
Distribution of Genes and Microbial Taxa Related to Soil Phosphorus Cycling across Soil Depths in Subtropical Forests

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Soil Samples Collection
2.3. Soil Chemical and Physical Analysis
2.4. DNA Extraction and Metagenomic Sequencing
2.5. Metagenomics Analysis
2.6. Statistical Analysis
3. Results
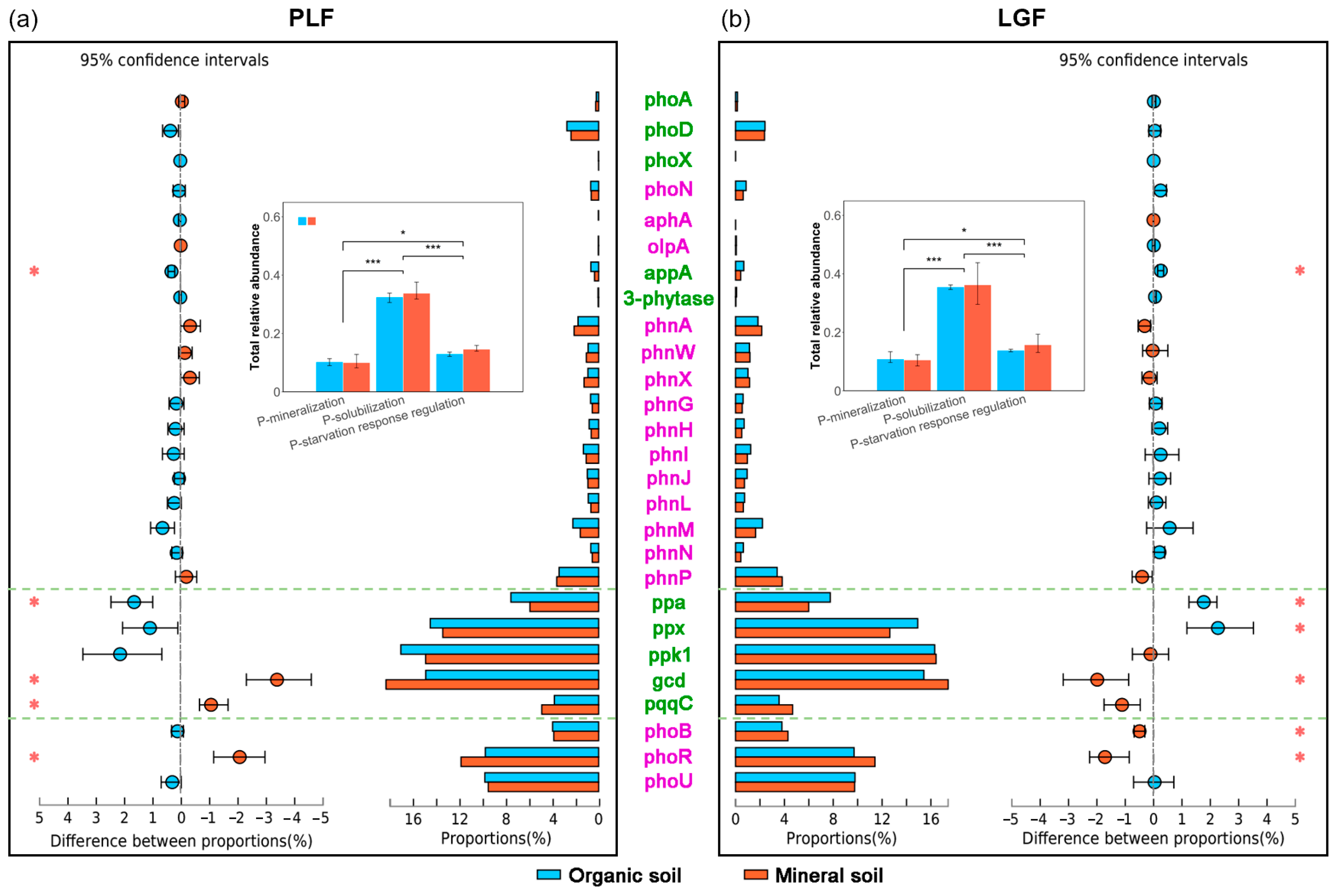
3.1. Differences in Relative Abundance of P Cycling Genes across Soil Depths
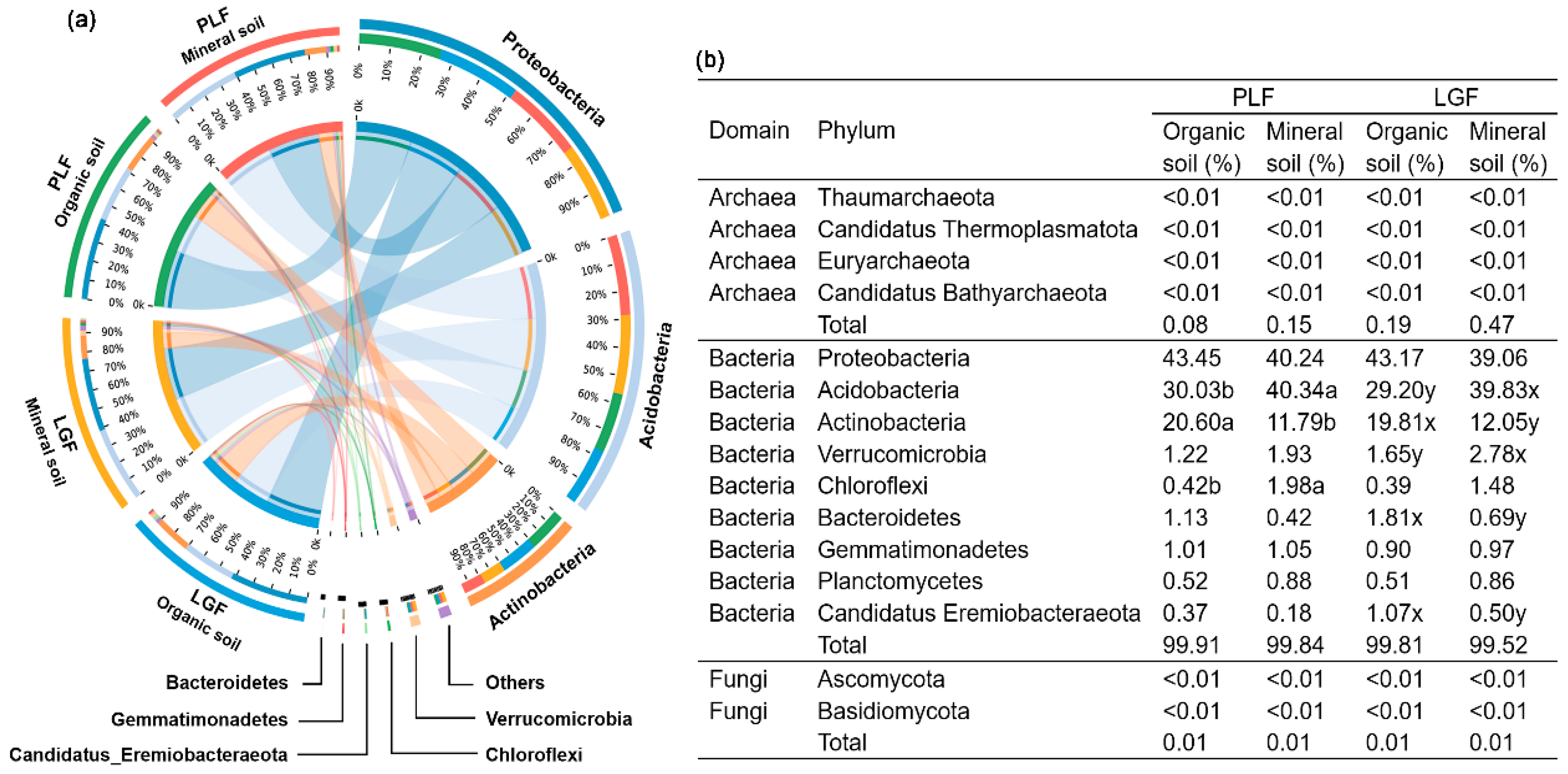
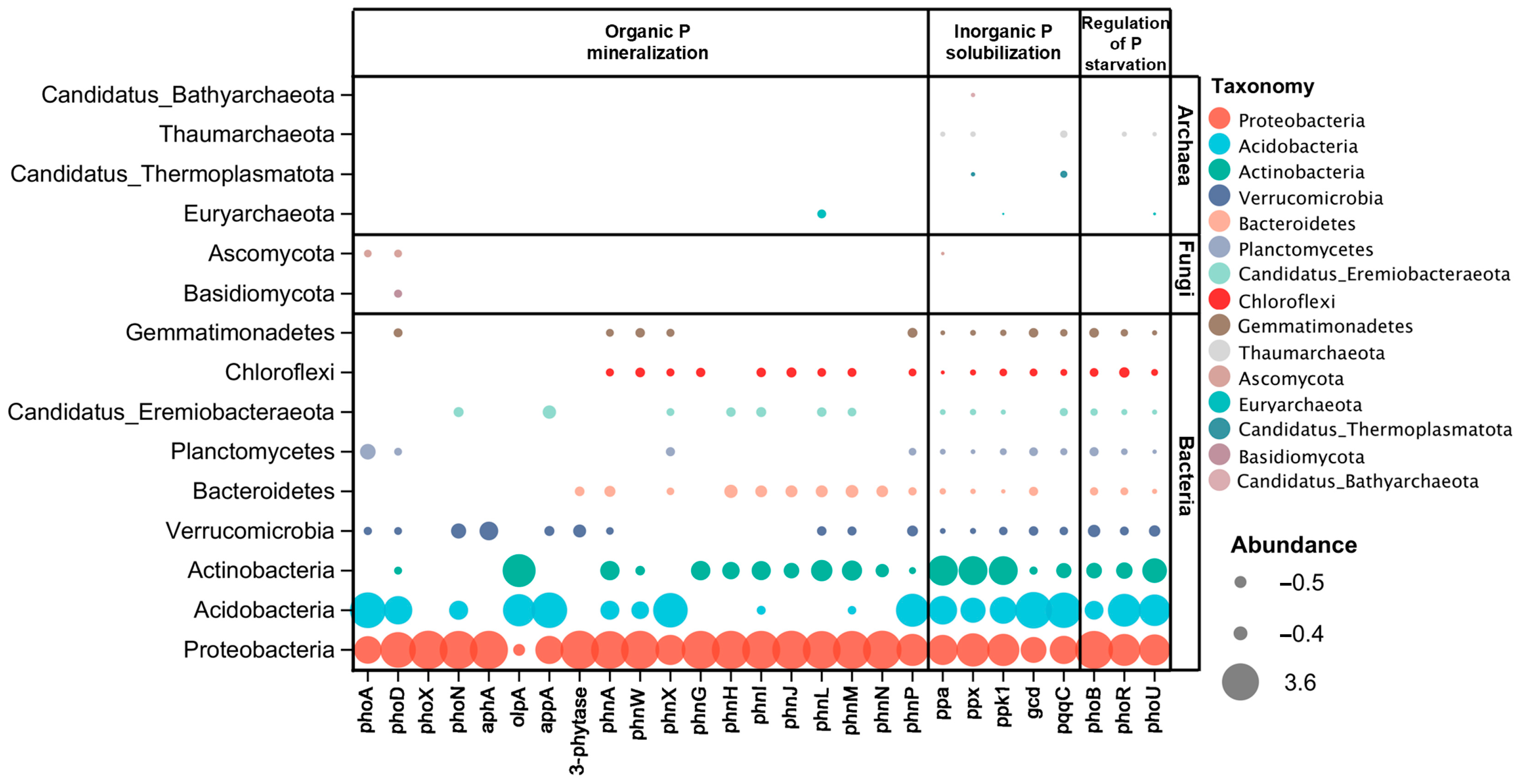
3.2. Differences in Taxonomic Assignments of P Cycling Genes across Soil Depths
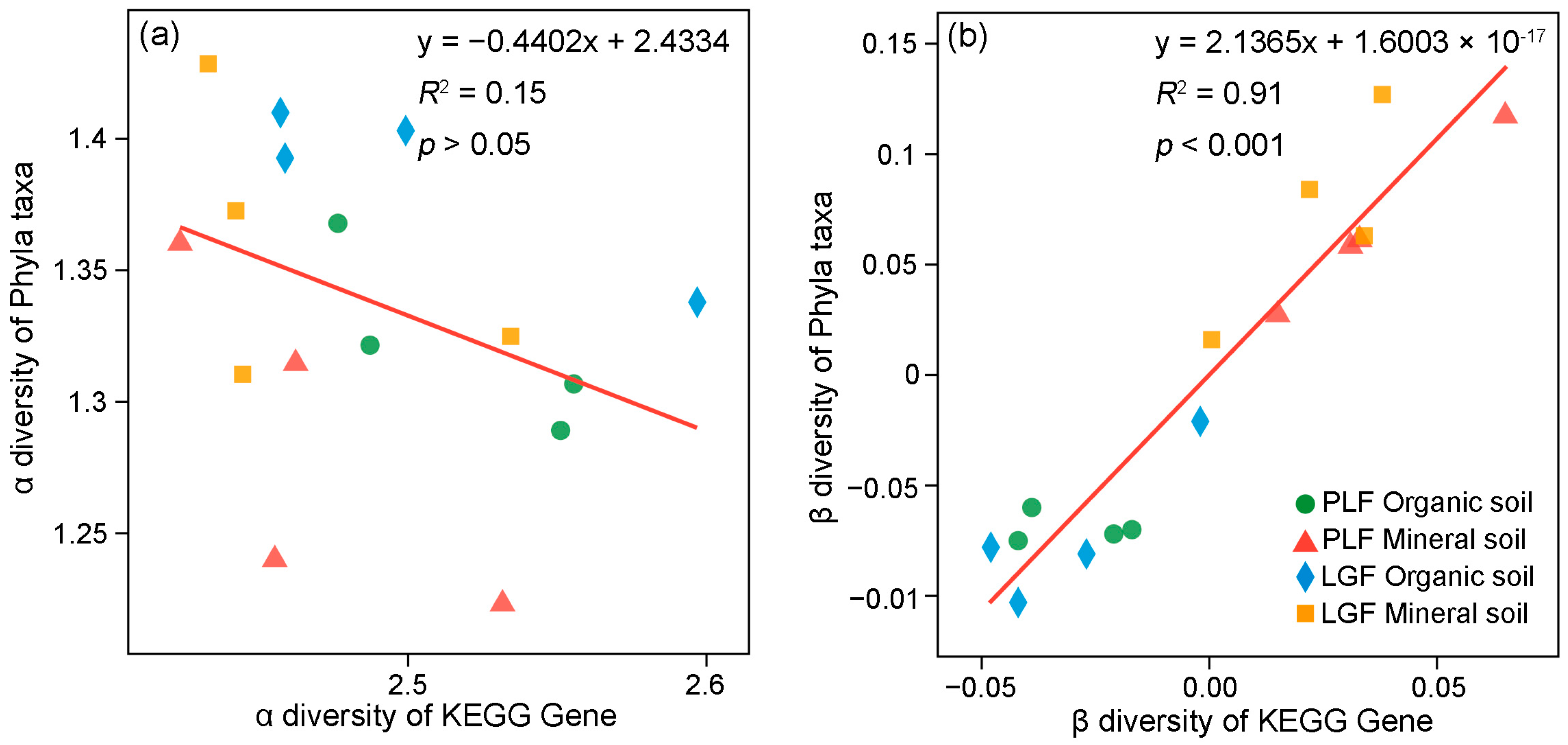
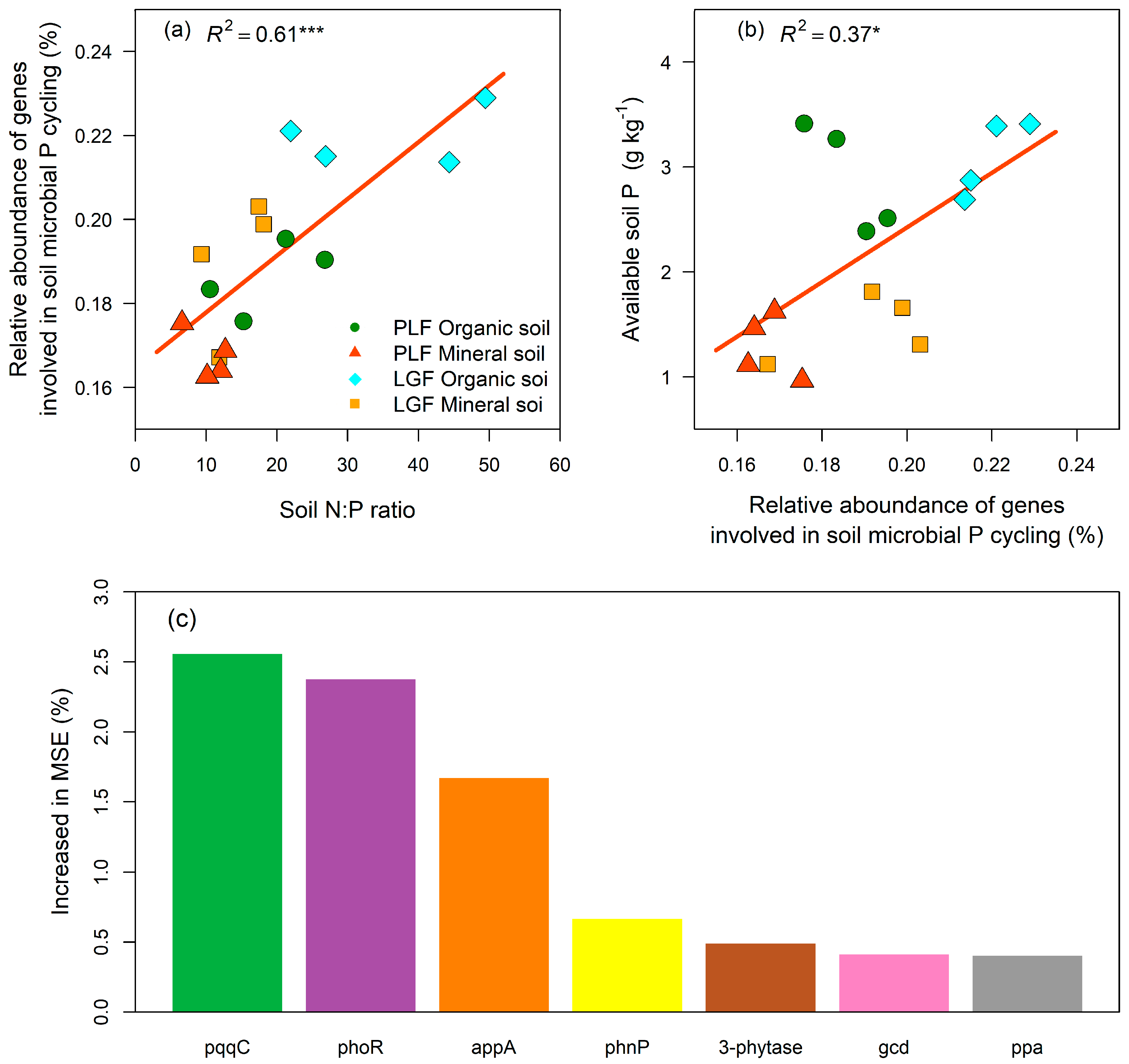
3.3. Linkages between P Cycling Genes and Soil P Status
4. Discussion
4.1. Key Genes Associated with Soil P Cycling in Organic and Mineral Soil
4.2. Phosphorus Cycling Genes Harboring Microbial Taxa Change with Soil Depths
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Du, E.Z.; Terrer, C.; Pellegrini, A.F.A.; Ahlström, A.; van Lissa, C.J.; Zhao, X.; Xia, N.; Wu, X.H.; Jackson, R.B. Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 2020, 13, 221–226. [Google Scholar] [CrossRef]
- Barea, J.M.; Richardson, A.E. Phosphate mobilisation by soil microorganisms. In Principles of Plant-Microbe Interactions; Lugtenberg, B., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 225–234. [Google Scholar]
- Arif, M.S.; Shahzad, S.M.; Yasmeen, T.; Riaz, M.; Ashraf, M.; Ashraf, M.A.; Mubarik, M.S.; Kausar, R. Improving plant phosphorus (P) acquisition by phosphate-solubilizing bacteria. In Essential Plant Nutrients; Naeem, M., Ansari, A.A., Gill, S.S., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 513–556. [Google Scholar]
- Oliverio, A.M.; Bissett, A.; McGuire, K.; Saltonstall, K.; Turner, B.L.; Fierer, N. The role of phosphorus limitation in shaping soil bacterial communities and their metabolic capabilities. mBio 2020, 11, e01718–e01720. [Google Scholar] [CrossRef] [PubMed]
- Hou, E.; Luo, Y.; Kuang, Y.; Chen, C.; Wen, D. Global meta-analysis shows pervasive phosphorus limitation of aboveground plant production in natural terrestrial ecosystems. Nat. Commun. 2020, 11, 637. [Google Scholar] [CrossRef]
- Liang, J.L.; Liu, J.; Jia, P.; Yang, T.T.; Zeng, Q.W.; Zhang, S.C.; Liao, B.; Shu, W.S.; Li, J.T. Novel phosphate-solubilizing bacteria enhance soil phosphorus cycling following ecological restoration of land degraded by mining. ISME J. 2020, 14, 1600–1613. [Google Scholar] [CrossRef] [PubMed]
- Naylor, D.; McClure, R.; Jansson, J. Trends in microbial community composition and function by soil depth. Microorganisms 2022, 10, 540. [Google Scholar] [CrossRef]
- Dai, Z.M.; Liu, G.F.; Chen, H.H.; Chen, C.R.; Wang, J.K.; Ai, S.Y.; Wei, D.; Li, D.M.; Ma, B.; Tang, C.X.; et al. Long-term nutrient inputs shift soil microbial functional profiles of phosphorus cycling in diverse agroecosystems. ISME J. 2020, 14, 757–770. [Google Scholar] [CrossRef]
- Wu, X.J.; Cui, Z.L.; Peng, J.J.; Zhang, F.S.; Liesack, W. Genome-resolved metagenomics identifies the particular genetic traits of phosphate-solubilizing bacteria in agricultural soil. ISME Commun. 2022, 2, 17. [Google Scholar] [CrossRef]
- Yu, H.; He, Z.L.; Wang, A.J.; Xie, J.P.; Wu, L.Y.; Nostrand, J.D.V.; Jin, D.C.; Shao, Z.M.; Schadt, C.W.; Zhou, J.Z.; et al. Divergent responses of forest soil microbial communities under elevated CO2 in different depths of upper soil layers. Appl. Environ. Microbiol. 2017, 84, e01694-17. [Google Scholar]
- Griffiths, R.I.; Whiteley, A.S.; O’Donnell, A.G.; Bailey, M.J. Influence of depth and sampling time on bacterial community structure in an upland grassland soil. FEMS Microbiol. Ecol. 2003, 43, 35–43. [Google Scholar] [CrossRef]
- Fan, Y.; Lin, F.; Yang, L.; Zhong, X.; Wang, M.; Zhou, J.; Chen, Y.; Yang, Y. Decreased soil organic P fraction associated with ectomycorrhizal fungal activity to meet increased P demand under N application in a subtropical forest ecosystem. Biol. Fertil. Soils 2018, 54, 149–161. [Google Scholar] [CrossRef]
- Zhu, X.Y.; Fang, X.; Wang, L.F.; Xiang, W.H.; Alharbi, H.A.; Lei, P.F.; Kuzyakov, Y. Regulation of soil phosphorus availability and composition during forest succession in subtropics. For. Ecol. Manag. 2021, 502, 119706. [Google Scholar] [CrossRef]
- Siles, J.A.; Starke, R.; Martinovic, T.; Fernandes, P.M.L.; Orgiazzi, A.; Bastida, F. Distribution of phosphorus cycling genes across land uses and microbial taxonomic groups based on metagenome and genome mining. Soil Biol. Biochem. 2022, 174, 108826. [Google Scholar] [CrossRef]
- Kishore, N.; Pindi, P.K.; Reddy, S.R. Phosphate-Solubilizing Microorganisms: A Critical Review. In Plant Biology and Biotechnology: Volume I: Plant Diversity, Organization, Function and Improvement; Bahadur, B., Sahijram, M.V.R.L., Krishnamurthy, K.V., Eds.; Springer: New Delhi, India, 2015; pp. 307–347. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014 International soil classification system for naming soils and creating legends for soil maps. In World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Zeng, W.X.; Xiang, W.H.; Zhou, B.; Ouyang, S.; Zeng, Y.L.; Chen, L.; Freschet, G.T.; Valverde-Barrantes, O.J.; Milcu, A. Positive tree diversity effect on fine root biomass: Via density dependence rather than spatial root partitioning. Oikos 2021, 130, 1–14. [Google Scholar] [CrossRef]
- Wu, H.L.; Xiang, W.H.; Ouyang, S.; Forrester, D.I.; Zhou, B.; Chen, L.X.; Ge, T.D.; Lei, P.F.; Chen, L.; Zeng, Y.L.; et al. Linkage between tree species richness and soil microbial diversity improves phosphorus bioavailability. Funct. Ecol. 2019, 33, 1549–1560. [Google Scholar] [CrossRef]
- Ouyang, S.; Xiang, W.H.; Wang, X.P.; Zeng, Y.L.; Lei, P.F.; Deng, X.W.; Peng, C.H. Significant effects of biodiversity on forest biomass during the succession of subtropical forest in south China. For. Ecol. Manag. 2016, 372, 291–302. [Google Scholar] [CrossRef]
- Institute of Soil Science, Chinese Academy of Sciences. Analytical Methods of Soil Physics and Chemistry; Shanghai Scientific and Technical Publishers: Shanghai, China, 1978. [Google Scholar]
- Mehlich, A. Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Li, D.H.; Liu, C.M.; Luo, R.; Sadakane, K.; Lam, T.W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef]
- Noguchi, H.; Park, J.; Takagi, T. MetaGene: Prokaryotic gene finding from environmental genome shotgun sequences. Nucleic Acids Res. 2006, 34, 5623–5630. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H.H.; et al. Vegan: Community Ecology Package, R Package Version 2.0-10; RStudio: Boston, MA, USA, 2013. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Hu, Y.L.; Chen, J.; Hui, D.F.; Wang, Y.P.; Li, J.L.; Chen, J.W.; Chen, G.Y.; Zhu, Y.R.; Zhang, L.Y.; Zhang, D.Q.; et al. Mycorrhizal fungi alleviate acidification-induced phosphorus limitation: Evidence from a decade-long field experiment of simulated acid deposition in a tropical forest in south China. Glob. Chang. Biol. 2022, 28, 3605–3619. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Ma, Y.L.; Wei, R.F.; Chen, Y.L.; Weng, L.P.; Ouyang, X.X.; Li, Y.T. Phosphorus transport in different soil types and the contribution of control factors to phosphorus retardation. Chemosphere 2021, 276, 130012. [Google Scholar] [CrossRef]
- Wu, H.L.; Chen, L.; Ouyang, S.; Zhou, W.N.; Wu, M.G.; Zeng, L.X.; Lei, P.F.; Zeng, Y.L.; Deng, X.W.; Li, S.G.; et al. Phosphorus cycling and supply–demand balance across a chronosequence of Chinese fir plantations. Catena 2023, 228, 107117. [Google Scholar] [CrossRef]
- Wan, W.J.; Hao, X.L.; Xing, Y.H.; Liu, S.; Zhang, X.Y.; Li, X.; Chen, W.L.; Huang, Q.Y. Spatial differences in soil microbial diversity caused by pH-driven organic phosphorus mineralization. Land Degrad. Dev. 2021, 32, 766–776. [Google Scholar] [CrossRef]
- Meyer, J.B.; Frapolli, M.; Keel, C.; Maurhofer, M. Pyrroloquinoline quinone biosynthesis gene pqqC, a novel molecular marker for studying the phylogeny and diversity of phosphate-solubilizing pseudomonads. Appl. Environ. Microbiol. 2011, 77, 7345–7354. [Google Scholar] [CrossRef]
- An, R.; Moe, L.A. Regulation of pyrroloquinoline quinone-dependent glucose dehydrogenase activity in the model rhizosphere-dwelling bacterium Pseudomonas putida KT2440. Appl. Environ. Microbiol. 2016, 82, 4955–4964. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef]
- Chen, X.L.; Chen, H.Y.H.; Chang, S.X. Meta-analysis shows that plant mixtures increase soil phosphorus availability and plant productivity in diverse ecosystems. Nat. Ecol. Evol. 2022, 6, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Augusto, L.; Goll, D.S.; Ringeval, B.; Wang, Y.; Helfenstein, J.; Huang, Y.; Yu, K.; Wang, Z.; Yang, Y. Global patterns and drivers of soil total phosphorus concentration. Earth Syst. Sci. Data 2021, 13, 5831–5846. [Google Scholar] [CrossRef]
- Hsieh, Y.J.; Wanner, B.L. Global regulation by the seven-component Pi signaling system. Curr. Opin. Microbiol. 2010, 13, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Santos-Beneit, F. The Pho regulon: A huge regulatory network in bacteria. Front. Microbiol. 2015, 6, 402. [Google Scholar] [CrossRef]
- Tanuwidjaja, I.; Vogel, C.; Pronk, G.J.; Schöler, A.; Kublik, S.; Vestergaard, G.; Kogel-Knabner, I.; Mrkonjic Fuka, M.; Schloter, M.; Schulz, S. Microbial key players involved in P turnover differ in artificial soil mixtures depending on clay mineral composition. Microb. Ecol. 2021, 81, 897–907. [Google Scholar] [CrossRef]
- Brown, M.R.W.; Arthur, K. Inorganic polyphosphate in the origin and survival of species. Proc. Natl. Acad. Sci. USA 2004, 101, 16085–16087. [Google Scholar] [CrossRef]
- Ghonsikar, C.P.; Miller, R.H. Soil inorganic polyphosphates of microbial origin. Plant Soil 1973, 38, 651–655. [Google Scholar] [CrossRef]
- Kavvadias, V.; Doula, M.; Papadopoulou, M.; Theocharopoulos, S. Long-term application of olive-mill wastewater affects soil chemical and microbial properties. Soil Res. 2015, 53, 461–473. [Google Scholar] [CrossRef]
- Naghshbandi, M.P.; Moghimi, H. Chapter Eighteen—Stabilization of phytase on multi-walled carbon nanotubes via covalent immobilization. In Methods in Enzymology; Kumar, C.V., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 431–451. [Google Scholar]
- Turner, B.L.; Mahieu, N.; Condron, L.M. The phosphorus composition of temperate pasture soils determined by NaOH-EDTA extraction and solution 31P NMR spectroscopy. Org. Geochem. 2003, 34, 1199–1210. [Google Scholar] [CrossRef]
- Yao, Q.; Li, Z.; Song, Y.; Wright, S.J.; Guo, X.; Tringe, S.G.; Tfaily, M.M.; Paša-Tolić, L.; Hazen, T.C.; Turner, B.L.; et al. Community proteogenomics reveals the systemic impact of phosphorus availability on microbial functions in tropical soil. Nat. Ecol. Evol. 2018, 2, 499–509. [Google Scholar] [CrossRef]
- Qiao, Y.; Wang, J.; Liu, H.M.; Huang, K.; Yang, Q.S.; Lu, R.L.; Yan, L.M.; Wang, X.H.; Xia, J.Y. Depth-dependent soil C-N-P stoichiometry in a mature subtropical broadleaf forest. Geoderma 2020, 370, 114357. [Google Scholar] [CrossRef]
- Zheng, B.; Zhu, Y.; Sardans, J.; Peñuelas, J.; Su, J. QMEC: A tool for high throughput quantitative assessment of microbial functional potential in C, N, P, and S biogeochemical cycling. Sci. China Life Sci. 2018, 61, 1451–1462. [Google Scholar] [CrossRef] [PubMed]
- Ragot, S.A.; Kertesz, M.A.; Mészáros, É.; Frossard, E.; Bünemann, E.K. Soil phoD and phoX alkaline phosphatase gene diversity responds to multiple environmental factors. FEMS Microbiol. Ecol. 2017, 93, 212. [Google Scholar] [CrossRef] [PubMed]
- Paula, F.S.; Chin, J.P.; Schnürer, A.; Müller, B.; Manesiotis, P.; Waters, N.; Macintosh, K.A.; Quinn, J.P.; Connolly, J.; Abram, F.; et al. The potential for polyphosphate metabolism in Archaea and anaerobic polyphosphate formation in Methanosarcina mazei. Sci. Rep. 2019, 9, 17101. [Google Scholar] [CrossRef]
- Mundra, S.; Kjonaas, O.J.; Morgado, L.N.; Krabberod, A.K.; Ransedokken, Y.; Kauserud, H. Soil depth matters: Shift in composition and inter-kingdom co-occurrence patterns of microorganisms in forest soils. FEMS Microbiol. Ecol. 2021, 97, fiab022. [Google Scholar] [CrossRef]
- Byers, A.K.; Garrett, L.G.; Armstrong, C.; Dean, F.; Wakelin, S.A. Soil depth as a driver of microbial and carbon dynamics in a planted forest (Pinus radiata) pumice soil. Soil 2023, 9, 55–70. [Google Scholar] [CrossRef]
- Feng, H.; Guo, J.; Wang, W.; Song, X.; Yu, S. Soil depth determines the composition and diversity of bacterial and archaeal communities in a Poplar plantation. Forests 2019, 10, 550. [Google Scholar] [CrossRef]
- Frey, B.; Walthert, L.; Perez-Mon, C.; Stierli, B.; Köchli, R.; Dharmarajah, A.; Brunner, I. Deep soil layers of drought-exposed forests harbor poorly known bacterial and fungal communities. Front. Microbiol. 2021, 12, 674160. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Ling, N.; Chen, D.M.; Guo, H.; Wei, J.X.; Bai, Y.F.; Shen, Q.R.; Hu, S.J. Differential responses of soil bacterial communities to long-term N and P inputs in a semi-arid steppe. Geoderma 2017, 292, 25–33. [Google Scholar] [CrossRef]
- Singavarapu, B.; Du, J.Q.; Beugnon, R.; Cesarz, S.; Eisenhauer, N.; Xue, K.; Wang, Y.F.; Bruelheide, H.; Wubeta, T. Functional potential of soil microbial communities and their subcommunities varies with tree mycorrhizal type and tree diversity. Microbiol. Spectr. 2023, 11, e04578-22. [Google Scholar] [CrossRef] [PubMed]
- Valentine, D.L. Adaptations to energy stress dictate the ecology and evolution of the Archaea. Nat. Rev. Microbiol. 2007, 5, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.; Mikutta, R.; Meyer-Stüve, S.; Guggenberger, G.; Schaarschmidt, F.; Lazar, C.S.; Dohrmann, R.; Schippers, A. Microbial community dynamics in soil depth profiles over 120,000 years of ecosystem development. Front. Microbiol. 2017, 8, 874. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PLF | LGF | |||
|---|---|---|---|---|
| Organic Soil | Mineral Soil | Organic Soil | Mineral Soil | |
| pH | 4.54 ± 0.15 | 4.69 ± 0.18 | 4.17 ± 0.05 y | 4.46 ± 0.08 x |
| SOC (g kg−1) | 75.72 ± 18.54 a | 23.44 ± 3.32 b | 73.35 ± 24.05 x | 28.19 ± 1.47 y |
| Total N (g kg−1) | 2.82 ± 0.26 a | 1.44 ± 0.09 b | 2.84 ± 0.59 x | 1.66 ± 0.15 y |
| NH4+-N (mg kg−1) | 22.74 ± 5.51 | 15.09 ± 6.38 | 31.98 ± 10.82 x | 14.29 ± 4.86 y |
| NO3--N (mg kg−1) | 14.32 ± 4.78 a | 2.20 ± 1.78 b | 15.56 ± 4.97 x | 3.31 ± 1.03 y |
| Total P (g kg−1) | 0.18 ± 0.09 | 0.15 ± 0.05 | 0.09 ± 0.04 | 0.11 ± 0.07 |
| Available P (mg kg−1) | 2.90 ± 0.52 a | 1.29 ± 0.30 b | 3.09 ± 0.37 x | 1.48 ± 0.32 y |
| C/N | 26.80 ± 4.31 a | 19.65 ± 3.31 b | 25.81 ± 4.26 x | 16.94 ± 2.50 y |
| C/P | 431.62 ± 176.81 | 157.89 ± 73.24 | 823.20 ± 450.86 x | 266.93 ± 245.90 y |
| N/P | 16.10 ± 7.06 | 8.04 ± 3.95 | 31.90 ± 13.30 x | 15.76 ± 4.28 y |
| Moisture content (%) | 49.19 ± 0.03 a | 30.73 ± 0.02 b | 53.74 ± 0.06 x | 36.20 ± 0.11 y |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, H.; Yang, J.; Su, S.; Liu, Y.; Feng, J.; Sheng, Y.; Wang, T.; Pan, J.; Tang, L.; Chen, L.; et al. Distribution of Genes and Microbial Taxa Related to Soil Phosphorus Cycling across Soil Depths in Subtropical Forests. Forests 2023, 14, 1665. https://doi.org/10.3390/f14081665
Lv H, Yang J, Su S, Liu Y, Feng J, Sheng Y, Wang T, Pan J, Tang L, Chen L, et al. Distribution of Genes and Microbial Taxa Related to Soil Phosphorus Cycling across Soil Depths in Subtropical Forests. Forests. 2023; 14(8):1665. https://doi.org/10.3390/f14081665
Chicago/Turabian StyleLv, Hao, Jie Yang, Siwen Su, Yue Liu, Jie Feng, Yuxiang Sheng, Ting Wang, Jinwen Pan, Li Tang, Liang Chen, and et al. 2023. "Distribution of Genes and Microbial Taxa Related to Soil Phosphorus Cycling across Soil Depths in Subtropical Forests" Forests 14, no. 8: 1665. https://doi.org/10.3390/f14081665
APA StyleLv, H., Yang, J., Su, S., Liu, Y., Feng, J., Sheng, Y., Wang, T., Pan, J., Tang, L., Chen, L., Ouyang, S., & Wang, G. (2023). Distribution of Genes and Microbial Taxa Related to Soil Phosphorus Cycling across Soil Depths in Subtropical Forests. Forests, 14(8), 1665. https://doi.org/10.3390/f14081665