Biochar Application: A Viable and Pyrolysis Temperature Dependent Option for Enhancing Leaf Secondary Metabolites of Cyclocarya paliurus



Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment Design, Pot Cultivation, Investigation and Sampling
2.2. Analysis of the Properties of Sawdust and Biochar
2.3. Analysis of Soil Properties and Microbial Biomass
2.4. Analysis of Leaf Secondary Metabolites
2.5. Statistical Analysis
3. Results
3.1. Poplar Sawdust and Biochar Properties
3.2. Soil Physical and Chemical Properties
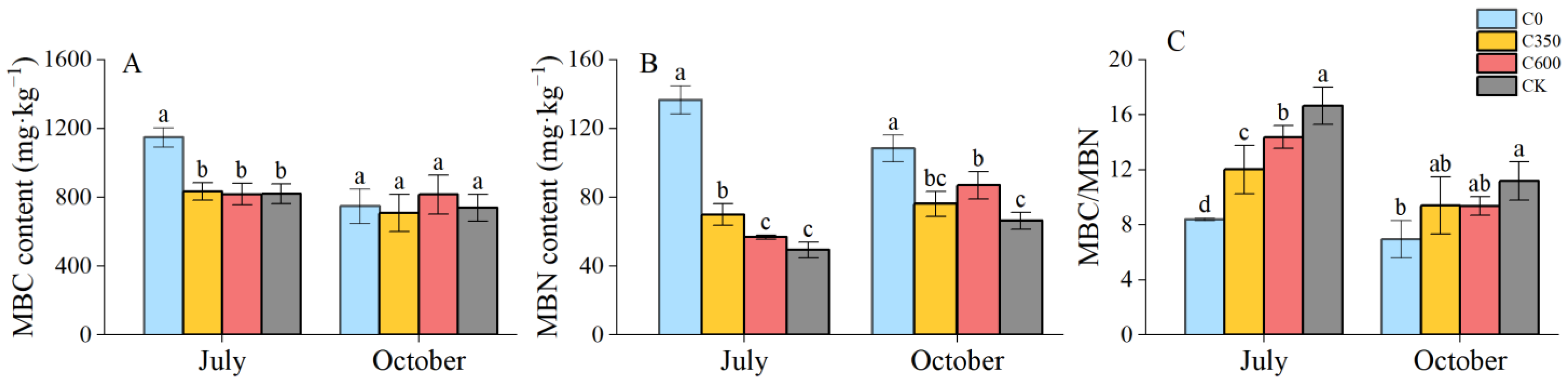
3.3. Soil Microbial Biomass
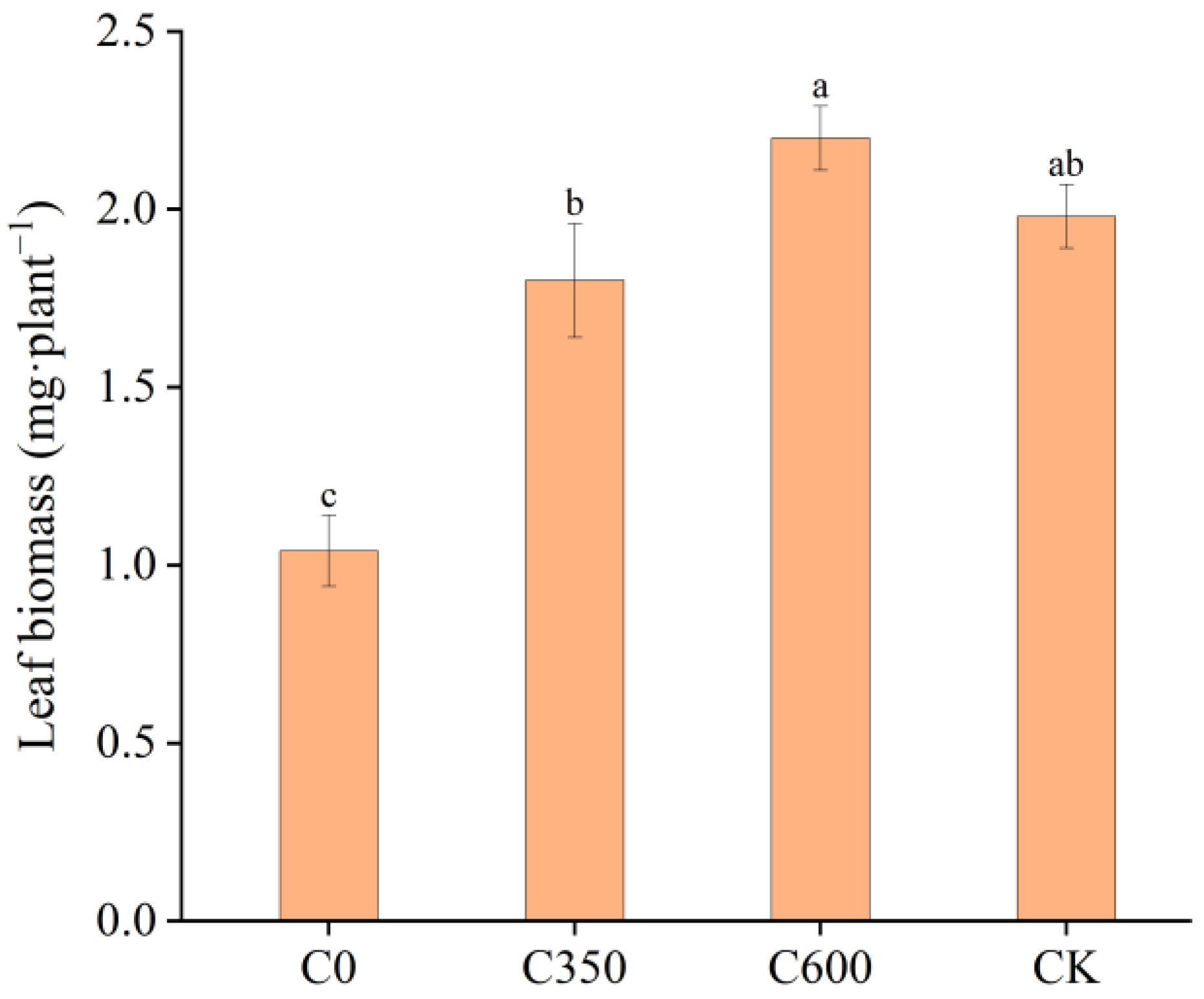
3.4. Seedling Growth and Leaf Biomass of C. paliurus
3.5. Leaf Secondary Metabolite Accumulation
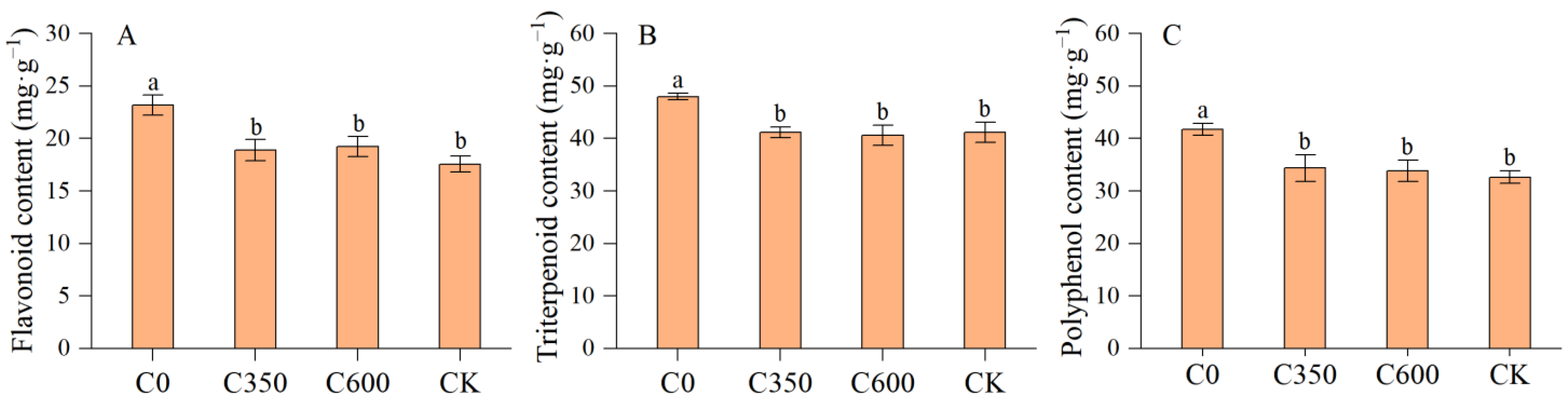
3.5.1. Secondary Metabolite Contents
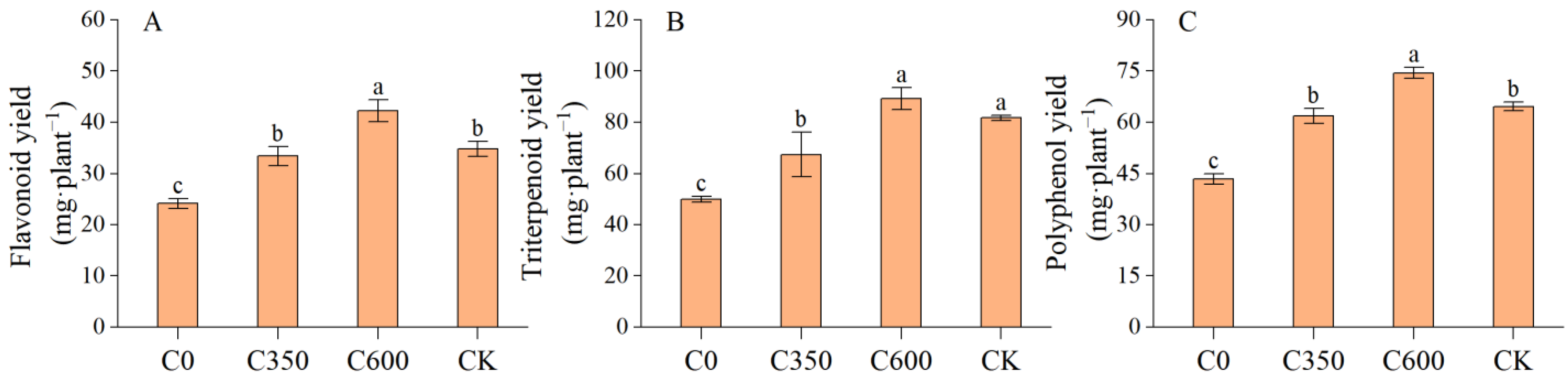
3.5.2. Secondary Metabolite Yield
3.6. Linking Leaf Biomass and Secondary Metabolite Content of Soil Properties
4. Discussion
4.1. Trade-off between Primary Growth and Secondary Metabolism under Biochars Application
4.2. Effect of Direct Application of Poplar Sawdust on Growth and Secondary Metabolism
4.3. Response of Secondary Metabolite Monomers to Biochars and Poplar Sawdust Application
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xie, G.U.; Fu, T.C.; Ma, L.Y.; Li, H.; Bao, W.Q.; Li, S. An overview of definition and classification of forestry residue. J. China Agric. Univ. 2018, 23, 141–149. [Google Scholar] [CrossRef]
- Duan, X.F.; Zhou, Z.F.; Xu, J.M.; Wang, R. Utilization situation and suggestion of forestry residues resources in China. China Wood-Based Panel. 2017, 24, 1–5. [Google Scholar]
- Xu, Y.; Du, X.Z.; Qi, Y.J.; Ma, L.; Zheng, G. Utilization of wood processing residues. China For. Prod. Ind. 2015, 42, 40–44. [Google Scholar] [CrossRef]
- Qiu, H.G.; Yan, J.B.; Lei, Z.; Sun, D. Rising wages and energy consumption transition in rural China. Energy Policy. 2018, 119, 545–553. [Google Scholar] [CrossRef]
- Sohi, S.P.; Krull, E.; Lopez-capel, E.; Bol, R. A review of biochar and its use and function in soil. Adv. Agron. 2010, 105, 47–82. [Google Scholar] [CrossRef]
- Lehmann, J.; Gaunt, J.; Rondon, M. Bio-char sequestration in terrestrial ecosystems—A review. Mitig. Adapt. Strateg. Glob. Change 2006, 11, 403–427. [Google Scholar] [CrossRef]
- Chen, F.W.; Zhang, W.M.; Meng, J. Biochar and Agro-ecological Environment: Review and Prospect. J. Agro-Environ. Sci. 2014, 33, 811–818. [Google Scholar] [CrossRef]
- Liang, B.; Lehmann, J.; Solomon, D.; Kinyangi, J.; Grossman, J.; O’Neill, B.; Skjemstad, J.O.; Thies, J.; Luizão, F.J.; Petersen, J.; et al. Black carbon increases cation exchange capacity in soils. Soil Sci. Soc. Am. J. 2006, 70, 1719–1730. [Google Scholar] [CrossRef]
- Spokas, A.K.; Cantrell, B.K.; Novak, M.J.; Archer, D.W.; Ippolito, J.A.; Collins, H.P.; Boateng, A.A.; Lima, I.M.; Lamb, M.C.; McAloon, A.J.; et al. Biochar: A synthesis of its agronomic impact beyond carbon sequestration. J. Environ. Qual. 2012, 41, 973–989. [Google Scholar] [CrossRef]
- Tomczyk, A.; Sokolowska, Z.; Boguta, P. Biochar physicochemical properties: Pyrolysis temperature and feedstock kind effects. Rev. Environ. Sci. Bio/Technol. 2020, 19, 191–215. [Google Scholar] [CrossRef]
- Yuan, S.; Zhao, L.X.; Meng, H.B.; Shen, Y.J. The main types of biochar and their properties and expectative researches. J. Plant Nutr. Fertil. 2016, 22, 1402–1417. [Google Scholar]
- Ippolito, J.A.; Cui, L.Q.; Kammann, C.; Wrage-Mönnig, N.; Estavillo, J.M.; Fuertes-Mendizabal, T.; Cayuela, M.L.; Sigua, G.; Novak, J.; Spokas, K.; et al. Feedstock choice, pyrolysis temperature and type influence biochar characteristics: A comprehensive meta-data analysis review. Biochar 2020, 2, 421–438. [Google Scholar] [CrossRef]
- Ajayi, A.E.; Horn, R. Modification of chemical and hydrophysical properties of two texturally differentiated soils due to varying magnitudes of added biochar. Soil Tillage Res. 2016, 164, 34–44. [Google Scholar] [CrossRef]
- Tang, H.M. Effects of Microbial Fertilize and Biochar Applications on the Yield and Quality of Pinelia Ternata and Soil Microecology; Huazhong Agricultural University: Wuhan, China, 2020; Volume 2. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Chen, J.J.; Younis, A. Biochar, compost, and biochar-compost blend applications modulate growth, photosynthesis, osmolytes, and antioxidant system of medicinal plant Alpinia zerumbet. Front. Plant Sci. 2021, 12, 707061. [Google Scholar] [CrossRef]
- Saha, A.; Basak, B.B.; Gajbhiye, N.A.; Kalariya, K.A.; Manivel, P. Sustainable fertilization through co-application of biochar and chemical fertilizers improves yield, quality of Andrographis paniculata and soil health. Ind. Crops Prod. 2019, 140, 111607. [Google Scholar] [CrossRef]
- Coruzzi, G.; Bush, D.R. Nitrogen and Carbon Nutrient and Metabolite Signaling in Plants. Plant Physiol. 2001, 125, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Stamp, N. Out of the quagmire of plant defense hypotheses. Q. Rev. Biol. 2003, 78, 23–55. [Google Scholar] [CrossRef] [PubMed]
- Kuang, K.Q.; Li, P.Q. Flora of China; Science Press: Beijing, China, 1979; pp. 18–19. [Google Scholar]
- Chen, Y.; Chen, W.; Li, F.T.; Ma, L. Research progress on chemical constituents and pharmacological actions of Cyclocarya paliurus. Anim. Husb. Feed. Sci. 2019, 40, 12–13. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, S.L.; Feng, Z.W.; Wang, H.; Huang, H. Comparative study of selected soil properties following introduction of broad-leaf trees into clear-felled Chinese fir forest. Commun. Soil Sci. Plant Anal. 2005, 36, 1385–1403. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Feng, Z.S.; Wu, C.E.; Fang, S.Z.; Yang, W.X.; Yang, J.T.; Li, T.T. Technology optimization of total flavonoids extraction from Cyclocarya paliurus leaves by ultrasonic assistance. Trans. Chin. Soc. Agric. Mach. 2009, 49, 130–134. [Google Scholar]
- Yue, X.L.; Qin, J.; Fu, X.X.; Shang, X.L.; Fang, S.Z. Effects of nitrogen fertilization on secondary metabolite accumulation and antioxidant capacity of Cycolcurya paliurus (Batal.) Iljinskaja Leaves. J. Nanjing For. Univ. Nat. Sci. Ed. 2020, 44, 35–42. [Google Scholar] [CrossRef]
- Fan, J.P.; He, C.H. Simultaneous quantification of three major bioactive triterpene acids in the leaves of Diospyros kaki by high performance liquid chromatography method. J. Pharm. Biomed. Anal. 2006, 41, 950–956. [Google Scholar] [CrossRef]
- Alothman, M.; Bhat, R.; Karim, A.A. Antioxidant capacity and phenolic content of selected tropical fruits from Malaysia, extracted with different solvent. Food Chem. 2009, 115, 785–788. [Google Scholar] [CrossRef]
- Cao, Y.N.; Fang, S.Z.; Yin, Z.Q.; Fu, X.X.; Shang, X.L.; Yang, W.X.; Yang, H.M. Chemical fingerprint and multicomponent quantitative analysis for the quality evaluation of Cyclocarya paliurus leaves by HPLC–Q–TOF–MS. Molecules 2017, 22, 1927. [Google Scholar] [CrossRef]
- Steiner, C.; Teixeira, W.G.; Lehmann, J.; Nehls, T.; Macêdo, J.L.; Blum, W.; Zech, W. Long term effects of manure, charcoal and mineral fertilization on crop production and fertility on a highly weathered Central Amazonian upland soil. Plant Soil. 2007, 291, 275–290. [Google Scholar] [CrossRef]
- Jeffery, S.; Verheijen, F.G.A.; Velde, M.; Bastos, A.C. A quantitative review of the effects of biochar application to soils on crop productivity using meta-analysis. Agric. Ecosyst. Environ. 2011, 144, 175–187. [Google Scholar] [CrossRef]
- Major, J.; Rondon, M.; Molina, D.; Riha, S.J.; Lehmann, J. Maize yield and nutrition during 4 years after biochar application to a Colombian savanna oxisol. Plant Soil. 2010, 333, 117–128. [Google Scholar] [CrossRef]
- Zwieten, L.V.; Kimber, S.; Morris, S.; Chan, K.Y.; Downie, A.; Rust, J.; Joseph, S.; Cowie, A. Effects of biochar from slow pyrolysis of papermill waste on agronomic performance and soil fertility. Plant Soil. 2010, 327, 235–246. [Google Scholar] [CrossRef]
- Güereña, D.; Lehmann, J.; Hanley, K.; Enders, A.; Hyland, C.; Riha, S. Nitrogen dynamics following field application of biochar in a temperate North American maize-based production system. Plant Soil. 2013, 365, 239–254. [Google Scholar] [CrossRef]
- Deng, R.; Lan, Z.Y.; Shang, X.L.; Fang, S.Z. Effects of biochar application pyrolyzed at different temperatures on soil properties, growth and leaf secondary metabolite accumulation in Cyclocarya paliurus. Forsets 2022, 13, 1572. [Google Scholar] [CrossRef]
- Glaser, B.; Lehmann, J.; Zech, W. Ameliorating physical and chemical properties of highly weathered soils in the tropics with charcoal-a review. Biol. Fertil. Soils 2002, 35, 119–130. [Google Scholar] [CrossRef]
- Weber, K.; Quicker, P. Properties of biochar. Fuel 2018, 217, 240–261. [Google Scholar] [CrossRef]
- Abideen, Z.; Koyro, H.W.; Huchzermeyer, B.; Ansari, R.; Zulfiqar, F.; Gulet, B. Ameliorating effects of biochar on photosynthetic efficiency and antioxidant defence of Phragmites karka under drought stress. Plant Biol. 2020, 22, 259–266. [Google Scholar] [CrossRef]
- Frischknecht, P.M.; Schuhmacher, K.; Müller-Schärer, H.; Baumann, T.W. Phenotypic plasticity of Senecio vulgaris from contrasting habitat types: Growth and pyrrolizidine alkaloid formation. J. Chem. Ecol. 2001, 27, 343–358. [Google Scholar] [CrossRef]
- Chen, G.; Qiao, J.; Zhao, G.H.; Zhang, H.M.; Shen, Y.Q.; Cheng, W.D. Rice-straw biochar regulating effect on Chrysanthemum morifolium Ramat. cv. ‘Hangbaiju’. Agron. J. 2018, 110, 1996–2003. [Google Scholar] [CrossRef]
- Petruccelli, R.; Bonetti, A.; Traversi, M.L.; Faraloni, C.; Valagussa, M.; Pozzi, A. Influence of biochar application on nutritional quality of tomato (Lycopersicon esculentum). Crop Pasture Sci. 2015, 66, 747–755. [Google Scholar] [CrossRef]
- Zhao, J.; Ni, T.; Xun, W.B.; Huang, X.L.; Huang, Q.W.; Ran, W.; Shen, B.; Zhang, R.F.; Shen, Q.R. Influence of straw incorporation with and without straw decomposer on soil bacterial community structure and function in a rice-wheat cropping system. Appl. Microbiol. Biotechnol. Vol. 2017, 101, 4761–4773. [Google Scholar] [CrossRef]
- Kaye, J.; Hart, S. Competition for nitrogen between plants and soil microorganisms. Trends Ecol. Evol. 1997, 12, 139–143. [Google Scholar] [CrossRef]
- Li, H.; Dai, M.W.; Dai, S.L.; Dong, X.J. Current status and environment impact of direct straw return in China’s cropland–A review. Ecotoxicol. Environ. Saf. 2018, 159, 293–300. [Google Scholar] [CrossRef]
- Liang, J.P.; Jia, X.Y.; Liu, Y.L.; Wu, Y.; Zhou, R.; Feng, Q.J. Effects of drought stress on seedling growth and accumulation of secondary metabolites in the roots of Astragalus membranaceus var. mongholicus. Acta Ecol. Sinica. 2016, 36, 4415–4422. [Google Scholar] [CrossRef]
- Xu, X.L.; Ouyang, H.; Pei, Z.Y.; ZHOU, C.P. Long-term partitioning of ammonium and nitrate among different components in an alpine meadow ecosystem. Acta Bot. Sinica. 2004, 46, 279–283. [Google Scholar] [CrossRef]
- Jiang, J.; Song, M.H. Review of the roles of plants and soil microorganisms in regulating ecosystem nutrient cycling. Chin. J. Plant Ecol. 2010, 38, 979–988. [Google Scholar] [CrossRef]
- Hodge, A.; Robinson, D.; Fitter, A. Are microorganisms more effective than plants at competing for nitrogen? Trends Plant Sci. 2000, 5, 304–308. [Google Scholar] [CrossRef] [PubMed]
- Bryant, J.P.; Chapin, F.S.; Klein, D.R. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 1983, 40, 357–368. [Google Scholar] [CrossRef]
- Recous, S.; Robin, D.; Darwis, D.; Mary, B. Soil inorganic N availability: Effect on maize residue decomposition. Soil Biol. Biochem. 1995, 27, 1529–1538. [Google Scholar] [CrossRef]
- Liu, C.; Lu, M.; Cui, J.; Li, B.; Fanget, C.M. Effects of straw carbon input on carbon dynamics in agricultural soils: A meta-analysis. Glob. Change Biol. 2014, 20, 1366–1381. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Sun, B.F.; Lu, F.; Zhang, G.; Wang, X.K.; Ouyang, Z.Y. Straw incorporation strategy on cereal crop yield in China. Crop Sci. 2015, 55, 1773–1781. [Google Scholar] [CrossRef]
- Lou, Y.; Xu, M.; Wang, W.; Sun, X.L.; Zhao, K. Return rate of straw residue affects soil organic C sequestration by chemical fertilization. Soil Tillage Res. 2011, 113, 70–73. [Google Scholar] [CrossRef]
- Thomsen, I.K.; Christensen, B.T. Yields of wheat and soil carbon and nitrogen contents following long-term incorporation of barley straw and ryegrass catch crops. Soil Use Manag. 2004, 20, 432–438. [Google Scholar] [CrossRef]
- Lin, C.C.; Chen, D.W.; Dai, J.G. Advances of synthetic biology of flavonoids. Acta Pharm. Sin. 2022, 57, 1322–1335. [Google Scholar] [CrossRef]
- Ulbrich, B.; Zenk, M.H. Partial purification and properties of hydroxycinnamoyl-CoA: Quinate hydroxycinnamoyl transferase from higher plants. Phytochemistry 1979, 18, 929–933. [Google Scholar] [CrossRef]
- Villegas, R.J.; Kojima, M. Purification and characterization of hydroxycinnamoyl D-glucose. Quinate hydroxycinnamoyl transferase in the root of sweet potato, Ipomoea batatas Lam. J. Biol. Chem. 1986, 261, 8729–8733. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Ma, S.; Dai, M.; Feng, X.Y. Progress in Research on the biosynthesis pathway and metabolic regulation of phenolic acids. Food Sci. 2018, 39, 286–293. [Google Scholar] [CrossRef]
- Strissel, T.; Halbwirth, H.; Hoyer, U.; Zistler, C.; Stich, K.; Treutter, D. Growth-promoting nitrogen nutrition affects flavonoid biosynthesis in young apple (Malus domestica Borkh) leaves. Plant Biol. 2005, 7, 677–685. [Google Scholar] [CrossRef]
- Romanazzi, G.; Sanzani, S.M.; Bi, Y.; Tian, S.P.; Martínez, P.G.; Alkan, N. Induced resistance to control postharvest decay of fruit and vegetables. Postharvest Biol. Technol. 2016, 122, 82–94. [Google Scholar] [CrossRef]
- Ling, Y.; Qing, J.; Shang, X.L.; Yang, W.X.; Fang, S.Z. Effect of applying amount of nitrogen on growth and total phenolic accumulation in Cyclocarya paliurus seedling. J. Plant Resour. Environ. 2020, 29, 45–51. [Google Scholar] [CrossRef]
- Rivero, R.M.; Ruiz, J.M.; Romero, L. Can grafting in tomato plants strengthen resistance to thermal stress? J. Sci. Food Agric. 2003, 83, 1315–1319. [Google Scholar] [CrossRef]
- Jiao, X.L.; Gao, W.W. Advances in studies on influence of environmental factors on triterpenoid saponin synthesis in medicinal plants. Chin. Tradit. Herb. Drugs. 2011, 42, 398–420. [Google Scholar]
- Pecetti, L.; Tava, A.; Romani, M.; Benedetto, M.G.; Corsiet, P. Variety and environment effects on the dynamics of saponins in Lucerne (Medicago sativa L.). Eur. J. Agron. 2006, 25, 187–192. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | Organic C Content | Total N Content | C/N Ratio | pH | BET Surface Area | Total Pore Volume | t-Plot Mesopore Volume | t-Plot Micropore Area |
|---|---|---|---|---|---|---|---|---|
| g·kg−1 | m2·g−1 | cm3·g−1 | cm2·g−1 | |||||
| Poplar sawdust | 45.53 ± 0.06 c | 0.17 ± 0.00 b | 267.82 ± 10.78 b | 6.84 ± 0.18 c | 1.81 b | 0.003 b | 0.003 b | - |
| Biochar-350 °C | 74.09 ± 0.32 b | 0.19 ± 0.01 b | 385.42 ± 21.03 a | 7.43 ± 0.01 b | 2.18 b | 0.003 b | 0.003 b | - |
| Biochar-600 °C | 87.04 ± 0.43 a | 0.41 ± 0.04 a | 211.16 ± 12.54 c | 9.82 ± 0.31 a | 147.05 a | 0.08 a | 0.02 a | 108.28 |
| Treatment | Soil Bulk Density | Soil Porosity | ||
|---|---|---|---|---|
| Capillary Porosity | Non-Capillary Porosity | Total | ||
| (g·cm−3) | (%) | |||
| C0 | 0.98 ± 0.02 b | 44.93 ± 0.38 a | 7.97 ± 0.98 a | 52.93 ± 1.14 ab |
| C350 | 0.98 ± 0.01 b | 45.40 ± 1.21 a | 7.92 ± 0.57 a | 53.61 ± 1.62 ab |
| C600 | 0.97 ± 0.02 b | 45.99 ± 0.77 a | 7.41 ± 1.03 a | 54.41 ± 1.34 a |
| CK | 1.03 ± 0.02 a | 44.72 ± 1.40 a | 6.97 ± 0.52 a | 51.47 ± 0.88 b |
| Treatment | Month | pH | Organic C Content (g·kg−1) | Total N Content (g·kg−1) | C/N Ratio | Total Inorganic N Content (mg·kg−1) |
|---|---|---|---|---|---|---|
| C0 | July | 6.36 ± 0.14 a | 18.69 ± 1.06 b | 1.82 ± 0.16 a | 10.28 ± 0.79 c | 19.12 ± 1.19 d |
| C350 | 6.54 ± 0.23 a | 37.57 ± 0.45 a | 1.63 ± 0.12 ab | 23.06 ± 1.31 b | 79.25 ± 5.77 b | |
| C600 | 6.45 ± 0.31 a | 39.09 ± 0.99 a | 1.48 ± 0.69 b | 26.48 ± 0.96 a | 92.82 ± 8.79 a | |
| CK | 6.36 ± 0.16 a | 13.86 ± 1.87 c | 1.73 ± 0.23 ab | 7.46 ± 0.92 d | 49.59 ± 0.71 c | |
| C0 | October | 7.91 ± 0.06 b | 18.17 ± 2.71 c | 1.87 ± 0.15 a | 9.71 ± 1.04 c | 11.98 ± 0.77 a |
| C350 | 7.95 ± 0.15 b | 34.96 ± 4.85 b | 1.73 ± 0.22 a | 20.23 ± 2.91 b | 10.06 ± 0.72 bc | |
| C600 | 8.17 ± 0.06 a | 45.59 ± 2.08 a | 1.90 ± 0.17 a | 24.03 ± 1.30 a | 10.91 ± 0.44 b | |
| CK | 7.60 ± 0.02 c | 11.00 ± 0.96 d | 1.34 ± 0.10 b | 8.20 ± 0.25 d | 9.74 ± 0.29 c |
| Secondary Metabolite Monomers Content (mg·plant−1) | C0 | C350 | C600 | CK |
|---|---|---|---|---|
| Quercetin-3-O-glucuronide | 1.46 ± 0.51 a | 1.60 ± 0.16 a | 2.03 ± 0.54 a | 2.37 ± 0.21 a |
| Quercetin-3-O-galactoside | 5.18 ± 1.70 a | 3.51 ± 0.18 a | 5.08 ± 0.92 a | 3.75 ± 0.70 a |
| Isoquercitrin | 2.75 ± 0.36 b | 3.85 ± 0.79 ab | 4.49 ± 0.60 ab | 5.25 ± 0.57 a |
| Kaempferol-3-O-glucuronide | 3.15 ± 0.70 a | 3.10 ± 0.29 a | 5.66 ± 0.62 a | 3.052 ± 0.31 a |
| Kaempferol-3-O-glucoside | 0.93 ± 0.22 a | 0.96 ± 0.09 a | 1.13 ±0.08 a | 1.15 ± 0.20 a |
| Kaempferol-3-O-rhamnoside | 0.41 ± 0.08 b | 0.40 ± 0.03 b | 0.63 ± 0.07 a | 0.78 ± 0.05 a |
| Arjunolic acid | 0.29 ± 0.04 a | 0.39 ± 0.10 a | 0.70 ± 0.28 a | 0.44 ± 0.05 a |
| Cyclocaric acid B | 0.26 ± 0.04 c | 0.34 ± 0.03 bc | 0.56 ± 0.05 a | 0.42 ± 0.04 b |
| Pterocaryoside B | 5.49 ± 1.06 a | 5.09 ± 1.05 a | 6.12 ± 0.40 a | 5.44 ± 0.36 a |
| Pterocaryoside A | 0.34 ± 0.12 a | 0.32 ± 0.17 a | 0.42 ± 0.20 a | 0.38 ± 0.17 a |
| Hederagenin | 1.85 ± 0.07 a | 1.85 ± 0.83 a | 1.74 ± 0.19 a | 2.00 ± 0.07 a |
| Oleanolic acid | 0.02 ± 0.01 a | 0.25 ± 0.09 a | 0.24 ± 0.08 a | 0.29 ± 0.08 a |
| 3-O-caffeoylquinic acid | 10.77 ± 1.36 a | 10.61 ± 0.96 a | 15.72 ± 2.28 a | 11.21 ± 1.63 a |
| 4-O-caffeoylquinic acid | 12.72 ± 1.28 a | 16.08 ± 2.19 a | 17.71 ± 2.54 a | 16.17 ± 3.23 a |
| 4, 5-di-O-caffeoylquinic acid | 1.89 ± 0.33 a | 3.11 ± 0.39 a | 3.06 ± 0.84 a | 3.68 ± 0.60 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Tian, Y.; Liu, C.; Yang, R.; Jin, Y.; Li, T. Biochar Application: A Viable and Pyrolysis Temperature Dependent Option for Enhancing Leaf Secondary Metabolites of Cyclocarya paliurus. Forests 2023, 14, 1298. https://doi.org/10.3390/f14071298
Li W, Tian Y, Liu C, Yang R, Jin Y, Li T. Biochar Application: A Viable and Pyrolysis Temperature Dependent Option for Enhancing Leaf Secondary Metabolites of Cyclocarya paliurus. Forests. 2023; 14(7):1298. https://doi.org/10.3390/f14071298
Chicago/Turabian StyleLi, Wenhao, Ye Tian, Chengyu Liu, Rui Yang, Yuanyuan Jin, and Tong Li. 2023. "Biochar Application: A Viable and Pyrolysis Temperature Dependent Option for Enhancing Leaf Secondary Metabolites of Cyclocarya paliurus" Forests 14, no. 7: 1298. https://doi.org/10.3390/f14071298
APA StyleLi, W., Tian, Y., Liu, C., Yang, R., Jin, Y., & Li, T. (2023). Biochar Application: A Viable and Pyrolysis Temperature Dependent Option for Enhancing Leaf Secondary Metabolites of Cyclocarya paliurus. Forests, 14(7), 1298. https://doi.org/10.3390/f14071298