
Genome-Wide Analysis of Alternative Splicing Events Responding to High Temperatures in Populus tomentosa Carr.

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatment
2.2. Physiological-Index Test
2.3. RNA Extraction, cDNA Library Preparation, and RNA-seq
2.4. Identification of AS Events
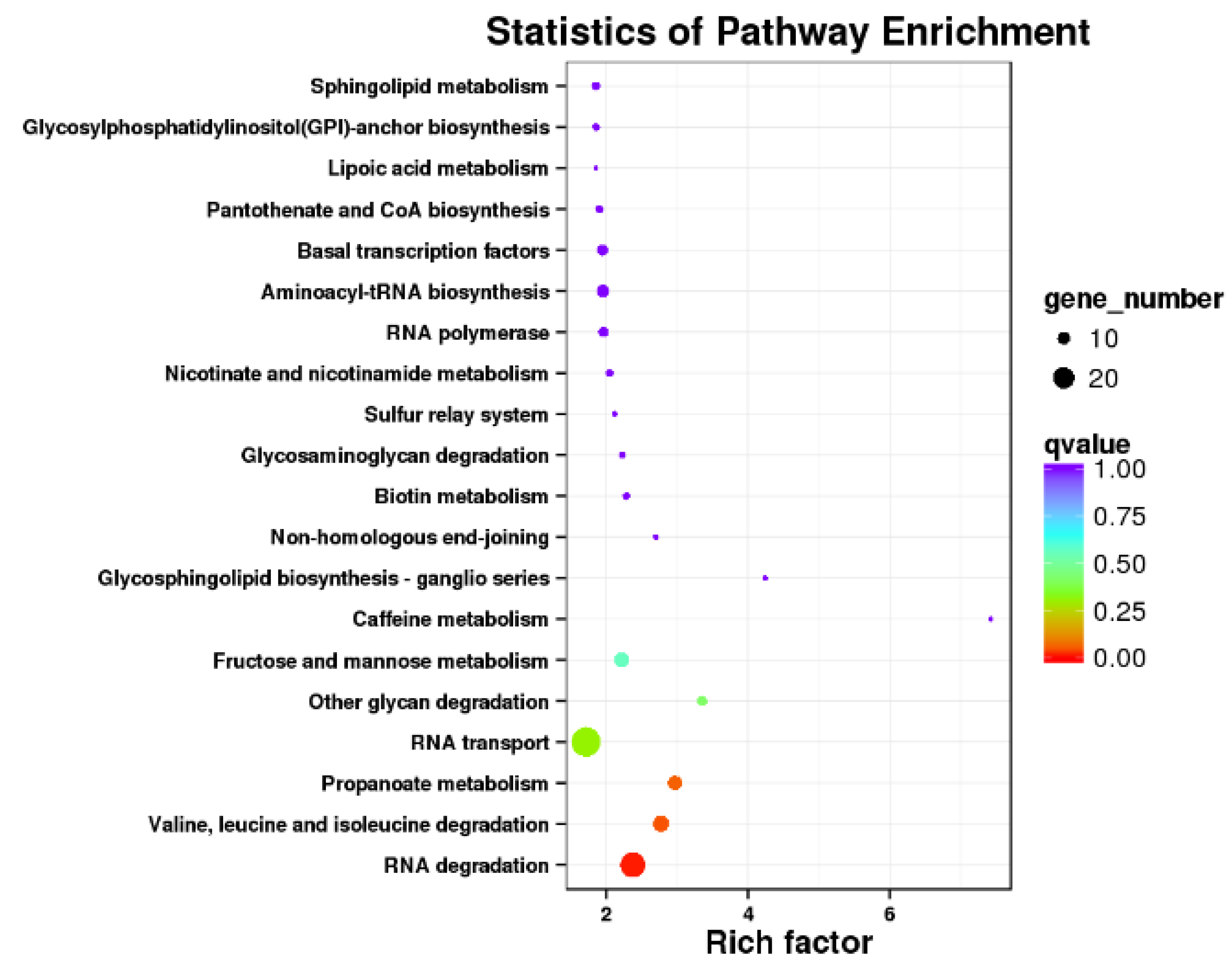
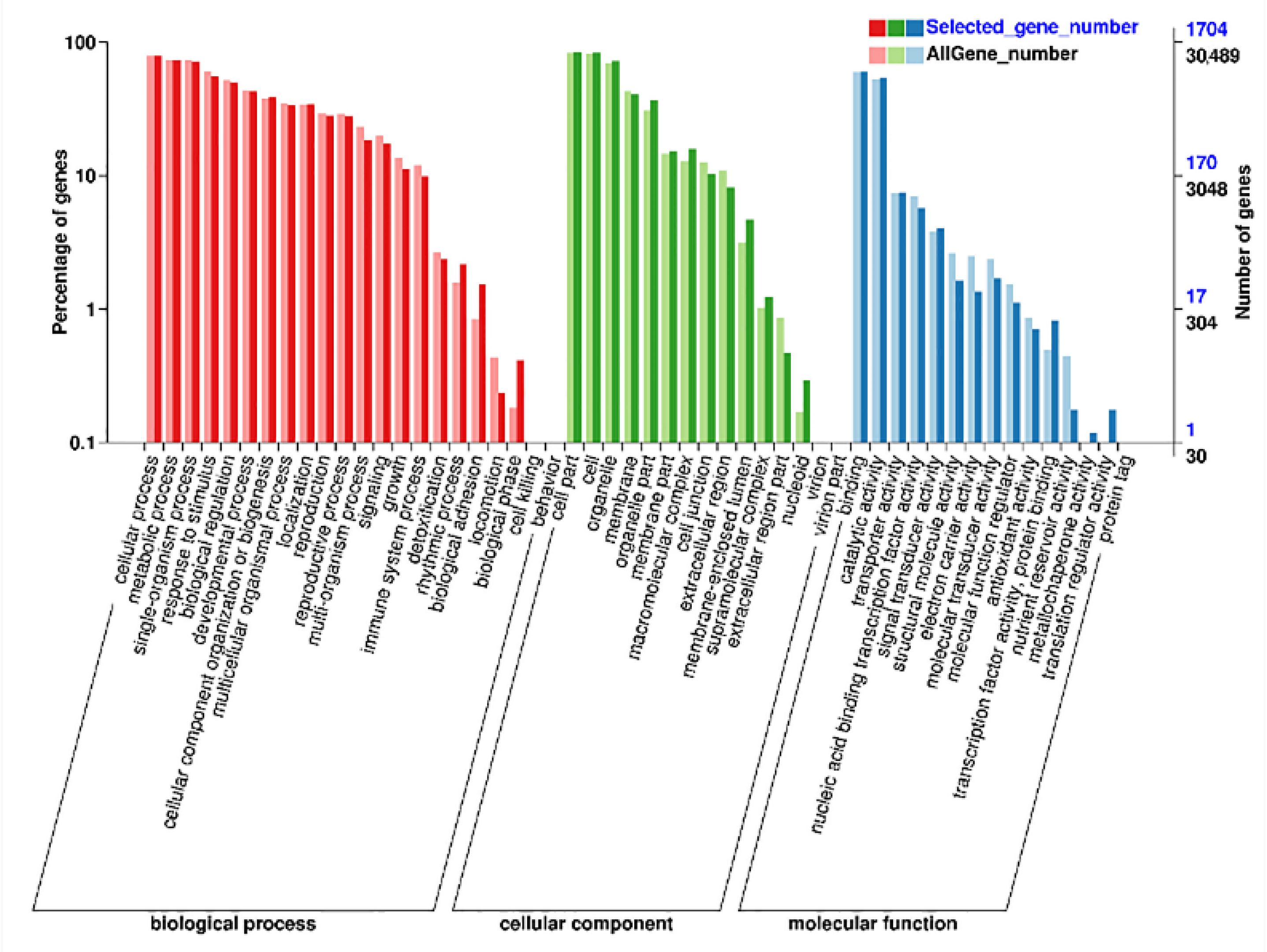
2.5. Functional Annotation and Enrichment Pathway of AS Genes
3. Results
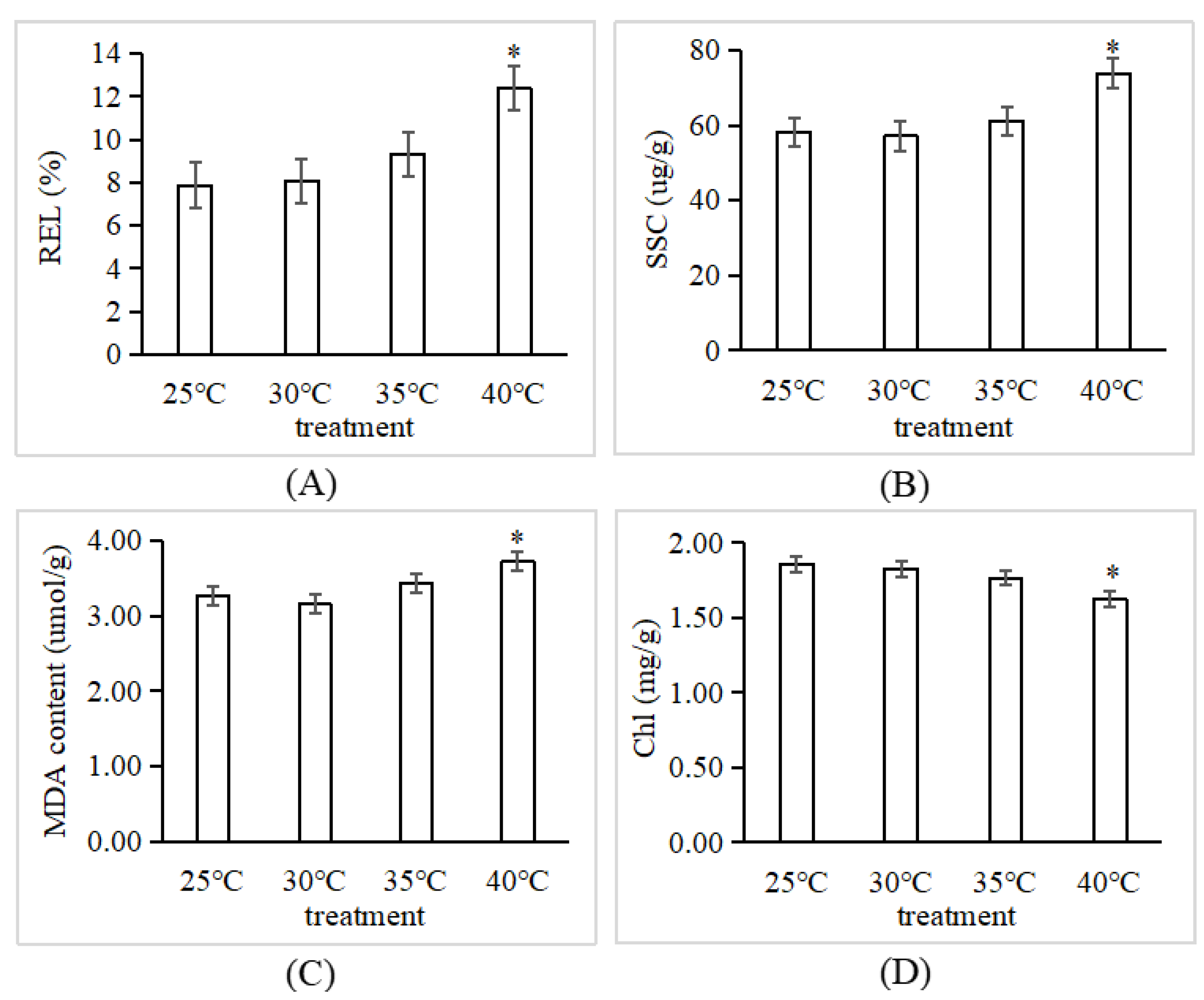
3.1. P. tomentosa Displayed a Strong Resistance to High Temperature
3.2. Overview of RNA-seq Data
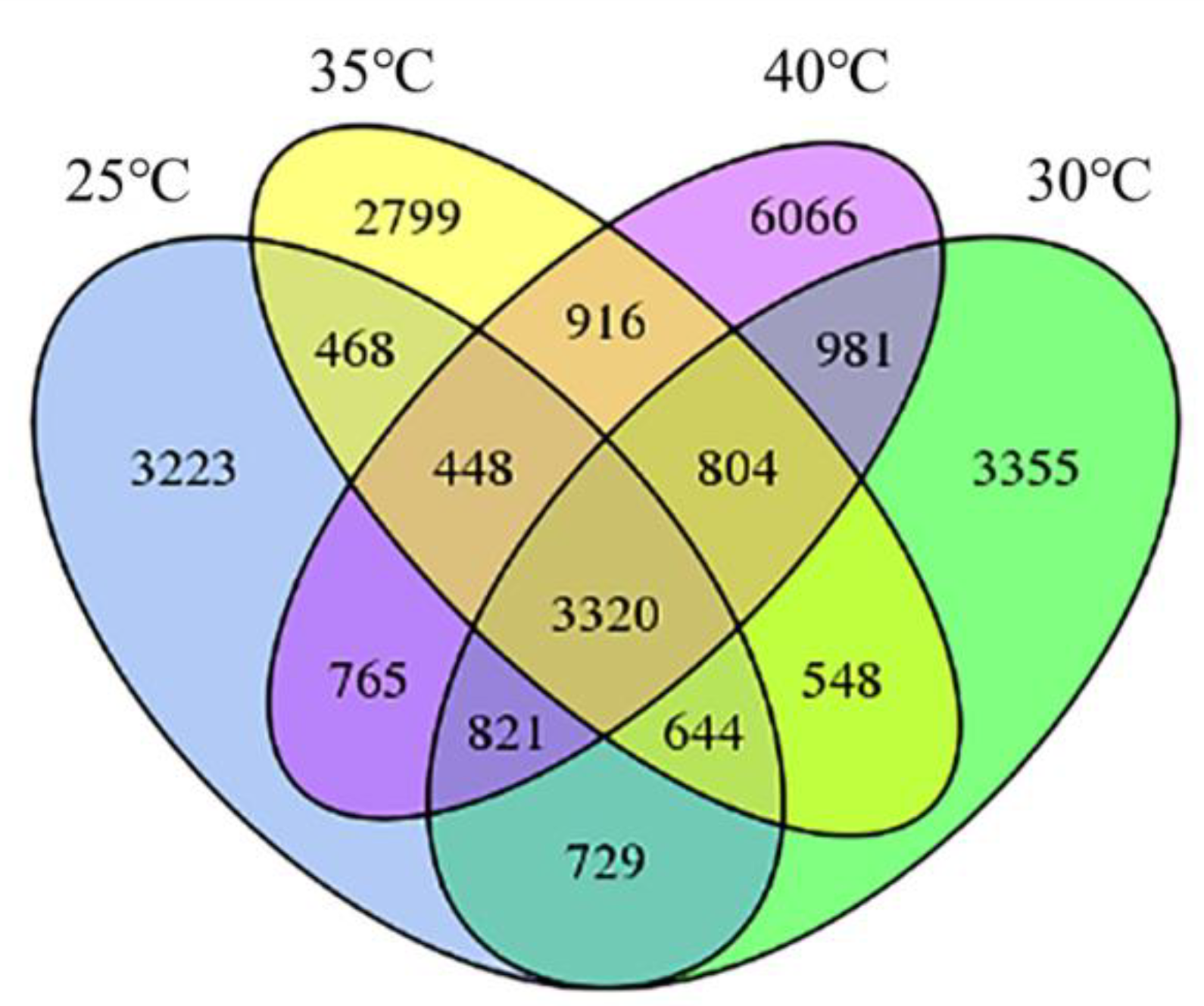
3.3. AS Events in Response to High Temperatures in Poplar
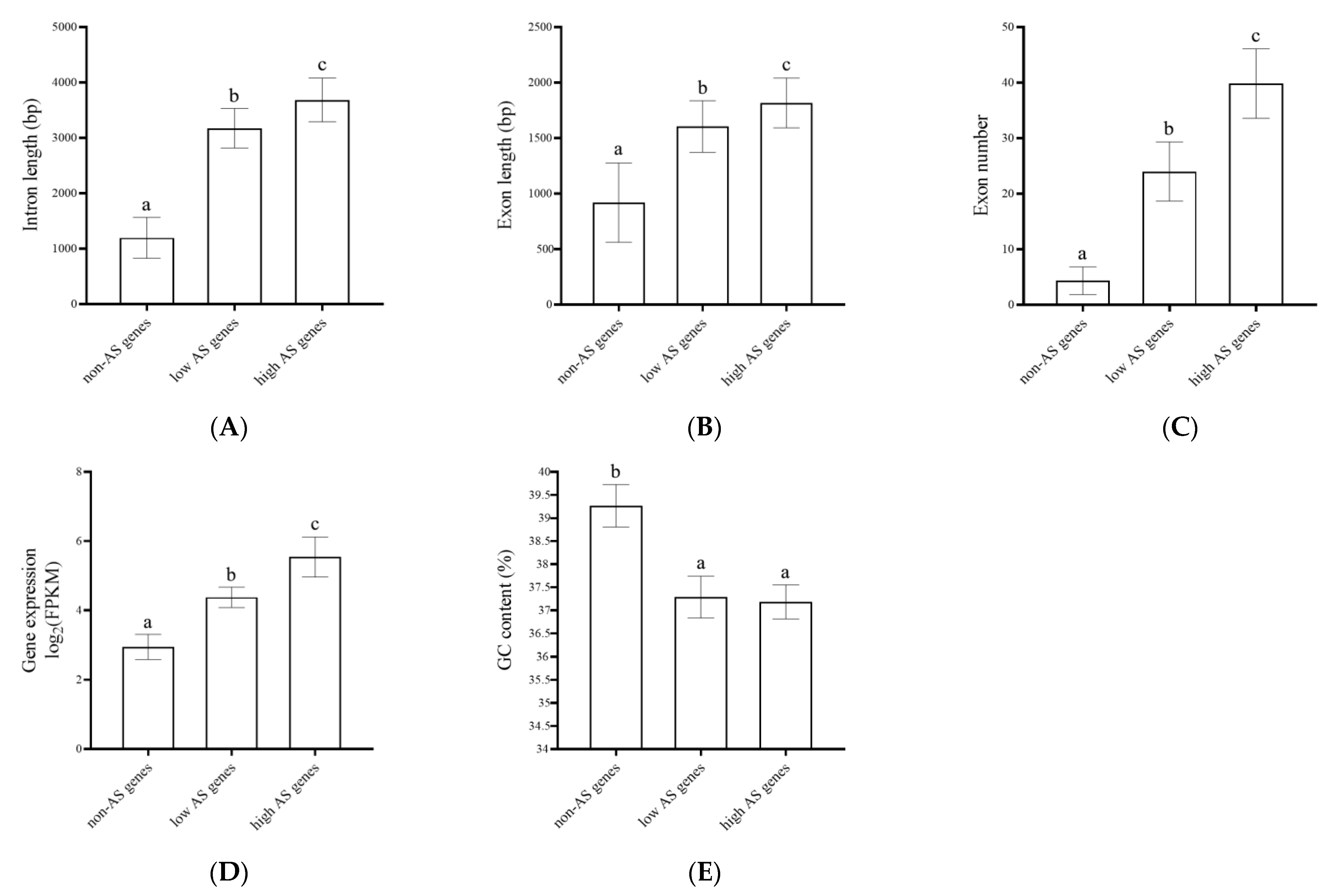
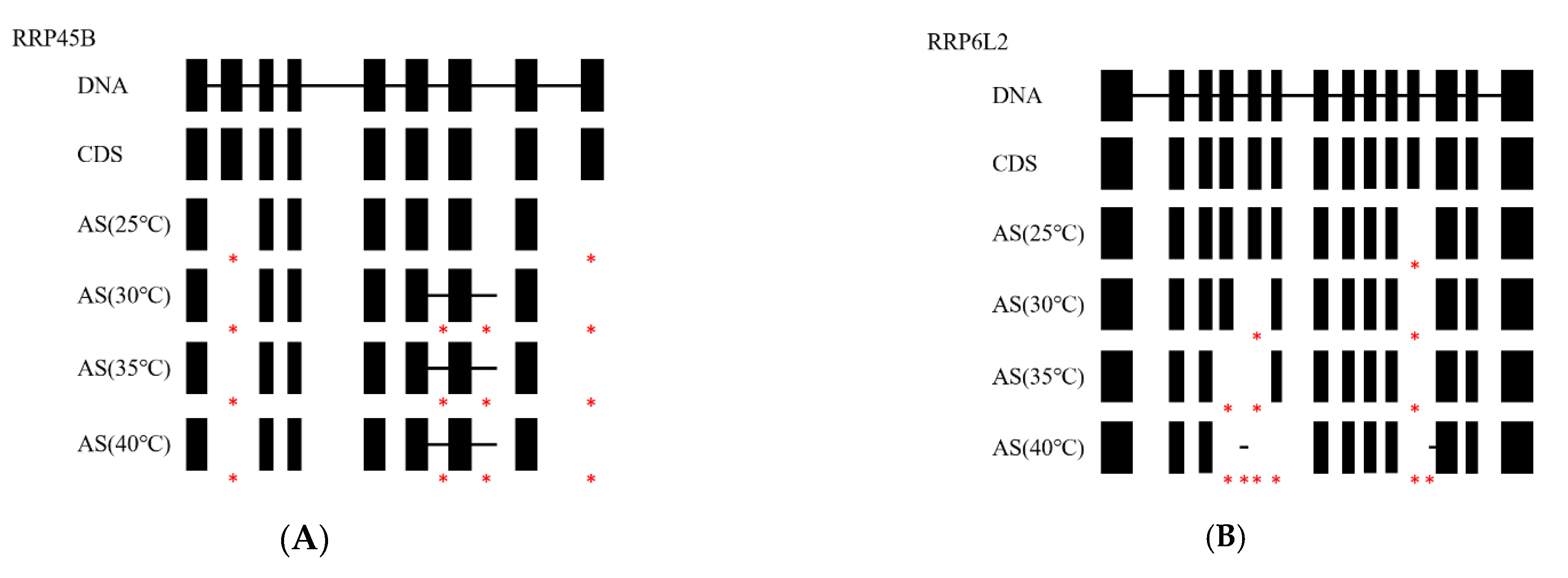
3.4. Structural Analysis of AS Genes in Poplars
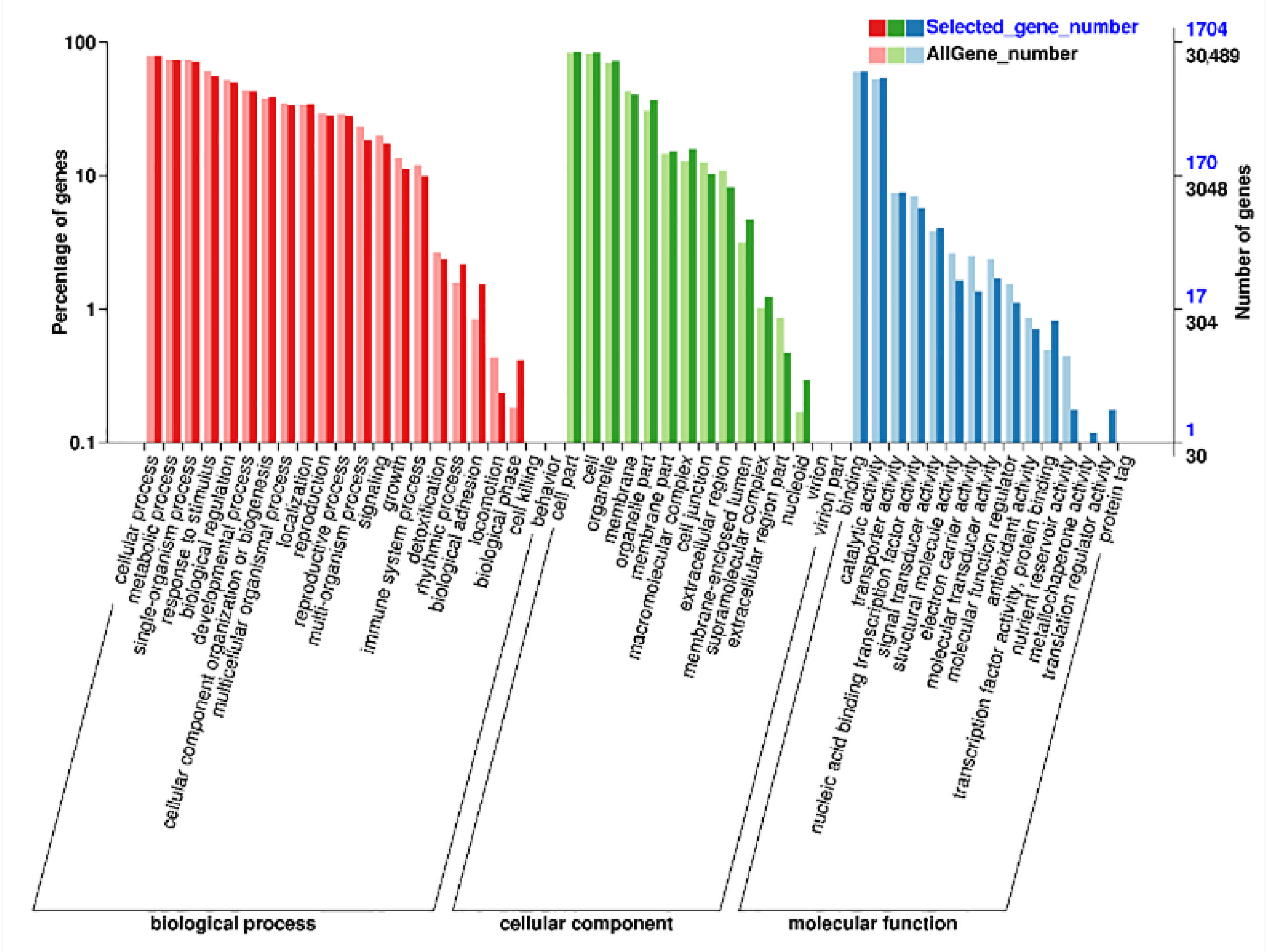
3.5. AS Events Induced by Heat Stress and Function Annotation
4. Discussion
4.1. AS Events Were Specific to the Environmental Conditions
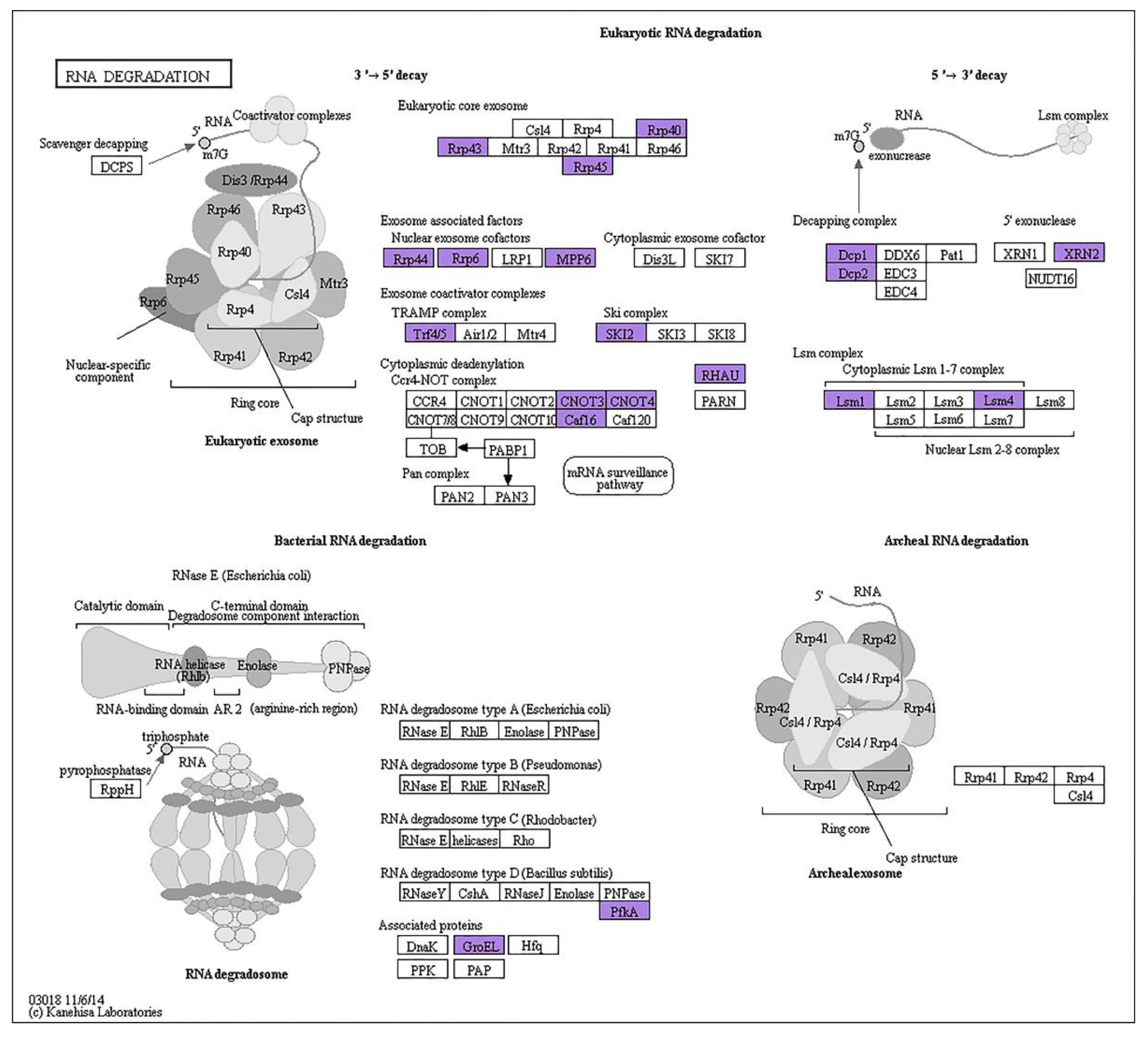
4.2. AS Events in the RNA Degradation Pathway Particularly Affect the Plant Performance in Abiotic Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Godoy, H.; Kornblihtt, A. Alternative splicing and transcription elongation in Plants. Front. Plant Sci. 2019, 10, 309. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.S.N.; Marquez, Y.; Kalyna, M. Complexity of the alternative splicing landscape in plants. Plant Cell 2013, 25, 3657–3683. [Google Scholar] [CrossRef] [PubMed]
- Kannan, S.; Halter, G.; Renner, T.; Waters, E. Patterns of alternative splicing vary between species during heat stress. AoB Plants 2018, 10, 013. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Zhou, Y.; Liu, Z.; Zhang, L.; Song, G.; Guo, Z.; Wang, W.; Qu, X.; Zhu, Y.; Yang, D. An alternatively spliced heat shock transcription factor, Os HSFA 2dI, functions in the heat stress-induced unfolded protein response in rice. Plant Biol. 2015, 17, 419–429. [Google Scholar] [CrossRef]
- Jiang, G.; Zhang, D.; Li, Z.; Liang, H.; Deng, R.; Su, X.; Jiang, Y.; Duan, X. Alternative splicing of MaMYB16L regulates starch degradation in banana fruit during ripening. J. Integr. Plant Biol. 2021, 63, 1341–1352. [Google Scholar] [CrossRef] [PubMed]
- Xiang, L.; Liu, X.; Shi, Y.; Li, Y.; Li, W.; Li, F.; Chen, K. Comparative transcriptome analysis revealed two alternative splicing bHLHs account for flower color alteration in Chrysanthemum. Int. J. Mol. Sci. 2022, 22, 12769. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Guo, Q.; Xu, W.; Liu, P.; Yan, K. Rapid regulation of alternative splicing in response to environmental stresses. Fdront. Plant Sci. 2022, 13, 832177. [Google Scholar] [CrossRef]
- Guo, M.; Zhang, Y.; Jia, X.; Wang, X.; Zhang, Y.; Liu, J.; Yang, Q.; Ruan, W.; Yi, K. Alternative splicing of REGULATOR OF LEAF INCLINATION 1 modulates phosphate starvation signaling and plant growth. Plant Cell 2022, 34, 3319–3338. [Google Scholar] [CrossRef]
- Shen, Y.; Zhou, Z.; Wang, Z.; Li, W.; Fang, C.; Wu, M.; Ma, Y.; Liu, T.; Kong, L.; Peng, D.; et al. Global dissection of alternative splicing in paleopolyploid Soybean. Plant Cell 2014, 26, 996–1008. [Google Scholar] [CrossRef]
- Li, L.; Hu, T.; Li, X.; Mu, S.; Cheng, Z.; Ge, W.; Gao, J. Genome-wide analysis of shoot growth-associated alternative splicing in moso bamboo. Mol. Genet. Genom. 2016, 291, 1695–1714. [Google Scholar] [CrossRef]
- Xu, P.; Kong, Y.; Song, D.; Huang, C.; Li, X.; Li, L. Conservation and functional influence of alternative splicing in wood formation of Populus and Eucalyptus. BMC Genom. 2014, 15, 780. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Yu, K.; Han, L.; Li, X.; Zhang, Y. Global profiling of alternative splicing landscape responsive to salt stress in wheat (Triticum aestivum L.). Plant Growth Regul. 2020, 92, 107–116. [Google Scholar] [CrossRef]
- Li, S.; Yu, X.; Cheng, Z.; Zeng, C.; Li, W.; Zhang, L.; Peng, M. Large-scale analysis of the cassava transcriptome reveals the impact of cold stress on alternative splicing. J. Exp. Bot. 2020, 71, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, S.R.; Danilevskaya, O.N.; Meng, X.; Beatty, M.; Zastrow-Hayes, G.; Harris, C.; Allen, B.V.; Habben, J.; Li, B. Genome-wide analysis of alternative splicing during development and drought stress in maize. Plant Physiol. 2016, 170, 586–599. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; He, F.; Berkowitz, O.; Liu, J.; Cao, P.; Tang, M.; Shi, H.; Wang, W.; Li, Q.; Shen, Z.; et al. Alternative splicing plays a critical role in maintaining mineral nutrient homeostasis in rice (Oryza sativa). Plant Cell 2018, 30, 2267–2285.12. [Google Scholar] [CrossRef] [PubMed]
- Bush, S.; Chen, L.; Tovar-Corona, J.; Urrutia, A. Alternative splicing and the evolution of phenotypic novelty. Philos. Trans. R. Soc. B-Biol. Sci. 2017, 372, 20150474. [Google Scholar] [CrossRef]
- Zhu, J.; Yan, X.; Liu, S.; Xia, X.; An, Y.; Xu, Q.; Zhao, S.; Liu, L.; Guo, R.; Zhang, Z.; et al. Alternative splicing of CsJAZ1 negatively regulates flavan-3-ol biosynthesis in tea plants. Plant J. 2022, 110, 243–261. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circular. 1950, 347, 357–359. [Google Scholar]
- Ding, F.; Cui, P.; Wang, Z.; Zhang, S.; Ali, S.; Xiong, L. Genome-wide analysis of alternative splicing of pre-mRNA under salt stress in Arabidopsis. BMC Genom. 2014, 15, 431. [Google Scholar] [CrossRef]
- Li, Y.; Mi, X.; Zhao, S.; Zhu, J.; Guo, R.; Xia, X.; Liu, L.; Liu, S.; Wei, C. Comprehensive profiling of alternative splicing landscape during cold acclimation in tea plant. BMC Genom. 2020, 21, 65. [Google Scholar] [CrossRef]
- Zhu, G.; Li, W.; Zhang, F.; Guo, W. RNA-seq analysis reveals alternative splicing under salt stress in cotton, Gossypium davidsonii. BMC Genom. 2018, 19, 73. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Qin, J.; Tian, X.; Xu, S.; Wang, Y.; Li, H.; Wang, X.; Peng, H.; Yao, Y.; Hu, Z.; et al. Global profiling of alternative splicing landscape responsive to drought, heat and their combination in wheat (Triticum aestivum L.). Plant Biotechnol. J. 2018, 16, 714–726. [Google Scholar] [CrossRef] [PubMed]
- Braunschweig, U.; Gueroussov, S.; Plocik, A.M.; Graveley, B.R.; Blencowe, B.J. Dynamic integration of splicing within gene regulatory pathways. Cell 2013, 152, 1252–1269. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.; Karin, M. The control of mRNA stability in response to extracellular stimuli. Mol. Cells 2002, 14, 323–331. [Google Scholar]
- Nakaminami, K.; Matsui, A.; Shinozaki, K.; Seki, M. RNA regulation in plant abiotic stress responses. Biochim. Biophys. Acta-Gene Regul. Mech. 2012, 1819, 149–153. [Google Scholar] [CrossRef]
- Yan, X.; Yan, Z.; Han, Y. RRP42, a Subunit of exosome, plays an important role in female gametophytes development and mesophyll cell morphogenesis in Arabidopsis. Front. Plant Sci. 2017, 8, 981. [Google Scholar] [CrossRef]
- Walley, J.W.; Kelley, D.R.; Nestorova, G.; Hirschberg, D.L.; Dehesh, K. Arabidopsis deadenylases AtCAF1a and AtCAF1b play overlapping and distinct roles in mediating environmental stress responses. Plant Physiol. 2010, 152, 866–875. [Google Scholar] [CrossRef]
- Sarowar, S.; Oh, H.W.; Cho, H.S.; Baek, K.H.; Seong, E.S.; Joung, Y.H.; Choi, G.J.; Lee, S.; Choi, D. Capsicum annuum CCR4-associated factor CaCAF1 is necessary for plant development and defence response. Plant J. 2007, 51, 792–802. [Google Scholar] [CrossRef]
- Xu, J.; Yang, J.Y.; Niu, Q.W.; Chua, N.H. Arabidopsis DCP2, DCP1, and VARICOSE form a decapping complex required for postembryonic development. Plant Cell 2006, 18, 3386–3398. [Google Scholar] [CrossRef]
- Roy, K.; Chanfreau, G. Stress-induced nuclear RNA degradation pathways regulate yeast bromodomain factor 2 to promote cell survival. PLoS Genet. 2014, 10, e1004661. [Google Scholar] [CrossRef]
- Hooker, T.S.; Lam, P.; Zheng, H.; Kunst, L. A core subunit of the RNA-processing/degrading exosome specifically influences cuticular wax biosynthesis in Arabidopsis. Plant Cell 2007, 19, 904–913. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Wang, Y.; Qiu, C.; Qian, W.; Xie, H.; Ding, Z. Alternative splicing in tea plants was extensively triggered by drought, heat and their combined stresses. Peer J. 2020, 8, e8258. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | 25 °C | 30 °C | 35 °C | 40 °C |
|---|---|---|---|---|
| Clean reads (bp) | 22,466,189 | 22,478,489 | 22,692,788 | 21,995,836 |
| GC content | 44.94% | 44.96% | 45.22% | 43.78% |
| % ≥ Q30 | 94.98% | 95.04% | 95.36% | 95.20% |
| Mapped reads (bp) | 16,741,731 | 16,669,136 | 16,990,918 | 15,627,313 |
| Unique mapped reads (bp) | 16,071,329 | 15,956,074 | 16,233,291 | 15,132,363 |
| Multiple mapped reads (bp) | 670,403 | 713,063 | 757,628 | 494,950 |
| Treatment Temperature | AS Events | IR | A5SS | A3SS | ES | Sum |
|---|---|---|---|---|---|---|
| 25 °C | AS events | 4659 | 2723 | 2042 | 994 | 10,418 |
| Ratio (%) * | 44.72 | 26.14 | 19.6 | 9.54 | ||
| AS genes | 2599 | 1271 | 965 | 759 | 4105 | |
| 30 °C | AS events | 5713 | 2531 | 1931 | 1027 | 11,202 |
| Ratio (%) * | 51.00 | 22.59 | 17.24 | 9.17 | ||
| AS genes | 2723 | 1323 | 1076 | 792 | 4276 | |
| 35 °C | AS events | 4377 | 2760 | 1901 | 909 | 9947 |
| Ratio (%) * | 44.00 | 27.75 | 19.11 | 9.14 | ||
| AS genes | 2550 | 1231 | 994 | 708 | 4079 | |
| 40 °C | AS events | 5975 | 3735 | 2968 | 1443 | 14,121 |
| Ratio (%) * | 42.31 | 26.45 | 21.02 | 10.22 | ||
| AS genes | 3055 | 1743 | 1469 | 1078 | 4915 |
| Gene | 25 °C | 30 °C | 35 °C | 40 °C |
|---|---|---|---|---|
| RRP40 | 1 | 1 | 1 | 2 |
| CHLH | 1 | 1 | 1 | 1 |
| RRP45B | 2 | 4 | 4 | 4 |
| RRP44A | 2 | 2 | 1 | 3 |
| RRP6L2 | 1 | 2 | 3 | 6 |
| DCP2 | 1 | 1 | 1 | 1 |
| XRN3 | 2 | 3 | 4 | 3 |
| URT1 | 1 | 2 | 1 | 2 |
| SKI2 | 2 | 4 | 3 | 8 |
| HVT1 | 1 | 1 | 1 | 1 |
| VIP2 | 1 | 1 | 2 | 2 |
| ABCI19 | 2 | 2 | 1 | 1 |
| ABCI20 | 2 | 1 | 1 | 3 |
| LSM1B | 1 | 1 | 2 | 2 |
| LSM4 | 1 | 1 | 2 | 2 |
| PFK2 | 1 | 1 | 1 | 1 |
| PFK3 | 1 | 2 | 1 | 2 |
| PFK4 | 2 | 2 | 2 | 4 |
| PFK5 | 1 | 1 | 1 | 2 |
| CPN60B2 | 1 | 1 | 1 | 1 |
| CPN60-2 | 1 | 1 | 1 | 1 |
| Potri.019G032800 | 1 | 1 | 2 | 2 |
| Potri.009G033400 | 1 | 3 | 1 | 3 |
| Potri.006G208600 | 1 | 1 | 1 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Wang, Y.; Wang, R.; Gong, L.; Wang, L.; Xu, J. Genome-Wide Analysis of Alternative Splicing Events Responding to High Temperatures in Populus tomentosa Carr. Forests 2023, 14, 1878. https://doi.org/10.3390/f14091878
Wang X, Wang Y, Wang R, Gong L, Wang L, Xu J. Genome-Wide Analysis of Alternative Splicing Events Responding to High Temperatures in Populus tomentosa Carr. Forests. 2023; 14(9):1878. https://doi.org/10.3390/f14091878
Chicago/Turabian StyleWang, Xue, Yan Wang, Ruixue Wang, Longfeng Gong, Lei Wang, and Jichen Xu. 2023. "Genome-Wide Analysis of Alternative Splicing Events Responding to High Temperatures in Populus tomentosa Carr." Forests 14, no. 9: 1878. https://doi.org/10.3390/f14091878
APA StyleWang, X., Wang, Y., Wang, R., Gong, L., Wang, L., & Xu, J. (2023). Genome-Wide Analysis of Alternative Splicing Events Responding to High Temperatures in Populus tomentosa Carr. Forests, 14(9), 1878. https://doi.org/10.3390/f14091878