ± 0.2 c | 56.0 ± 9.5 a | 2.4 ± 1.6 ab | 41.5 ± 9.4 ab |
| 1000 | 4.4 ± 0.2 bc | 62.2 ± 11.9 a | 3.7 ± 0.5 a | 52.4 ± 0.3 a |
| Trophic Level | Genus | 500 m | 600 m | 700 m | 800 m | 900 m | 1000 |
|---|---|---|---|---|---|---|---|
| FF | Filenchus | 17.2 ± 10.5 b | 16.9 ± 8.4 b | 28.4 ± 7.2 b | 56.8 ± 33.6 ab | 18.5 ± 11.5 b | 181.8 ± 130.2 a |
| BF | Aphanolaimus | 8.7 ± 7.7 a | 35.7 ± 23.9 a | 24.8 ± 24.6 a | 64.3 ± 10.4 a | 13.1 ± 11.8 a | 126.3 ± 110.6 a |
| HF | Malenchus | 11.0 ± 10.8 a | 18.4 ± 3.7 a | 85.4 ± 114.5 a | 32.9 ± 24.7 a | 15.9 ± 13.0 a | 83.9 ± 62.2 a |
| HF | Tylenchus | 8.1 ± 3.8 b | 10.2 ± 13.8 b | 44.2 ± 43.3 ab | 33.8 ± 41.0 ab | 22.0 ± 6.7 b | 128.4 ± 67.1 a |
| OP | Tripyla | 11.8 ± 9.1 ab | 47.6 ± 52.4 ab | 4.65 ± 8.1 ab | 16.1 ± 27.9 ab | 0 ± 0 b | 158.0 ± 124.9 a |
| BF | Alaimus | 7.4 ± 6.4 b | 13.6 ± 23.6 b | 24.3 ± 29.8 b | 27.7 ± 12.7 ab | 3.6 ± 6.3 b | 106.2 ± 57.7 a |
| OP | Eudorylaimus | 11.8 ± 12.3 a | 0 ± 0 a | 13.0 ± 5.4 a | 25.6 ± 34.3 a | 0 ± 0 a | 118.5 ± 104.3 a |
| BF | Eucephalobus | 8.5 ± 7.6 b | 0 ± 0 b | 5.8 ± 10.1 b | 20.7 ± 7.1 b | 11.8 ± 13.3 b | 118.6 ± 59.7 a |
| HF | Helicotylenchus | 8.7 ± 7.7 a | 8.5 ± 14.7 a | 0 ± 0 a | 67.9 ± 48.3 a | 0 ± 0 a | 74.3 ± 49.3 a |
| HF | Criconema | 0 ± 0 b | 19.6 ± 17.3 ab | 92.7 ± 68.8 a | 10.9 ± 9.4 ab | 17.8 ± 17.3 ab | 5.0 ± 8.7 b |
| OP | Prodorylamus | 13.7 ± 11.9 a | 3.4 ± 5.9 a | 0 ± 0 a | 50.5 ± 31.8 a | 36.5 ± 39.6 a | 34.8 ± 38.1 a |
| BF | Plectus | 2.4 ± 4.2 | 11.8 ± 13.1 | 8.3 ± 8.8 | 26.5 ± 11.4 | 8.5 ± 3.1 | 69.4 ± 71.1 |
| BF | Protorhabditis | 0 ± 0 a | 13.8 ± 23.9 a | 39.6 ± 47.6 a | 16.4 ± 16.5 a | 5.0 ± 1.0 a | 33.8 ± 28.4 a |
| BF | Prismatolainus | 8.7 ± 7.7 b | 0 ± 0 b | 11.8 ± 7.4 b | 13.8 ± 23.9 b | 0 ± 0 b | 69.8 ± 32.2 a |
| HF | Rotylenchulus | 1.6 ± 2.8 b | 1.7 ± 2.9 b | 4.8 ± 8.3 b | 22.5 ± 10.1 ab | 0 ± 0 b | 68.4 ± 44.1 a |
| HF | Tylenchorhynchus | 8.2 ± 14.1 a | 6.9 ± 11.9 a | 0 ± 0 a | 31.2 ± 32.3 a | 3.6 ± 6.3 a | 47.6 ± 43.4 a |
| FF | Oriverutus | 0 ± 0 b | 6.9 ± 11.9 ab | 13.0 ± 5.4 ab | 9.0 ± 15.5 ab | 0 ± 0 b | 66.7 ± 50.4 a |
| OP | Mesodorylaimus | 7.4 ± 6.4 a | 27.1 ± 18.3 a | 0 ± 0 a | 6.5 ± 5.7 a | 0 ± 0 a | 42.1 ± 38.5 a |
| FF | Tylencholaimus | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 67.5 ± 99.1 a |
| BF | Acrobeloides | 3.8 ± 6.6 a | 6.9 ± 11.9 a | 8.3 ± 8.8 a | 9.0 ± 15.5 a | 17.1 ± 6.2 a | 17.2 ± 15.5 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, K.; Qiang, Z.; Hu, Z.; Cheng, S.; Sun, R.; Fang, H.; Zhang, Z.; Ma, C. Elevational Gradients of Soil Nematode Communities in Subtropical Forest Ecosystems. Forests 2024, 15, 2149. https://doi.org/10.3390/f15122149
Ding K, Qiang Z, Hu Z, Cheng S, Sun R, Fang H, Zhang Z, Ma C. Elevational Gradients of Soil Nematode Communities in Subtropical Forest Ecosystems. Forests. 2024; 15(12):2149. https://doi.org/10.3390/f15122149
Chicago/Turabian StyleDing, Kexin, Zhenyu Qiang, Zhengkun Hu, Saisai Cheng, Ruibo Sun, Heng Fang, Zhen Zhang, and Chao Ma. 2024. "Elevational Gradients of Soil Nematode Communities in Subtropical Forest Ecosystems" Forests 15, no. 12: 2149. https://doi.org/10.3390/f15122149
APA StyleDing, K., Qiang, Z., Hu, Z., Cheng, S., Sun, R., Fang, H., Zhang, Z., & Ma, C. (2024). Elevational Gradients of Soil Nematode Communities in Subtropical Forest Ecosystems. Forests, 15(12), 2149. https://doi.org/10.3390/f15122149