
Study on the Response of Vegetation Water Use Efficiency to Drought in the Manas River Basin, Xinjiang, China

Abstract
1. Introduction
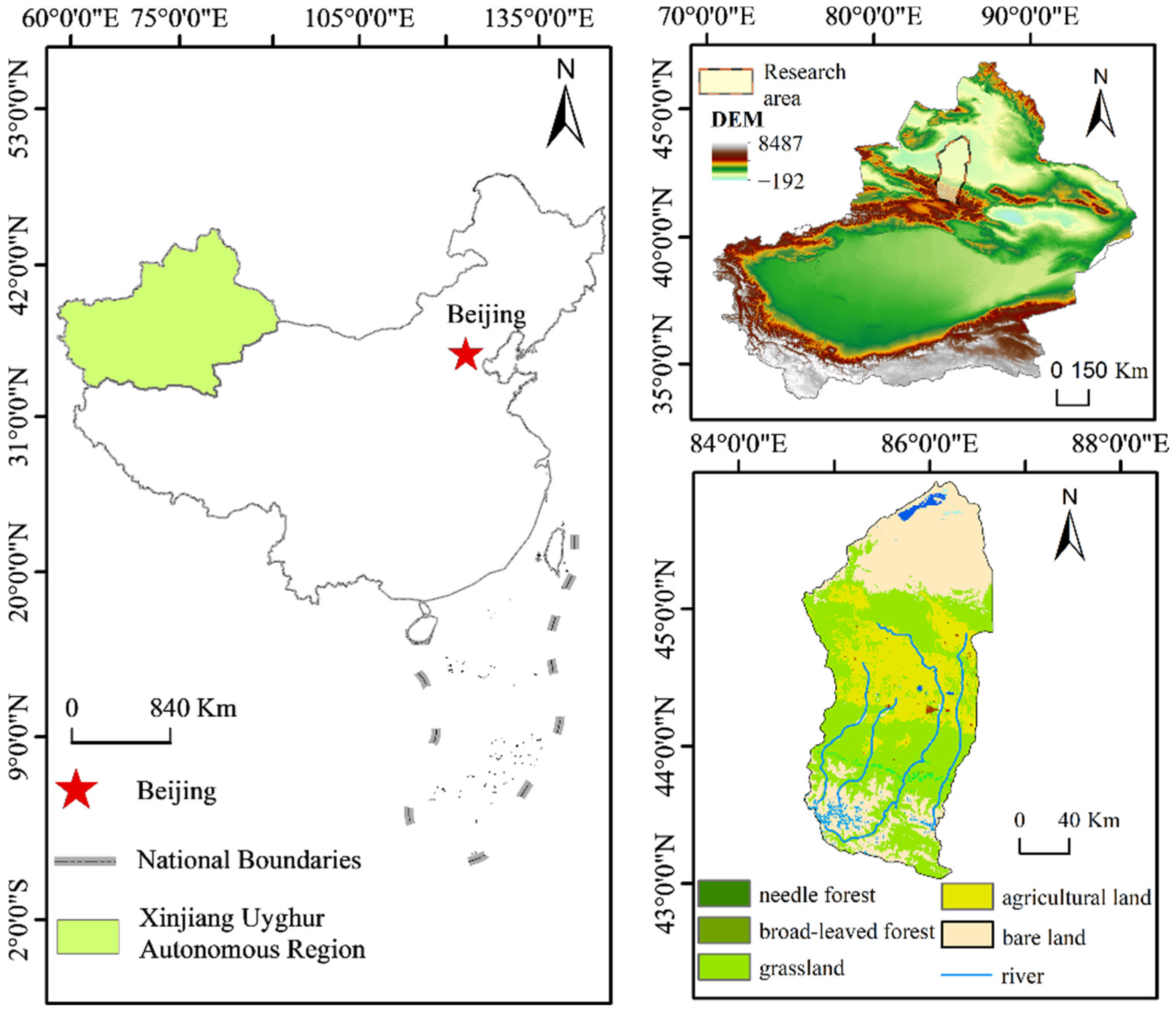
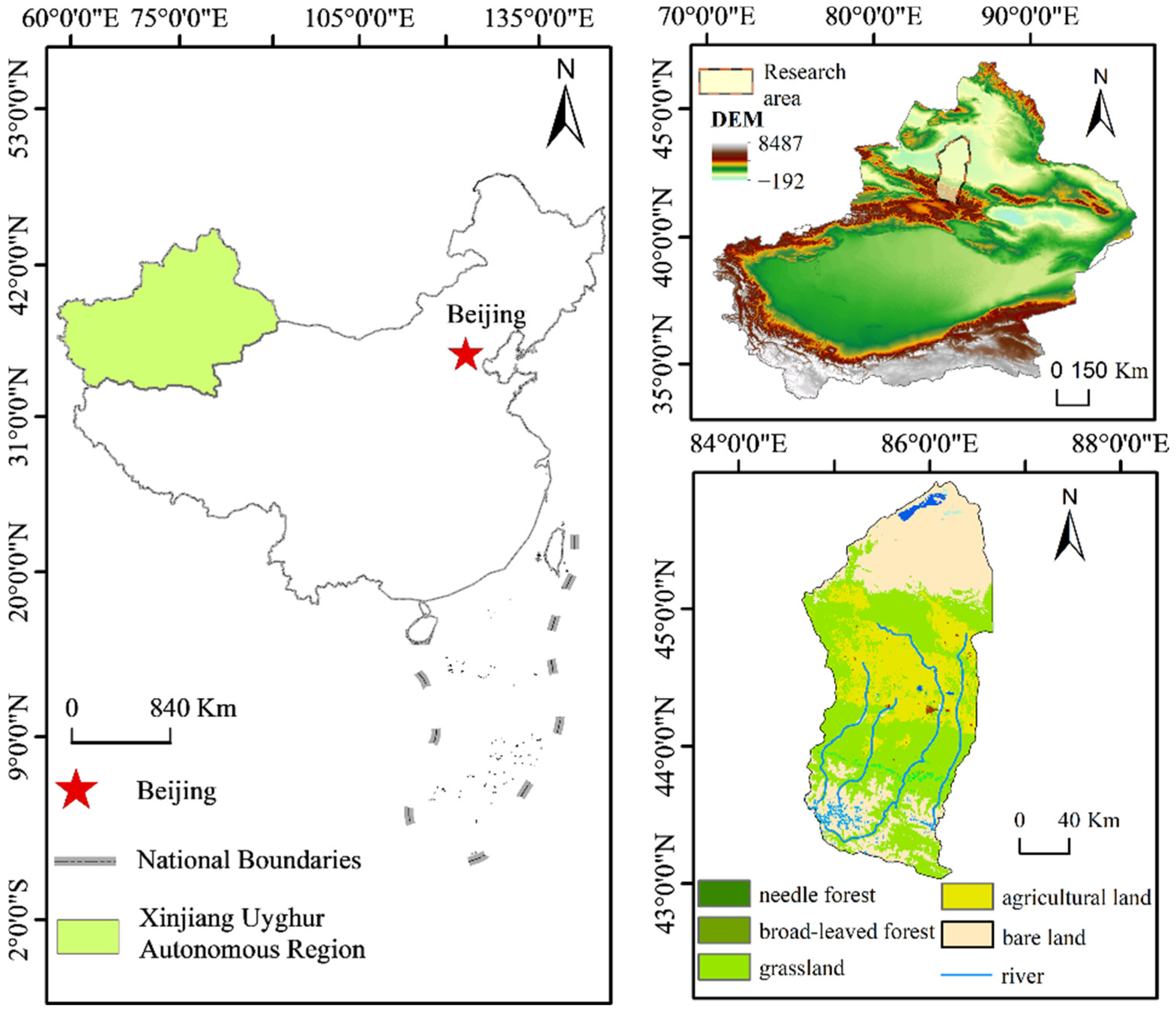
2. Overview of the Study Area
3. Research Methodology
3.1. Data Sources and Preprocessing
3.2. Research Methodology
3.2.1. Estimation of WUE
3.2.2. SPEI Calculation and Classification
3.2.3. Drought Frequency
3.2.4. Standardized Anomaly Index
3.2.5. Pearson’s Correlation Coefficient Method
3.2.6. Mutation Testing
3.2.7. Resilience Analysis
4. Results and Analyses
4.1. Characteristics of Spatial and Temporal Distributions of WUE
4.1.1. Temporal Distribution Characteristics of WUE
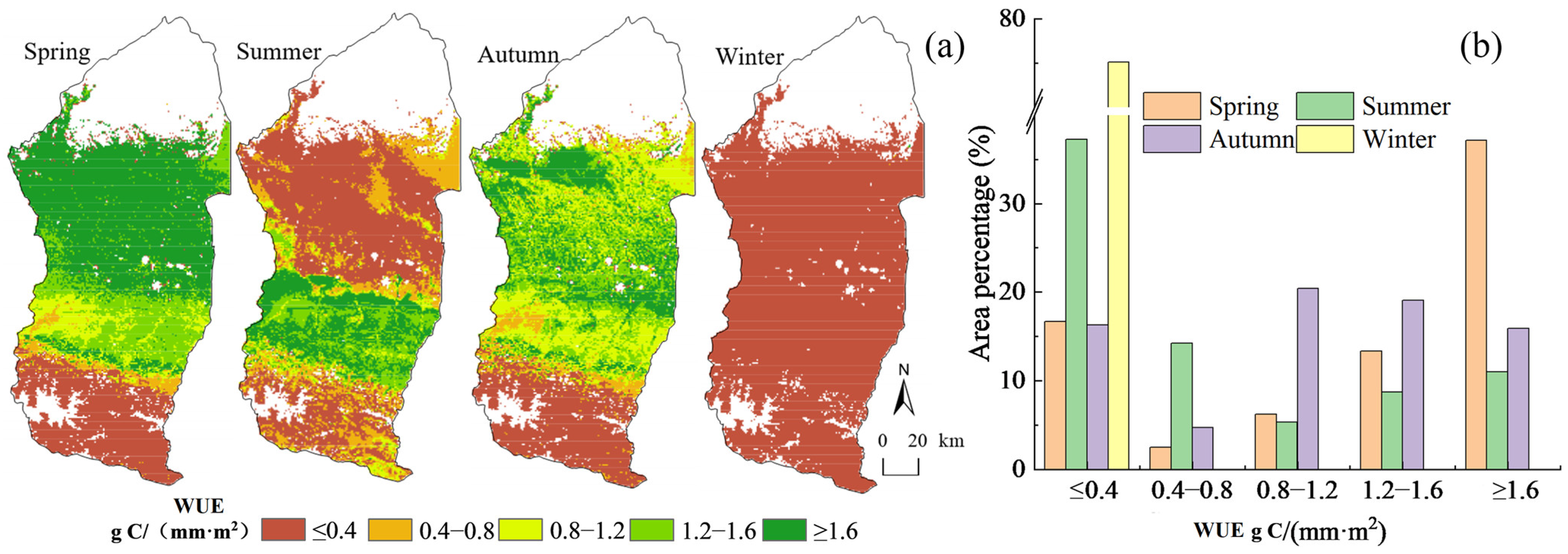
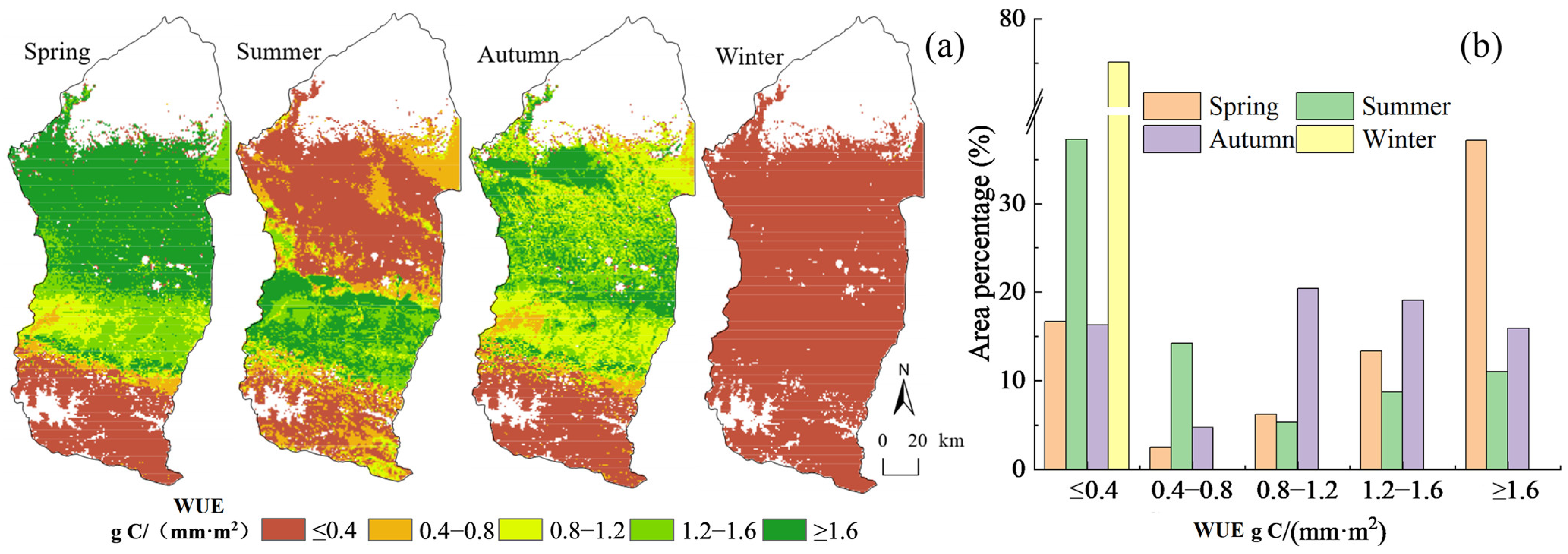
4.1.2. Spatial Distribution Characteristics of WUE
4.2. Characteristics of Spatial and Temporal Distributions of the Aridity Index
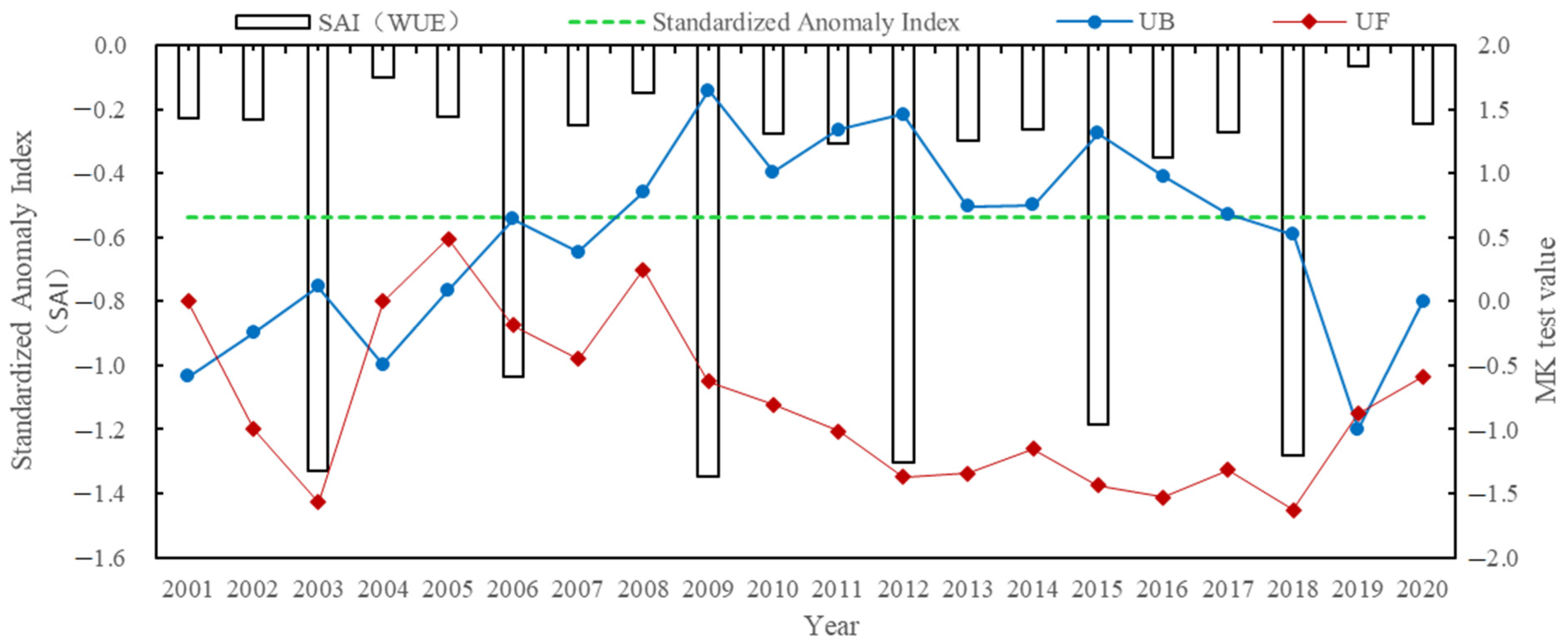
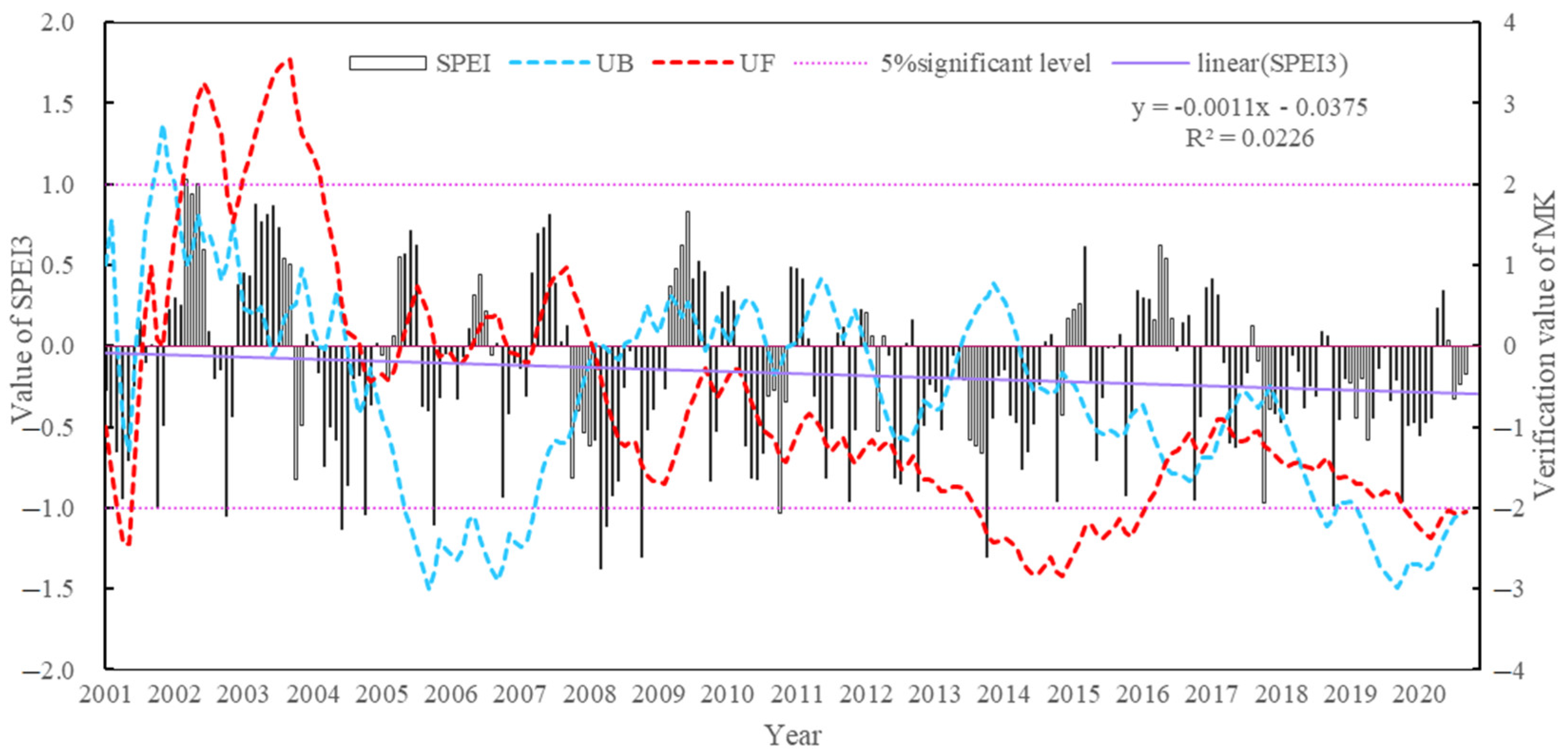
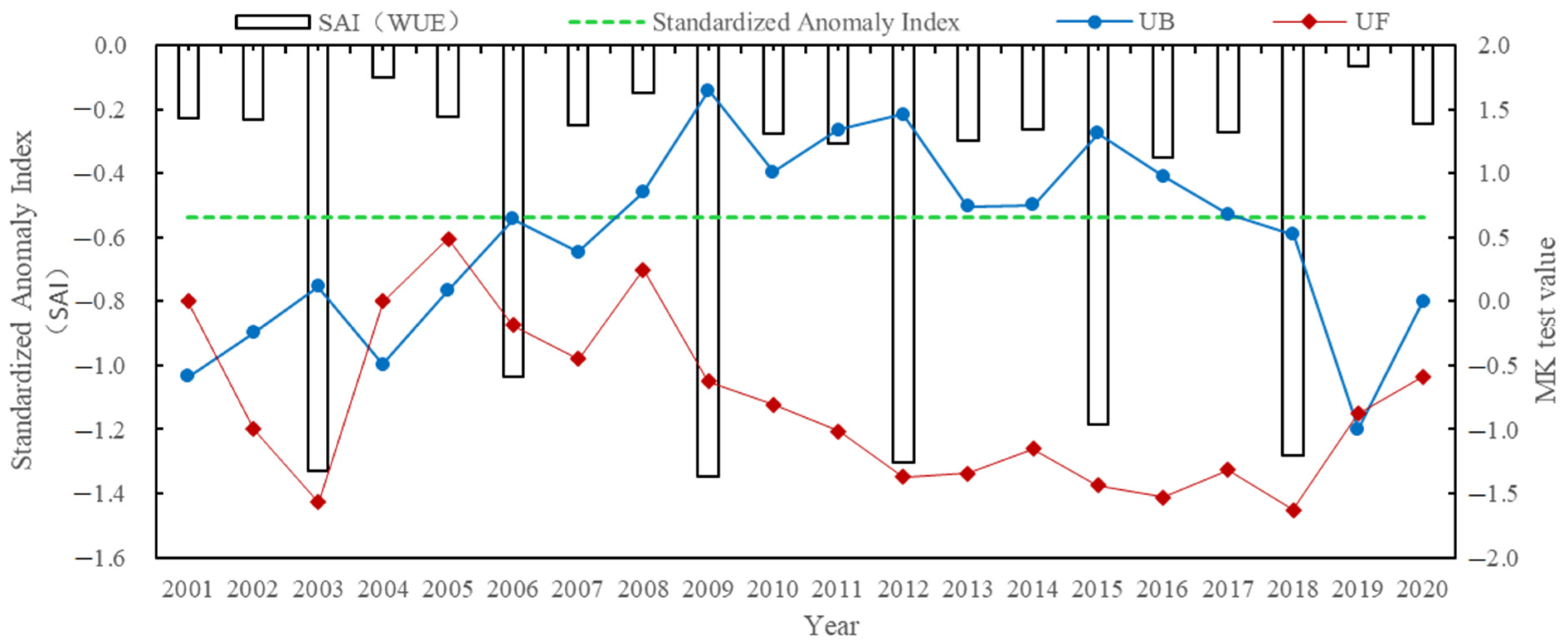
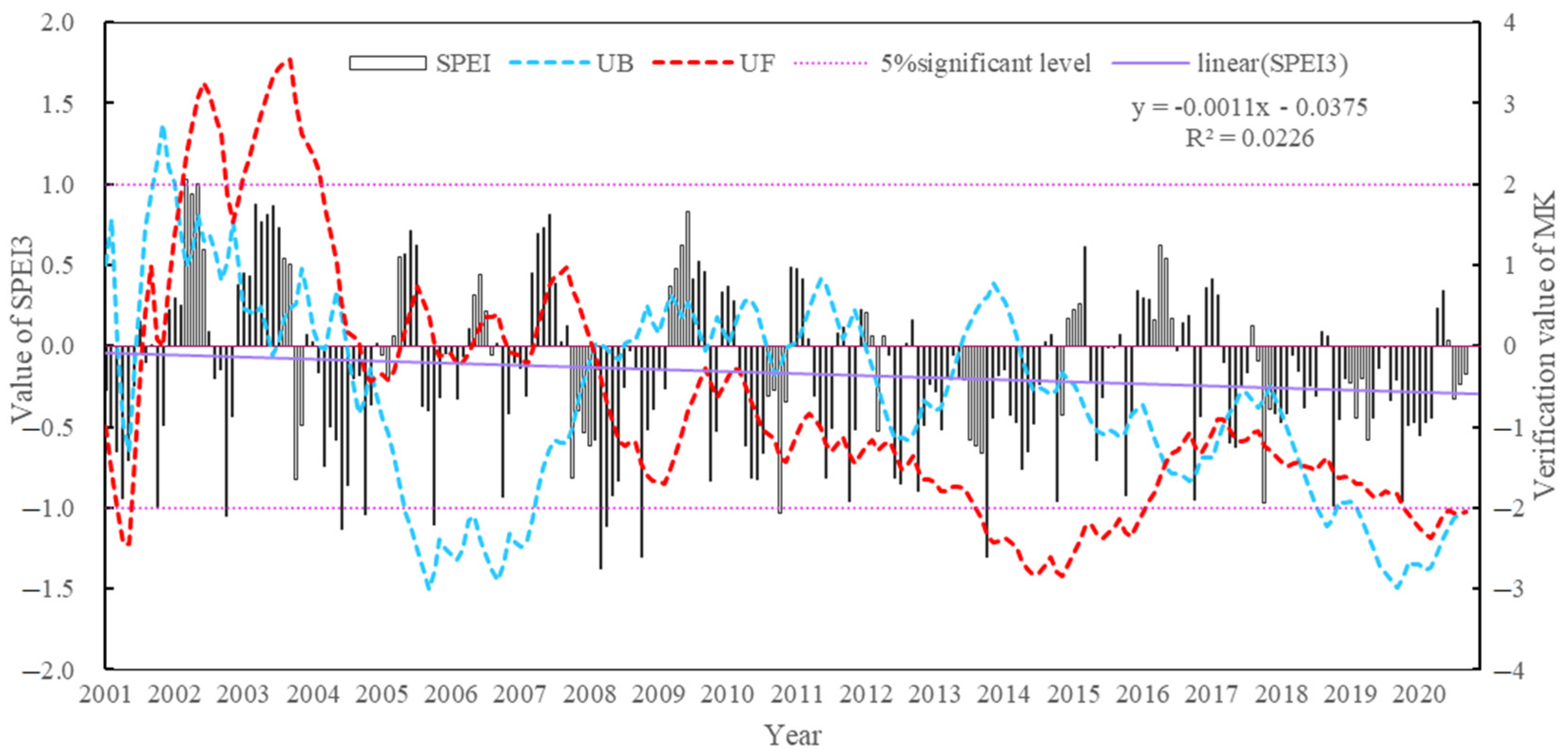
4.2.1. Temporal Distribution Characteristics of the Aridity Index
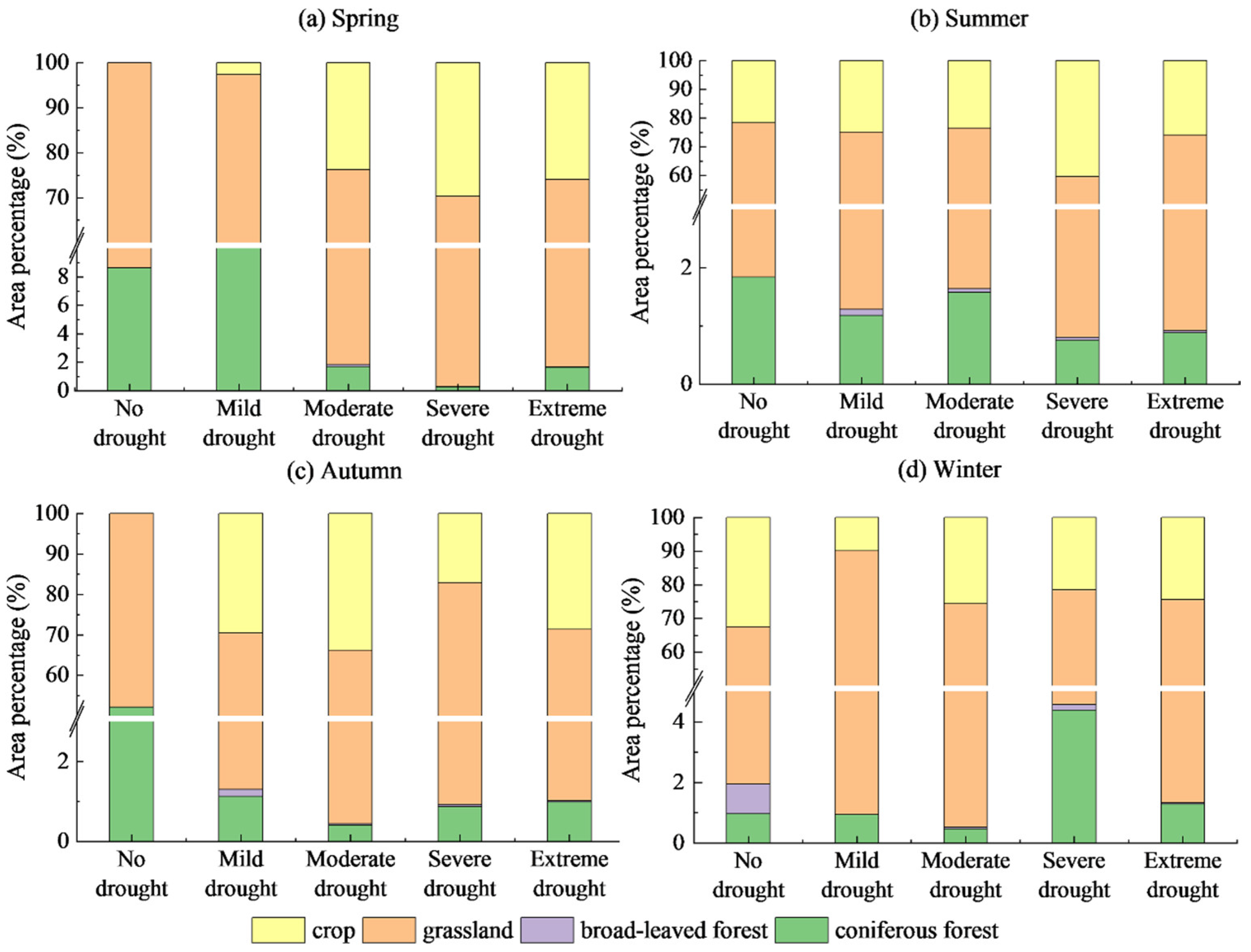
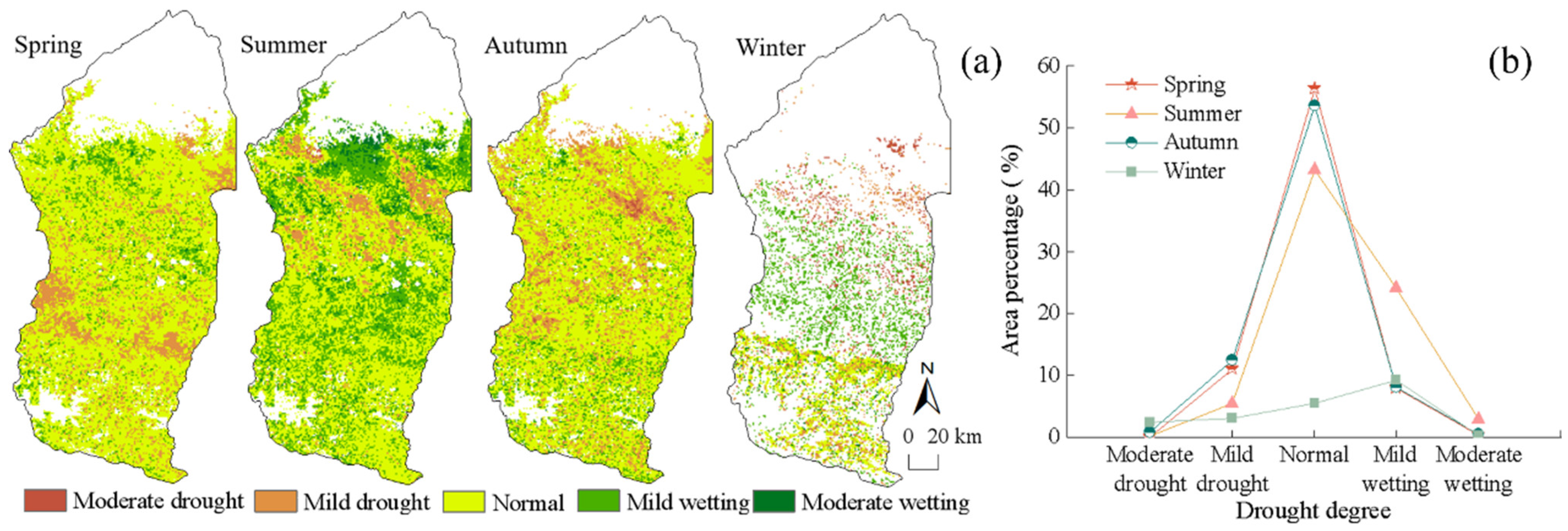
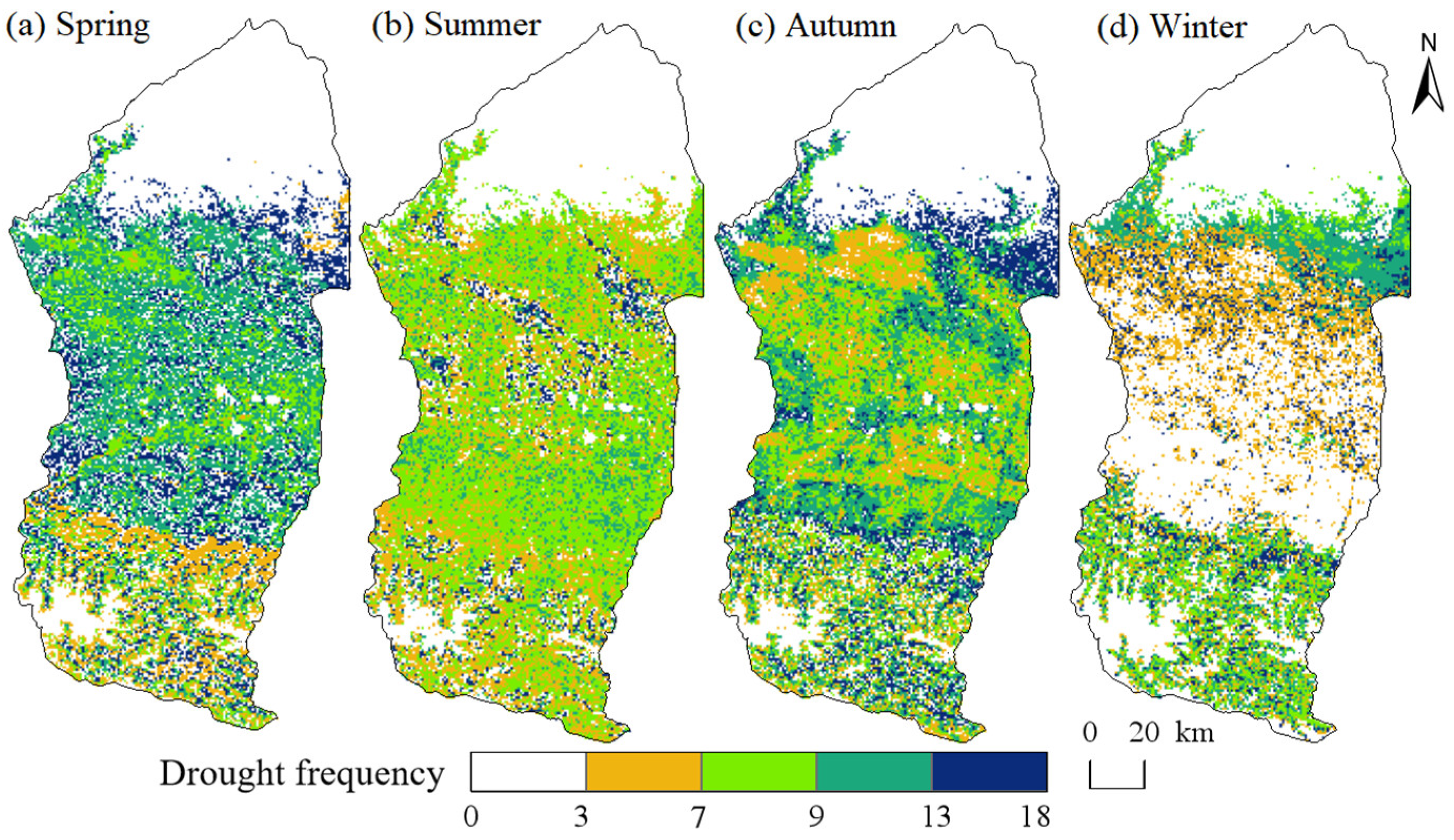
4.2.2. Spatial Distribution Characteristics of Drought Levels
4.3. WUE Response to Drought
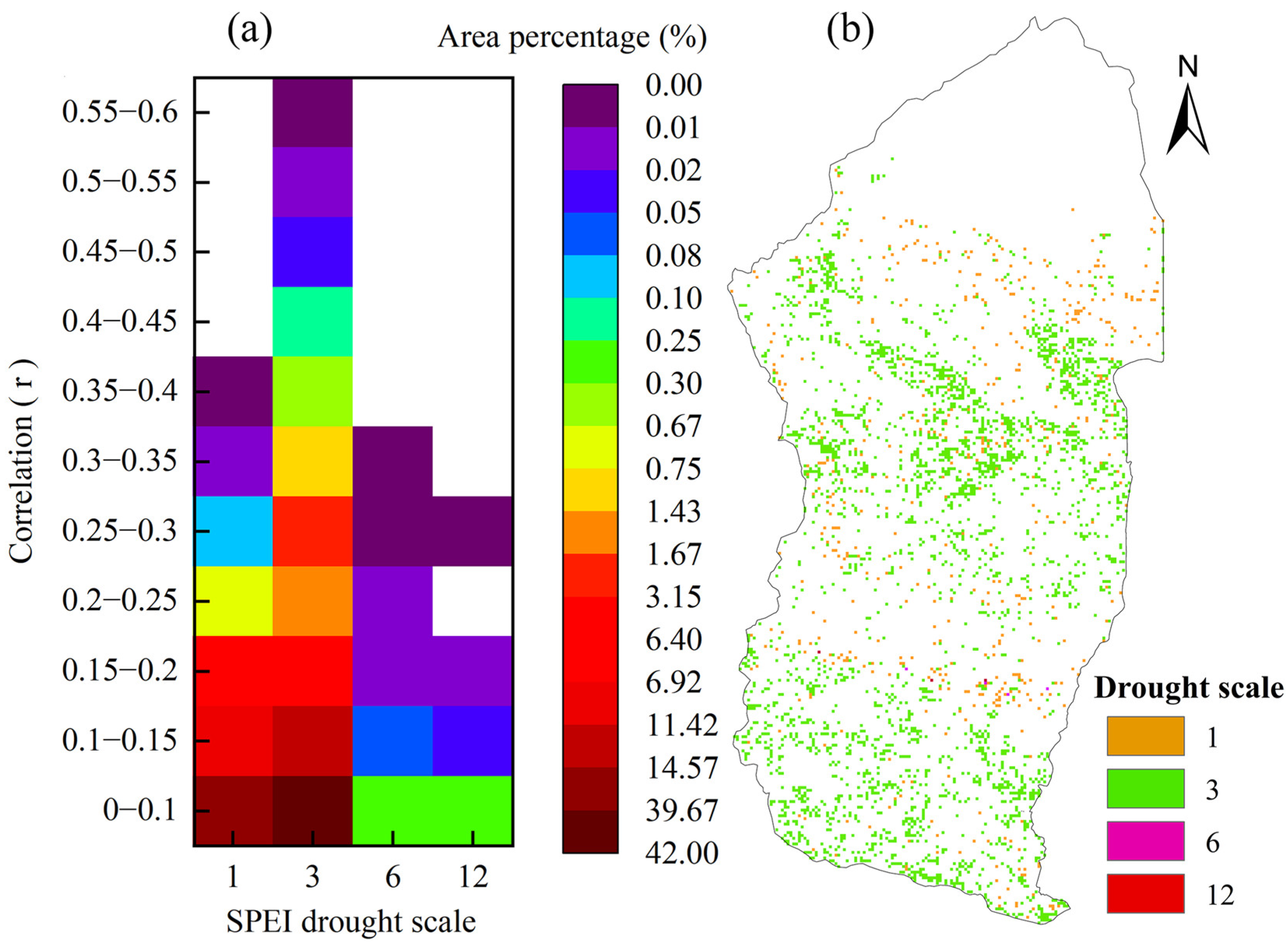
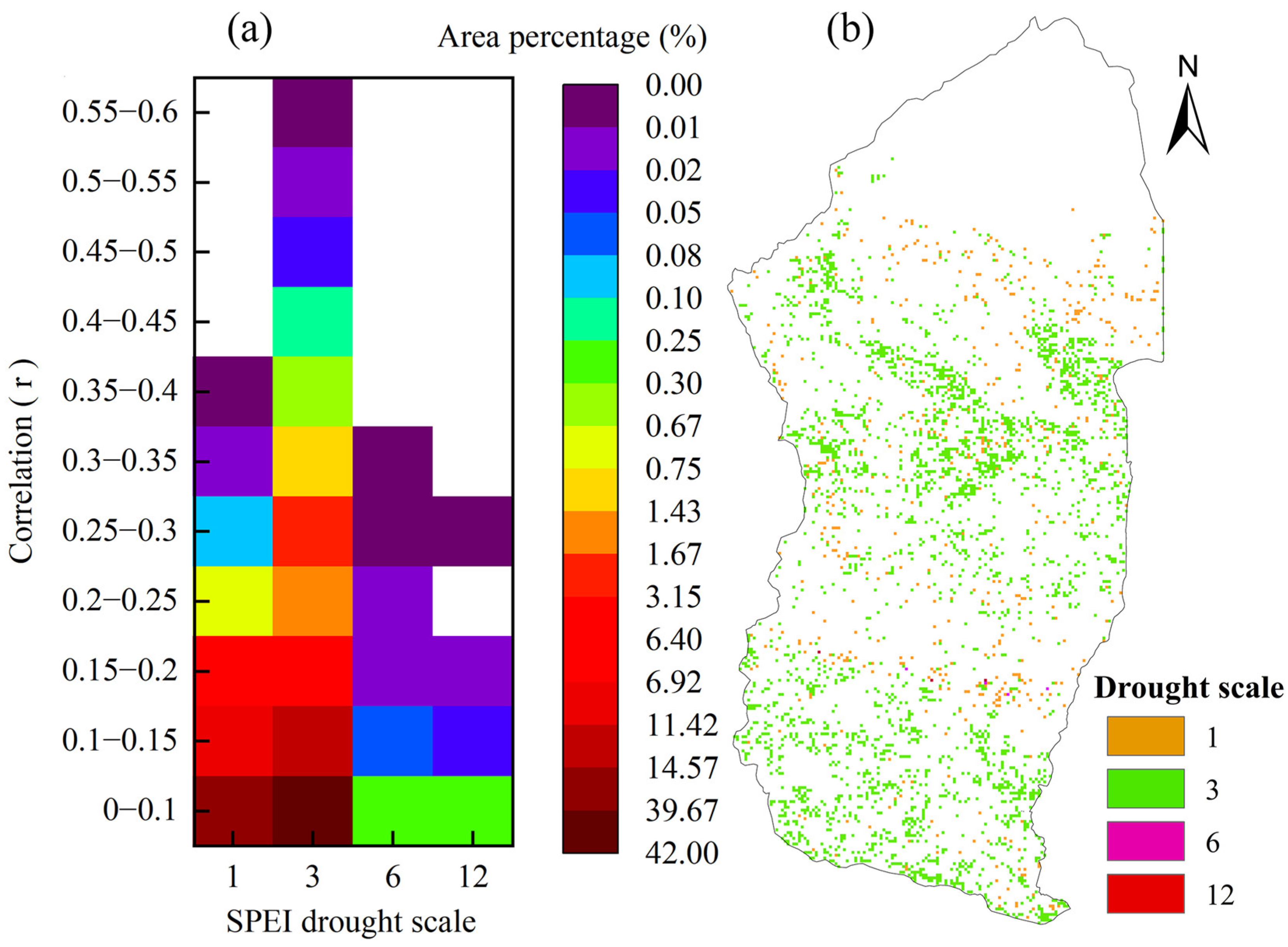
4.3.1. Hysteresis Effects of WUE on Drought
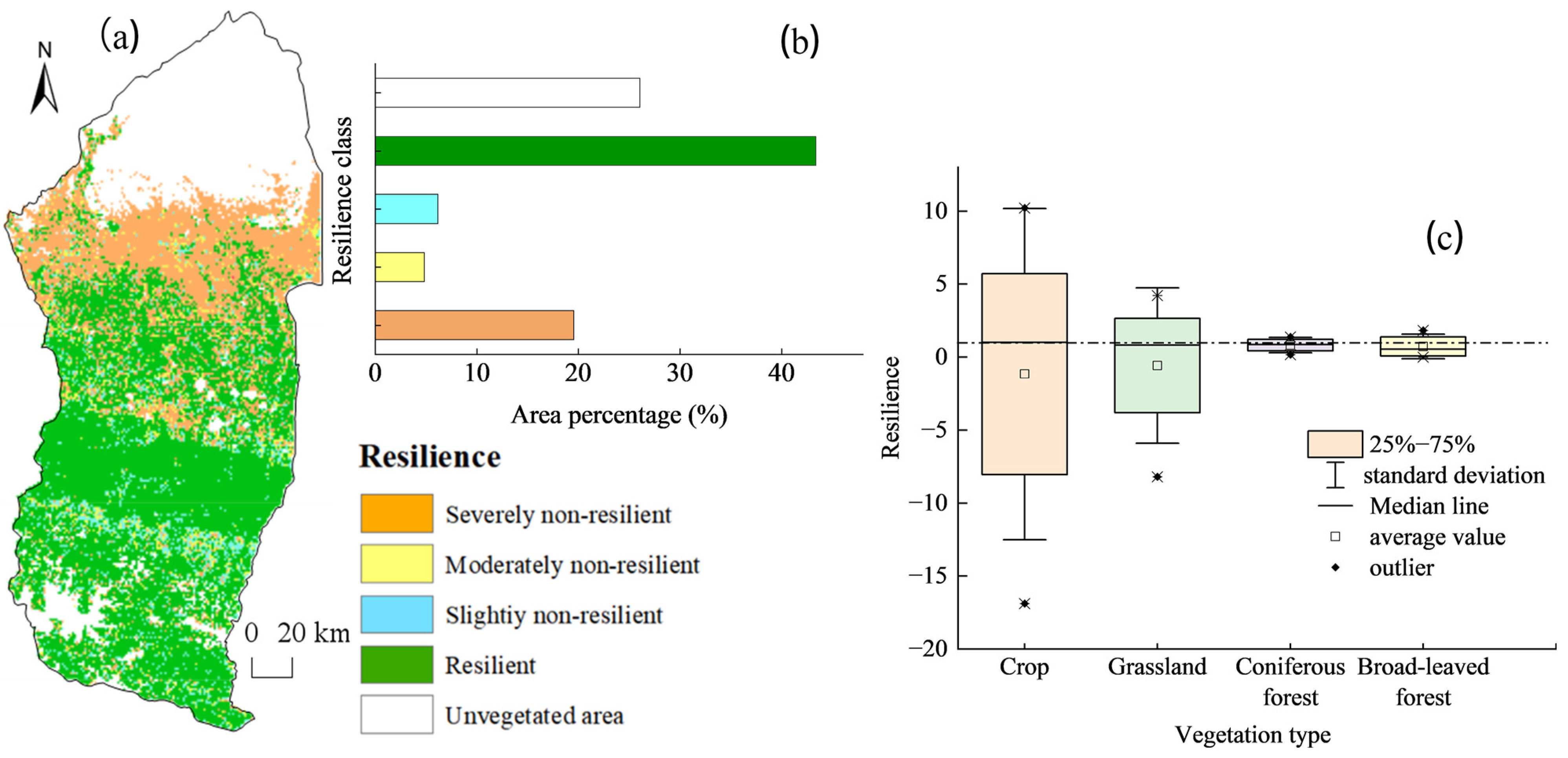
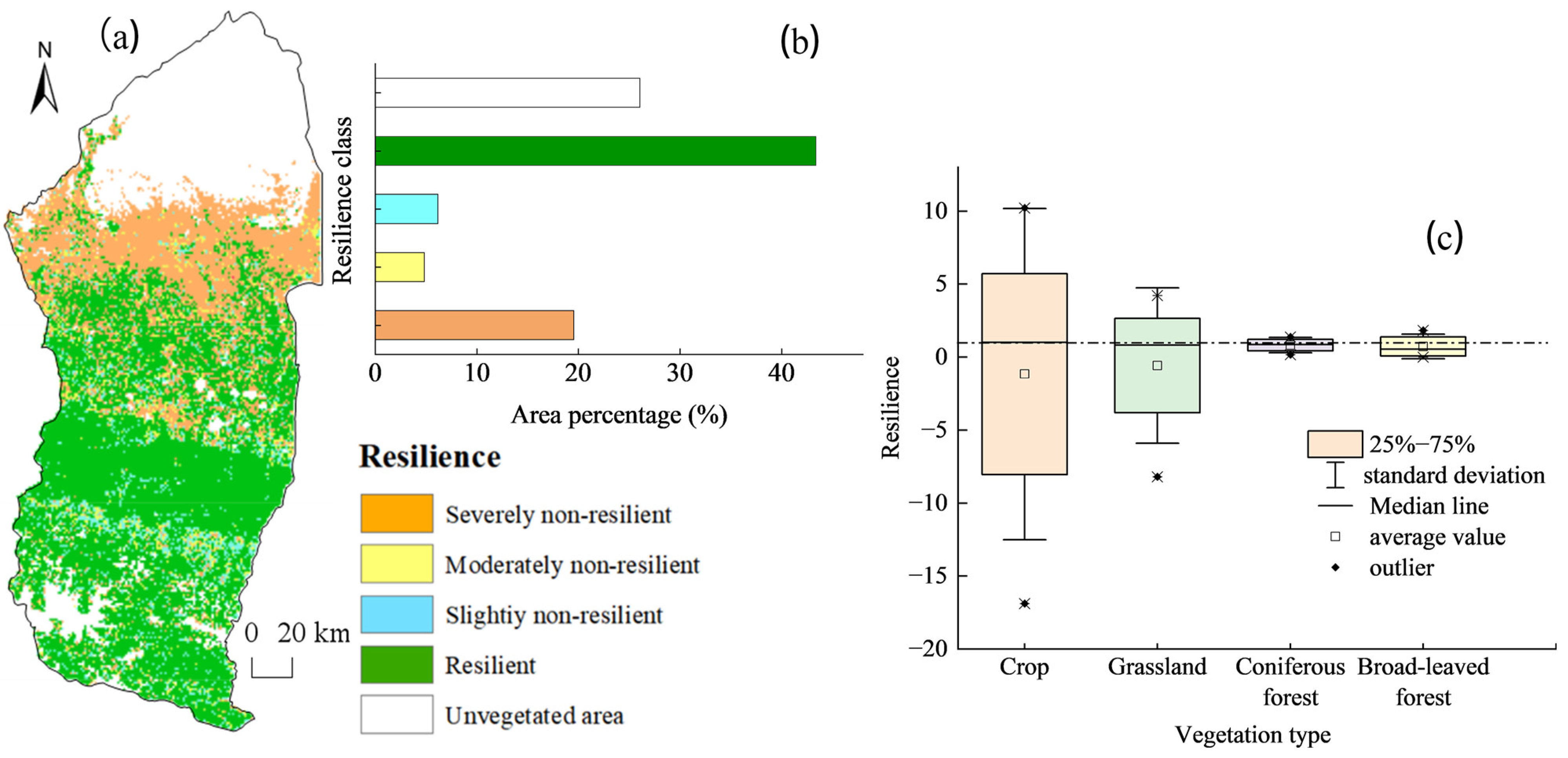
4.3.2. WUE Resilience Response to Drought
5. Discussion
5.1. Analysis of the Causes of Spatial and Temporal Drought Characteristics in the Study Area
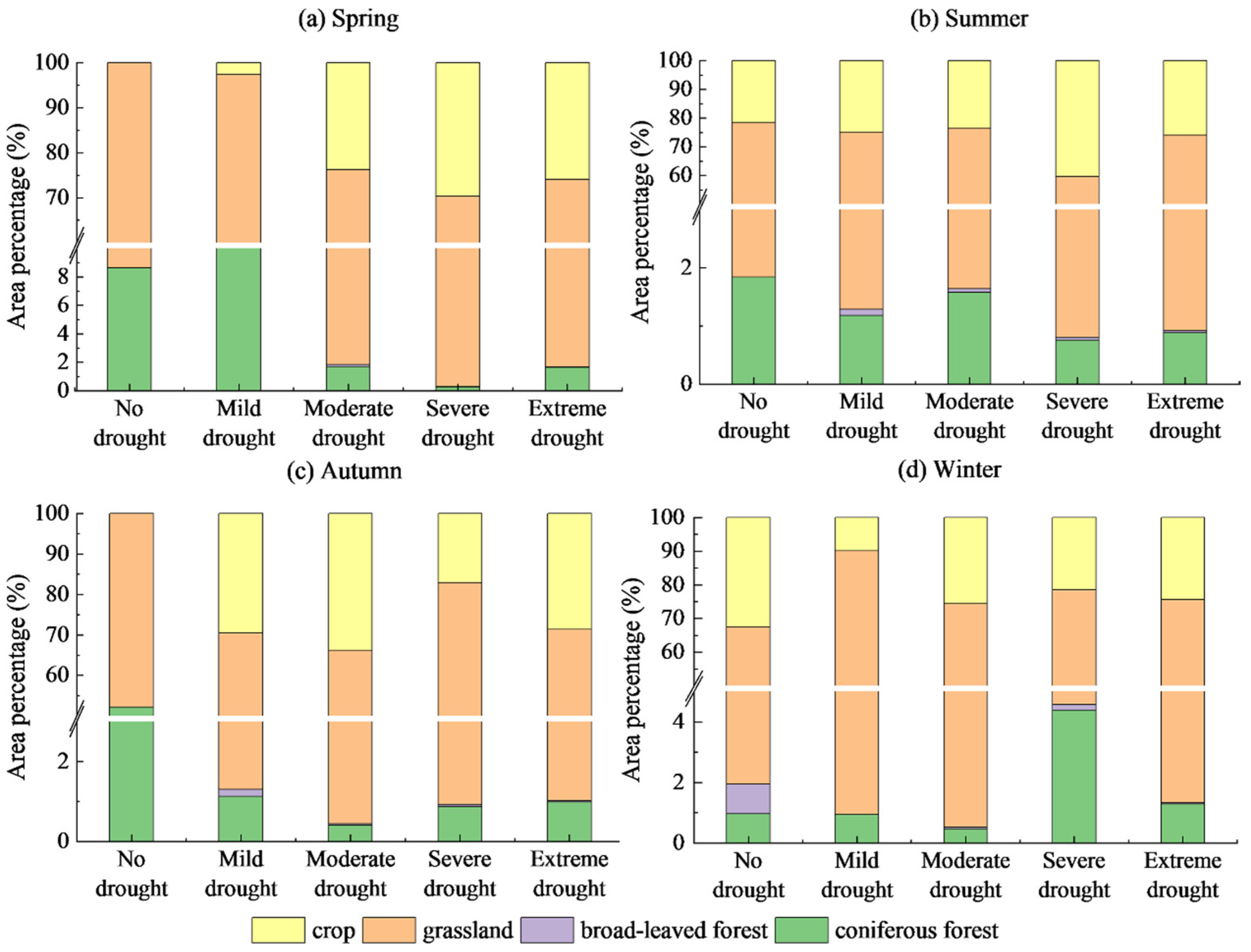
5.2. Differences in WUE Responses to Drought across Vegetation Types
6. Conclusions
- (1)
- The Manas River Basin was at a normal drought level according to the drought index classification during the study period, and the drought trends changed abruptly in 2002, 2005, 2008, and 2016. In the seasonal average SPEI drought category over the past 20 years, most areas were in the normal or slightly wet category, and a few areas were in the slightly dry category. The seasonal drought frequency was relatively high, with most areas experiencing more than seven droughts. Drought occurred more frequently during the growing season for vegetation, particularly during the growing season for grasslands, and less frequently for woodlands.
- (2)
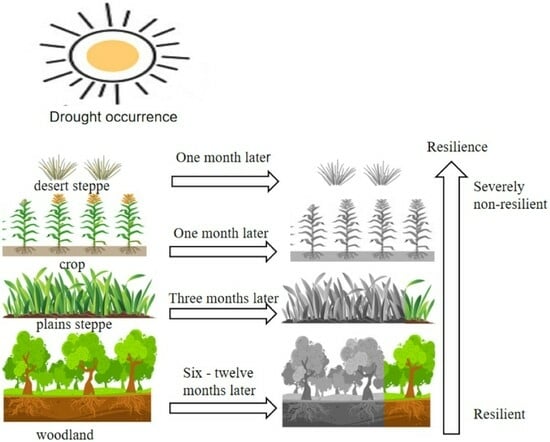
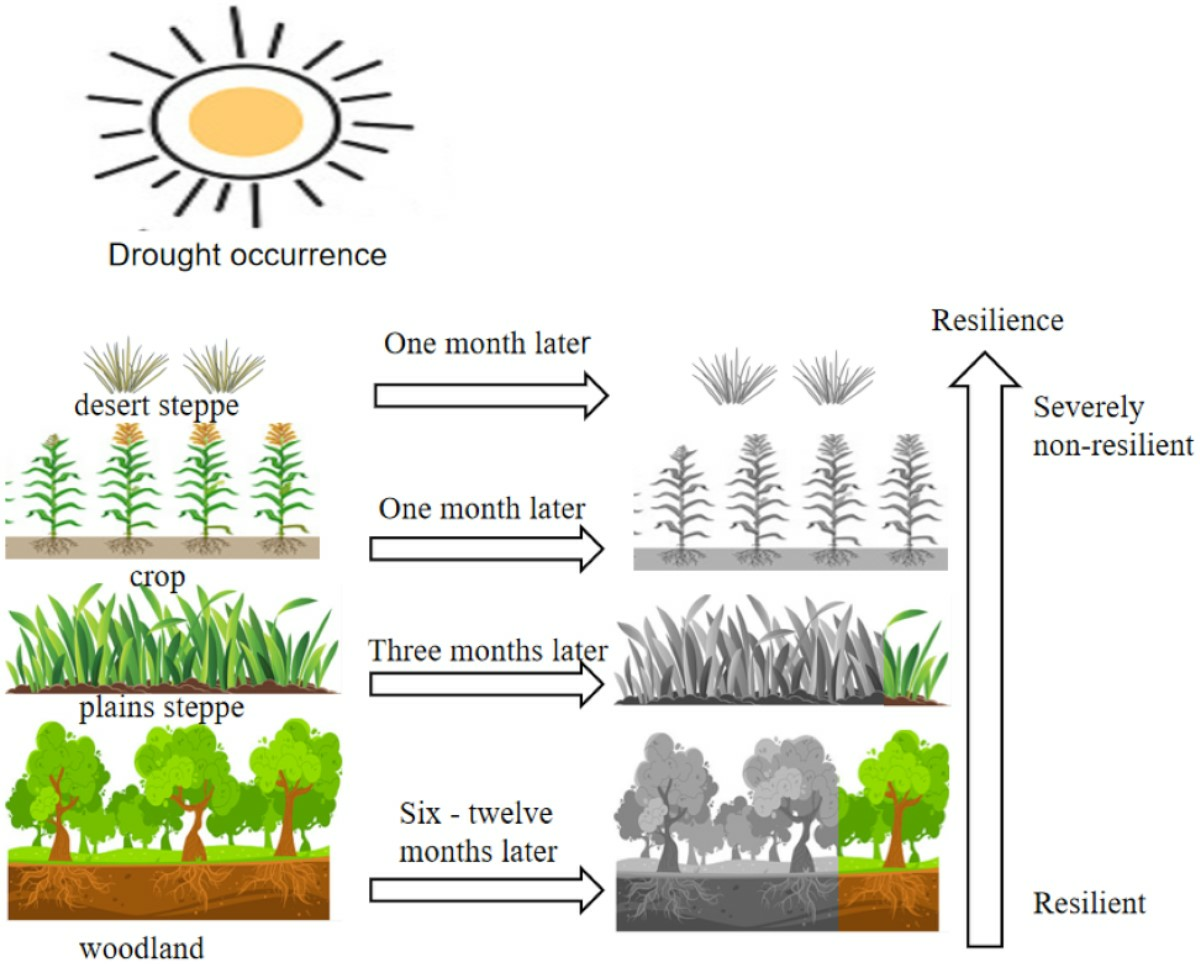
- The vegetation WUE in the Xinjiang Manas Basin showed a significant lag to drought. The average lag time was three months, accounting for 64.0% of the total basin area. The effect of drought on grassland WUE generally occurred in the same month, whereas the length of the lag time for the effect on cropland and grassland WUE in the upper watershed reached 3 months, and the lag time of woodland WUE to drought reached 6–12 months. Forests with high biomass and well-developed root systems can store nutrients such as carbohydrates and have higher resistance to drought and WUE utilisation.
- (3)
- Approximately 38.16% of the regional vegetation ecosystems in the Manas River Basin exhibit drought resistance. Southern mountain ecosystems have the strongest resistance to drought, whereas the grasslands in the northern region, which depend on deserts, have weaker resistance. The closer the area was to the upper part of the basin, the more resilient the vegetation was to drought, and the closer the area was to bare land, the less resilient the vegetation. The resistance of vegetation to drought also depends on the complexity of the ecosystem and the spatial and temporal distribution of water resources.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Amar, G.; Mamtimin, A.; Wang, Y.; Wang, Y.; Gao, J.; Yang, F.; Song, M.; Aihaiti, A.; Wen, C.; Liu, J. Factors controlling and variations of CO2 fluxes during the growing season in Gurbantunggut Desert. Ecol. Indic. 2023, 154, 110708. [Google Scholar] [CrossRef]
- Guo, H.; Bao, A.; Liu, T.; Ndayisaba, F.; Jiang, L.; Zheng, G.; Chen, T.; De Maeyer, P. Determining variable weights for an Optimal Scaled Drought Condition Index (OSDCI): Evaluation in Central Asia. Remote Sens. Environ. 2019, 231, 111220. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, Q.; Xu, C.-Y.; Du, J.; Sun, P.; Hu, P. Modified palmer drought severity index: Model improvement and application. Environ. Int. 2019, 130, 104951. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Adu, B.; Wu, J.; Qin, G.; Li, H.; Han, Y. Spatial and temporal variations of drought in Sichuan Province from 2001 to 2020 based on modified temperature vegetation dryness index (TVDI). Ecol. Indic. 2022, 139, 108883. [Google Scholar] [CrossRef]
- Yang, R.; Xing, B. Teleconnections of large-scale climate patterns to regional drought in mid-latitudes: A case study in Xinjiang, China. Atmosphere 2022, 13, 230. [Google Scholar] [CrossRef]
- Liu, Y.; Zheng, J.; Guan, J.; Han, W.; Liu, L. Concurrent and lagged effects of drought on grassland net primary productivity: A case study in Xinjiang, China. Front. Ecol. Evol. 2023, 11, 1131175. [Google Scholar] [CrossRef]
- Wang, M.; Ding, Z.; Wu, C.; Song, L.; Ma, M.; Yu, P.; Lu, B.; Tang, X. Divergent responses of ecosystem water-use efficiency to extreme seasonal droughts in Southwest China. Sci. Total. Environ. 2021, 760, 143427. [Google Scholar] [CrossRef]
- Zhao, T.; Zhang, Y.; Zhang, T.; Xu, M.; Zhu, J.; He, Y.; Yu, G. Drought occurrence and time-dominated variations in water use efficiency in an alpine meadow on the Tibetan Plateau. Ecohydrology 2021, 15, e2360. [Google Scholar] [CrossRef]
- Liu, P.; Zha, T.; Jia, X.; Black, T.A.; Jassal, R.S.; Ma, J.; Bai, Y.; Wu, Y. Different effects of spring and summer droughts on ecosystem carbon and water exchanges in a semiarid shrubland ecosystem in northwest China. Ecosystems 2019, 22, 1869–1885. [Google Scholar] [CrossRef]
- Han, W.; Guan, J.; Zheng, J.; Liu, Y.; Ju, X.; Liu, L.; Li, J.; Mao, X.; Li, C. Probabilistic assessment of drought stress vulnerability in grasslands of Xinjiang, China. Front. Plant Sci. 2023, 14, 1143863. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, W.; Schwalm, C.R.; Gentine, P.; Smith, W.K.; Ciais, P.; Kimball, J.S.; Gazol, A.; Kannenberg, S.A.; Chen, A.; et al. Widespread spring phenology effects on drought recovery of Northern Hemisphere ecosystems. Nat. Clim. Chang. 2023, 13, 182–188. [Google Scholar] [CrossRef]
- Bai, Y.; Liu, M.; Guo, Q.; Wu, G.; Wang, W.; Li, S. Diverse responses of gross primary production and leaf area index to drought on the Mongolian Plateau. Sci. Total. Environ. 2023, 902, 166507. [Google Scholar] [CrossRef] [PubMed]
- Zhan, C.; Liang, C.; Zhao, L.; Jiang, S.; Niu, K.; Zhang, Y. Drought-related cumulative and time-lag effects on vegetation dynamics across the Yellow River Basin, China. Ecol. Indic. 2022, 143, 109409. [Google Scholar] [CrossRef]
- Ahmadi, B.; Moradkhani, H. Revisiting hydrological drought propagation and recovery considering water quantity and quality. Hydrol. Process. 2019, 33, 1492–1505. [Google Scholar] [CrossRef]
- Ji, Y.; Li, Y.; Yao, N.; Biswas, A.; Zou, Y.; Meng, Q.; Liu, F. The lagged effect and impact of soil moisture drought on terrestrial ecosystem water use efficiency. Ecol. Indic. 2021, 133, 108349. [Google Scholar] [CrossRef]
- Wang, W.; Li, J.; Qu, H.; Xing, W.; Zhou, C.; Tu, Y.; He, Z. Spatial and temporal drought characteristics in the Huanghuaihai Plain and its influence on cropland water use efficiency. Remote Sens. 2022, 14, 2381. [Google Scholar] [CrossRef]
- Li, J.J.; Luo, G.P.; Ding, J.L.; Xu, W.Q.; Zheng, S.L. Impacts of the progress of artificial irrigation and drainage technology on the change of cropland pattern In the Manas Basin of Xinjiang in the last 50a. J. Nat. Resour. 2016, 31, 570–582. [Google Scholar]
- Qin, J.L.; Xue, L.Q. Characteristics of spatial and temporal variation of vegetation and its spatial relationship with topographic factors In the Manas Basin of Xinjiang, Northwest Arid Zone. J. Ecol. Environ. 2020, 29, 2179–2188. [Google Scholar] [CrossRef]
- Chen, F.L.; Yang, K.; Cai, W.J.; Long, A.H.; He, X.L. esearch on hydrological aridity index based on gamlss model—A case study of Manas River Basin. Geogr. Publ. Res. 2021, 40, 2670–2683. [Google Scholar]
- Tirivarombo, S.; Osupile, D.; Eliasson, P. Drought monitoring and analysis: Standardised precipitation evapotranspiration index (SPEI) and standardised precipitation index (SPI). Phys. Chem. Earth Parts A/B/C 2018, 106, 1–10. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, X.L.; Li, J.J.; Wang, M.T.; Ma, Z.L. Characterisation of spatial and temporal variations of seasonal drought in the basin area of Sichuan Province based on SPEI. Agric. Res. Arid Reg. 2018, 36, 242–250+256. [Google Scholar]
- Zhao, H.; Yao, J.Q.; Li, X.G.; Tao, H. Characterisation of wet and dry climate change in Xinjiang. J. Sun Yat-Sen Univ. Nat. Sci. Ed. 2020, 5, 126–133. [Google Scholar] [CrossRef]
- Zhang, T.; Zhou, J.; Yu, P.; Li, J.; Kang, Y.; Zhang, B. Response of ecosystem gross primary productivity to drought in northern China based on multi-source remote sensing data. J. Hydrol. 2023, 616, 128808. [Google Scholar] [CrossRef]
- Huang, Y.F.; Ng, J.L.; Fung, K.F.; Weng, T.K.; AlDahoul, N.; Ahmed, A.N.; Sherif, M.; Chaplot, B.; Chong, K.L.; Elshafie, A. Space–time heterogeneity of drought characteristics in Sabah and Sarawak, East Malaysia: Implications for developing effective drought monitoring and mitigation strategies. Appl. Water Sci. 2023, 13, 1–25. [Google Scholar] [CrossRef]
- Zhao, H.; Li, X.; Eziz, M.; Yao, J.Q. Changes in the characteristics of dry-wet periods in Xinjiang, China based on the SPEI index. Atmósfera 2022, 35, 483–496. [Google Scholar]
- Abdurehman, A.; Mei, Z. Characterisation of drought change in Xinjiang based on SPEI. Anhui Agric. Sci. 2022, 50, 178–183. [Google Scholar]
- Liu, Y.B.; Wang, H.Q.; Ju, W.M. Characteristics of drought impact on forest productivity in Jiangxi Province. Nat. Hazards 2016, 25, 67–77. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Gouveia, C.; Camarero, J.J.; Beguería, S.; Trigo, R.; López-Moreno, J.I.; Azorin-Molina, C.; Pasho, E.; Lorenzo-Lacruz, J.; Revuelto, J.; et al. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef]
- Zhang, Q.; Kong, D.; Singh, V.P.; Shi, P. Response of vegetation to different time-scales drought across China: Spatiotemporal patterns, causes and implications. Glob. Planet. Chang. 2017, 152, 1–11. [Google Scholar] [CrossRef]
- Hamed, K.H. Trend detection in hydrologic data: The Mann–Kendall trend test under the scaling hypothesis. J. Hydrol. 2008, 349, 350–363. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, W. Ecosystem water use efficiency in the three-North Region of China based on long-term satellite data. Sustainability 2021, 13, 7977. [Google Scholar] [CrossRef]
- Sharma, A.; Goyal, M.K. District-level assessment of the ecohydrological resilience to hydroclimatic disturbances and its controlling factors in India. J. Hydrol. 2018, 564, 1048–1057. [Google Scholar] [CrossRef]
- Walker, B.; Holling, C.S.; Carpenter, S.R.; Kinzig, A. Resilience, adaptability and transformability in social-ecological systems. Ecol. Soc. 2004, 9, 5. [Google Scholar] [CrossRef]
- Sharma, A.; Goyal, M.K. Assessment of ecosystem resilience to hydroclimatic disturbances in India. Glob. Chang. Biol. 2018, 24, e432–e441. [Google Scholar] [CrossRef]
- Zhang, H.; Song, J.; Wang, G.; Wu, X.; Li, J. Spatiotemporal characteristic and forecast of drought in northern Xinjiang, China. Ecol. Indic. 2021, 127, 107712. [Google Scholar] [CrossRef]
- Yizhong, W. Xinjiang Uygur Autonomous Region Manas River Basin Water Conservancy Record; Shihezi Water Electr: Shihezi, China, 2002. [Google Scholar]
- Shi, Y.G. Chinese Meteorological Disaster Dictionary: Xinjiang Volume; Meteorological Publishing House: Beijing, China, 2006; pp. 138–152. [Google Scholar]
- Wei, X.; He, W.; Zhou, Y.; Ju, W.; Xiao, J.; Li, X.; Liu, Y.; Xu, S.; Bi, W.; Zhang, X.; et al. Global assessment of lagged and cumulative effects of drought on grassland gross primary production. Ecol. Indic. 2022, 136, 108646. [Google Scholar] [CrossRef]
- Wei, X.; He, W.; Zhou, Y.; Cheng, N.; Xiao, J.; Bi, W.; Liu, Y.; Sun, S.; Ju, W. Increased sensitivity of global vegetation productivity to drought over the recent three decades. J. Geophys. Res. Atmos. 2023, 128, e2022JD037504. [Google Scholar] [CrossRef]
- Fan, Y.; Miguez-Macho, G.; Jobbágy, E.G.; Jackson, R.B.; Otero-Casal, C. Hydrologic regulation of plant rooting depth. Proc. Natl. Acad. Sci. USA 2017, 114, 10572–10577. [Google Scholar] [CrossRef]
- Schwinning, S.; Sala, O.E. Hierarchy of responses to resource pulses in arid and semi-arid ecosystems. Oecologia 2004, 141, 211–220. [Google Scholar] [CrossRef]
- Craine, J.M.; Ocheltree, T.W.; Nippert, J.B.; Towne, E.G.; Skibbe, A.M.; Kembel, S.W.; Fargione, J.E. Global diversity of drought tolerance and grassland climate-change resilience. Nat. Clim. Chang. 2013, 3, 63–67. [Google Scholar] [CrossRef]
- Stocker, B.D.; Tumber-Dávila, S.J.; Konings, A.G.; Anderson, M.C.; Hain, C.; Jackson, R.B. Global patterns of water storage in the rooting zones of vegetation. Nat. Geosci. 2023, 16, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, W.; Li, W.; Zhao, S.; Wang, S.; Liu, T. Assessment of the spatiotemporal characteristics of vegetation water use efficiency in response to drought in Inner Mongolia, China. Environ. Sci. Pollut. Res. 2023, 30, 6345–6357. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Timmermans, J.; Wen, W.; van Bodegom, P.M. Ecosystems threatened by intensified drought with divergent vulnerability. Remote Sens. Environ. 2023, 289, 113512. [Google Scholar] [CrossRef]
- Liu, W.; Su, J.; Li, S.; Lang, X.; Huang, X. Non-structural carbohydrates regulated by season and species in the subtropical monsoon broad-leaved evergreen forest of Yunnan Province, China. Sci. Rep. 2018, 8, 1083. [Google Scholar] [CrossRef]
- Shi, S.; Yu, J.; Wang, F.; Wang, P.; Zhang, Y.; Jin, K. Quantitative contributions of climate change and human activities to vegetation changes over multiple time scales on the Loess Plateau. Sci. Total. Environ. 2021, 755, 142419. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drought Type | Moderate Wet | Mild Wet | Normal | Mild Drought | Moderate Drought | Severe Drought | Extreme Drought |
|---|---|---|---|---|---|---|---|
| SPEI value | 1.0–1.5 | 0.5–1.0 | −0.5–0.5 | −1.0–−0.5 | −1.5–−1.0 | −2.0–−1.5 | ≤−2.0 |
| Drought Frequency | Area Proportion (%) | |||
|---|---|---|---|---|
| Spring | Summer | Autumn | Winter | |
| 0 | 38.51 | 28.04 | 28.59 | 53.60 |
| 3 | 0.59 | 1.34 | 1.74 | 1.26 |
| 7 | 4.83 | 7.51 | 15.22 | 17.39 |
| 9 | 5.32 | 49.96 | 21.00 | 4.97 |
| 13 | 17.21 | 8.89 | 22.37 | 15.38 |
| 18 | 33.55 | 4.26 | 11.08 | 7.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, J.; Zan, M.; Chen, Z.; Xue, C.; Yang, S. Study on the Response of Vegetation Water Use Efficiency to Drought in the Manas River Basin, Xinjiang, China. Forests 2024, 15, 114. https://doi.org/10.3390/f15010114
Kong J, Zan M, Chen Z, Xue C, Yang S. Study on the Response of Vegetation Water Use Efficiency to Drought in the Manas River Basin, Xinjiang, China. Forests. 2024; 15(1):114. https://doi.org/10.3390/f15010114
Chicago/Turabian StyleKong, Jingjing, Mei Zan, Zhizhong Chen, Cong Xue, and Shunfa Yang. 2024. "Study on the Response of Vegetation Water Use Efficiency to Drought in the Manas River Basin, Xinjiang, China" Forests 15, no. 1: 114. https://doi.org/10.3390/f15010114
APA StyleKong, J., Zan, M., Chen, Z., Xue, C., & Yang, S. (2024). Study on the Response of Vegetation Water Use Efficiency to Drought in the Manas River Basin, Xinjiang, China. Forests, 15(1), 114. https://doi.org/10.3390/f15010114