Author Contributions
Conceptualization, K.S.G., N.C.J., R.R.P. and A.J.A.; methodology, K.S.G. and A.J.A.; software, K.S.G. with support from A.J.A.; validation, K.S.G., D.A.N., N.C.J., R.R.P. and A.J.A.; formal analysis, K.S.G. with support from D.A.N. and A.J.A.; investigation, K.S.G. and A.J.A.; resources, Northern Arizona University, Valles Caldera National Preserve, K.S.G., N.C.J., R.R.P. and A.J.A.; data curation, K.S.G. and R.R.P.; writing—original draft preparation, K.S.G.; writing—review and editing, D.A.N., N.C.J., R.R.P. and A.J.A.; visualization, K.S.G. with support from A.J.A.; supervision, A.J.A.; project administration, K.S.G., N.C.J., R.R.P. and A.J.A.; funding acquisition, N.C.J., R.R.P. and A.J.A. All authors have read and agreed to the published version of the manuscript.
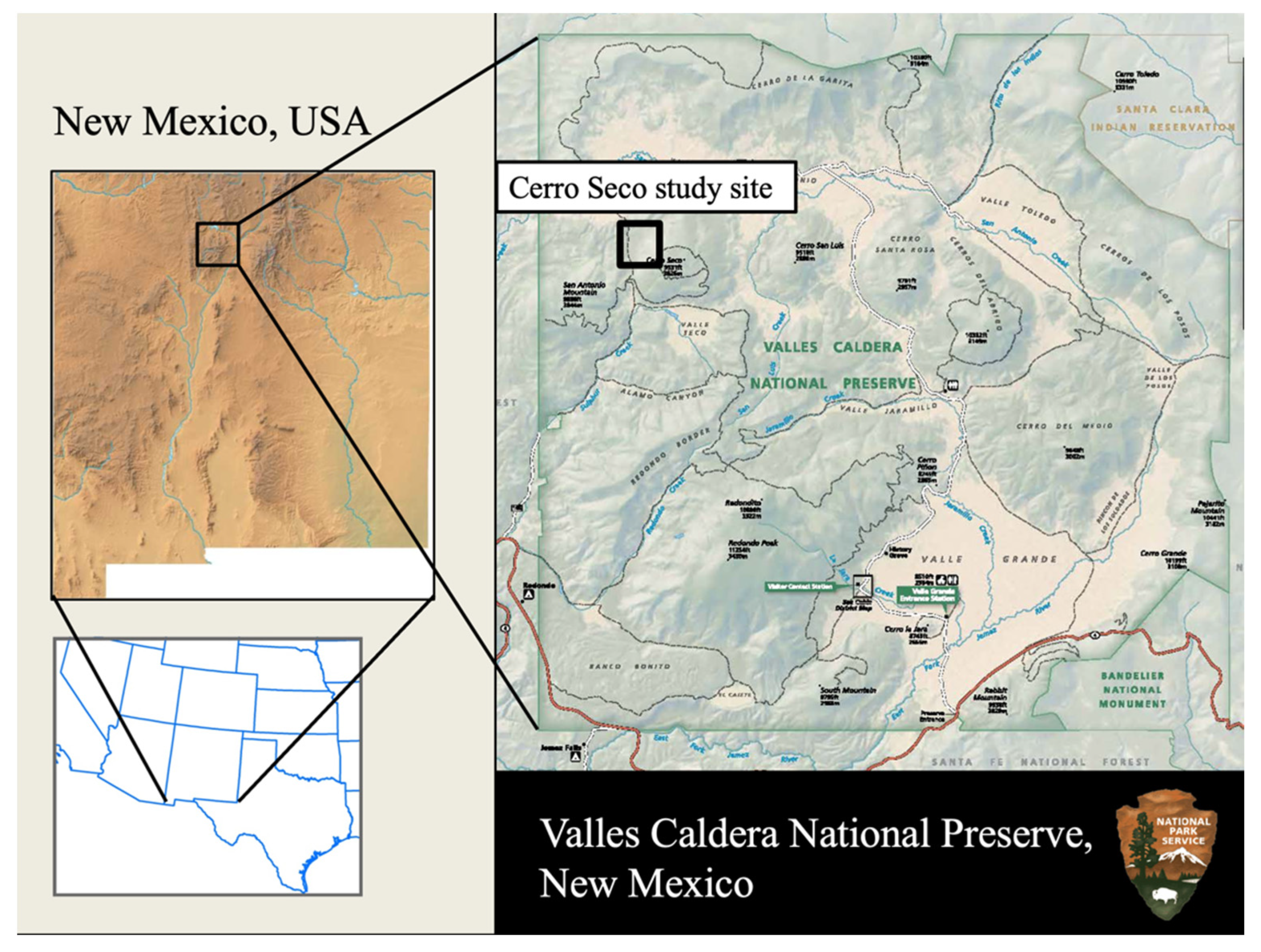
Figure 1.
Location of the Cerro Seco study site within Valles Caldera National Preserve and New Mexico, USA.
Figure 1.
Location of the Cerro Seco study site within Valles Caldera National Preserve and New Mexico, USA.
Figure 2.
One of three experimental feller buncher disturbance transects. Flags represent sampling points within and between the tracks (each cluster of three flags represents one sampling location). Each of the three transect blocks included three sections treated with 1, 3, and 9 passes, respectively. Samples and measurements were taken at three points per track and intertrack transect section (N = 54 sampling locations).
Figure 2.
One of three experimental feller buncher disturbance transects. Flags represent sampling points within and between the tracks (each cluster of three flags represents one sampling location). Each of the three transect blocks included three sections treated with 1, 3, and 9 passes, respectively. Samples and measurements were taken at three points per track and intertrack transect section (N = 54 sampling locations).
Figure 3.
(a) Soil surface resistance to penetration as measured with a pocket penetrometer (each observation is the mean of three replicate readings). (b) Shear strength as measured by TORVANE. Open boxplots show data from between the feller buncher tracks (I = intertrack), and filled boxplots show data from the tracks (T). Pink, purple, and blue represent one, three, and nine passes, respectively. The horizontal line within each box represents the median for that treatment, and the lower and upper bounds of the box indicate the 25th and 75th percentiles. Whiskers extend to 1.5× the interquartile range (IQR) from the lower and upper box bounds. Outlier observations more extreme than 1.5× the IQR are plotted as dots.
Figure 3.
(a) Soil surface resistance to penetration as measured with a pocket penetrometer (each observation is the mean of three replicate readings). (b) Shear strength as measured by TORVANE. Open boxplots show data from between the feller buncher tracks (I = intertrack), and filled boxplots show data from the tracks (T). Pink, purple, and blue represent one, three, and nine passes, respectively. The horizontal line within each box represents the median for that treatment, and the lower and upper bounds of the box indicate the 25th and 75th percentiles. Whiskers extend to 1.5× the interquartile range (IQR) from the lower and upper box bounds. Outlier observations more extreme than 1.5× the IQR are plotted as dots.
Figure 4.
Soil bulk densities (g/cm3) from four depths beneath the track and intertrack sampling locations that received 1, 3, or 9 passes (pink, blue and purple, respectively) from a feller buncher. Open boxplots show data from between the feller buncher tracks, and filled boxplots show data from the tracks. The horizontal line within each box represents the median for that treatment, and the lower and upper bounds of the box indicate the 25th and 75th percentiles, respectively. Whiskers extend to 1.5× the interquartile range from the lower and upper box bounds, and outlier observations are plotted as individual dots.
Figure 4.
Soil bulk densities (g/cm3) from four depths beneath the track and intertrack sampling locations that received 1, 3, or 9 passes (pink, blue and purple, respectively) from a feller buncher. Open boxplots show data from between the feller buncher tracks, and filled boxplots show data from the tracks. The horizontal line within each box represents the median for that treatment, and the lower and upper bounds of the box indicate the 25th and 75th percentiles, respectively. Whiskers extend to 1.5× the interquartile range from the lower and upper box bounds, and outlier observations are plotted as individual dots.
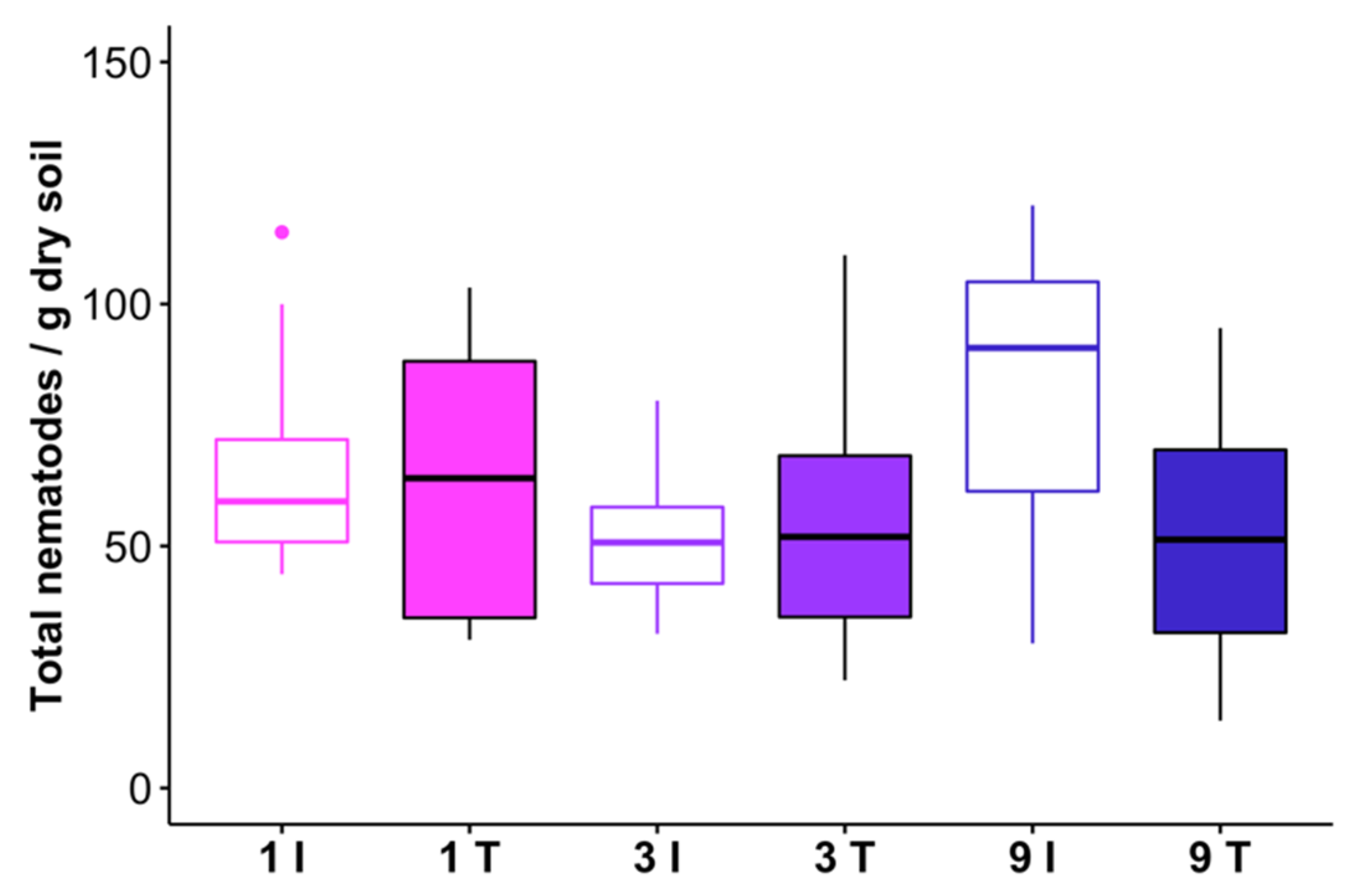
Figure 5.
Total nematode abundances in track (T) and intertrack (I) transect areas which received 1, 3, or 9 passes from the feller buncher. One outlier observation is not shown (276 nematodes/g dry soil in a 9 pass intertrack sample). The horizontal line within each box represents the treatment median, and the lower and upper bounds of the box indicate the 25th and 75th percentiles, respectively. Whiskers extend to 1.5× the interquartile range from the lower and upper box bounds. Outlier observations are represented as dots.
Figure 5.
Total nematode abundances in track (T) and intertrack (I) transect areas which received 1, 3, or 9 passes from the feller buncher. One outlier observation is not shown (276 nematodes/g dry soil in a 9 pass intertrack sample). The horizontal line within each box represents the treatment median, and the lower and upper bounds of the box indicate the 25th and 75th percentiles, respectively. Whiskers extend to 1.5× the interquartile range from the lower and upper box bounds. Outlier observations are represented as dots.
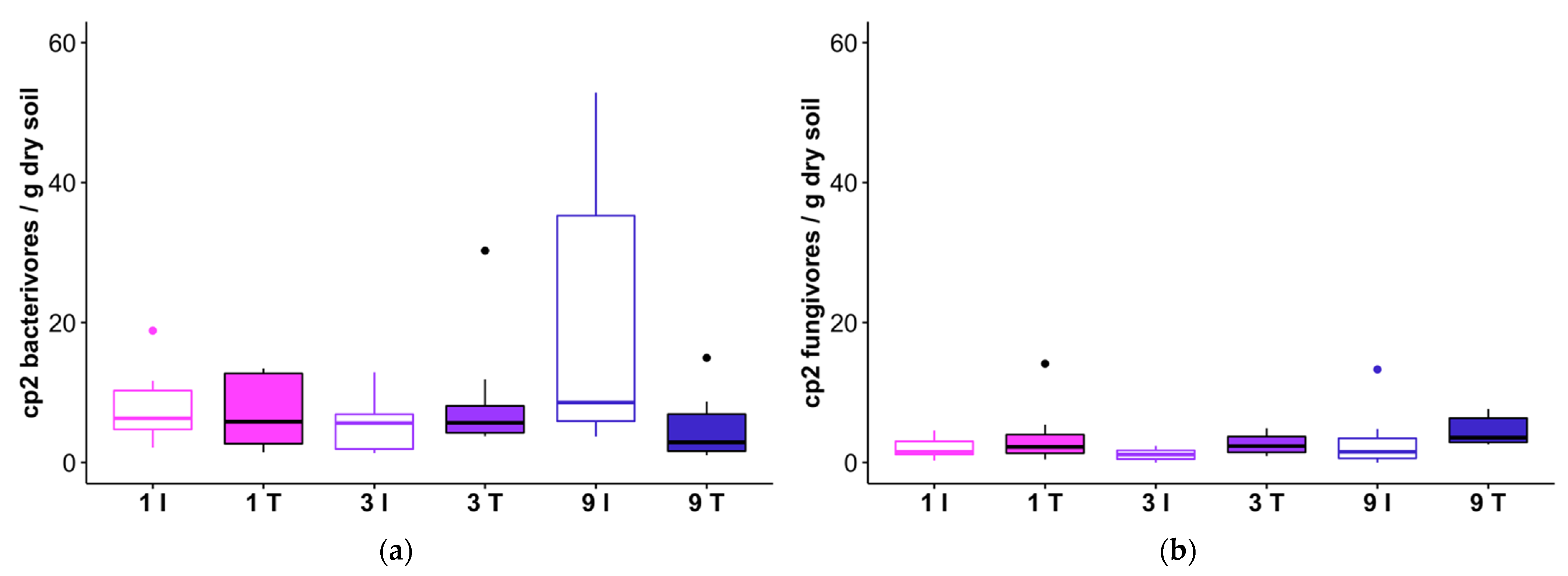
Figure 6.
Abundances of cp2 bacterivores (a) and cp2 fungivores (b) in track (T) and intertrack (I) transect areas which received 1, 3, or 9 passes from the feller buncher. The horizontal line within each box represents the median for that treatment combination, and the 25th and 75th percentiles for each treatment combination are delineated by the lower and upper bounds of the boxes, respectively. Whiskers extend to 1.5× the interquartile range.
Figure 6.
Abundances of cp2 bacterivores (a) and cp2 fungivores (b) in track (T) and intertrack (I) transect areas which received 1, 3, or 9 passes from the feller buncher. The horizontal line within each box represents the median for that treatment combination, and the 25th and 75th percentiles for each treatment combination are delineated by the lower and upper bounds of the boxes, respectively. Whiskers extend to 1.5× the interquartile range.
Figure 7.
Total abundances of cp3, cp4, and cp5 nematodes in feller buncher tracks (T) that received one, three, or nine passes, and matched intertrack (I) sampling locations (a); and correlations between total cp3, cp4, and cp5 nematodes and (b) bulk density at 9–12 cm, (c) surface penetration resistance, and (d) soil shear stress. (e) Harvester disturbance treatment responses of total cp3 and cp4 bacterivores. (f) Responses of Dorylaimida to harvester traffic. The horizontal line within each box represents the median for that treatment, and the lower and upper bounds of the box indicate the 25th and 75th percentiles, respectively. Whiskers extend to 1.5× the interquartile range from the lower and upper box bounds. Outlier observations are plotted as dots.
Figure 7.
Total abundances of cp3, cp4, and cp5 nematodes in feller buncher tracks (T) that received one, three, or nine passes, and matched intertrack (I) sampling locations (a); and correlations between total cp3, cp4, and cp5 nematodes and (b) bulk density at 9–12 cm, (c) surface penetration resistance, and (d) soil shear stress. (e) Harvester disturbance treatment responses of total cp3 and cp4 bacterivores. (f) Responses of Dorylaimida to harvester traffic. The horizontal line within each box represents the median for that treatment, and the lower and upper bounds of the box indicate the 25th and 75th percentiles, respectively. Whiskers extend to 1.5× the interquartile range from the lower and upper box bounds. Outlier observations are plotted as dots.
Figure 8.
Abundances of (
a) herbivore nematodes and (
b) nematodes in the family Tylenchidae (one observation of 174 Tylenchidae/g dry soil in a 9-intertrack treatment is not shown). The horizontal line within each box represents the median for that treatment, and the lower and upper bounds of the box indicate the 25th and 75th percentiles, respectively. Whiskers extend to 1.5× the interquartile range from the lower and upper box bounds, and outliers are plotted as dots. (
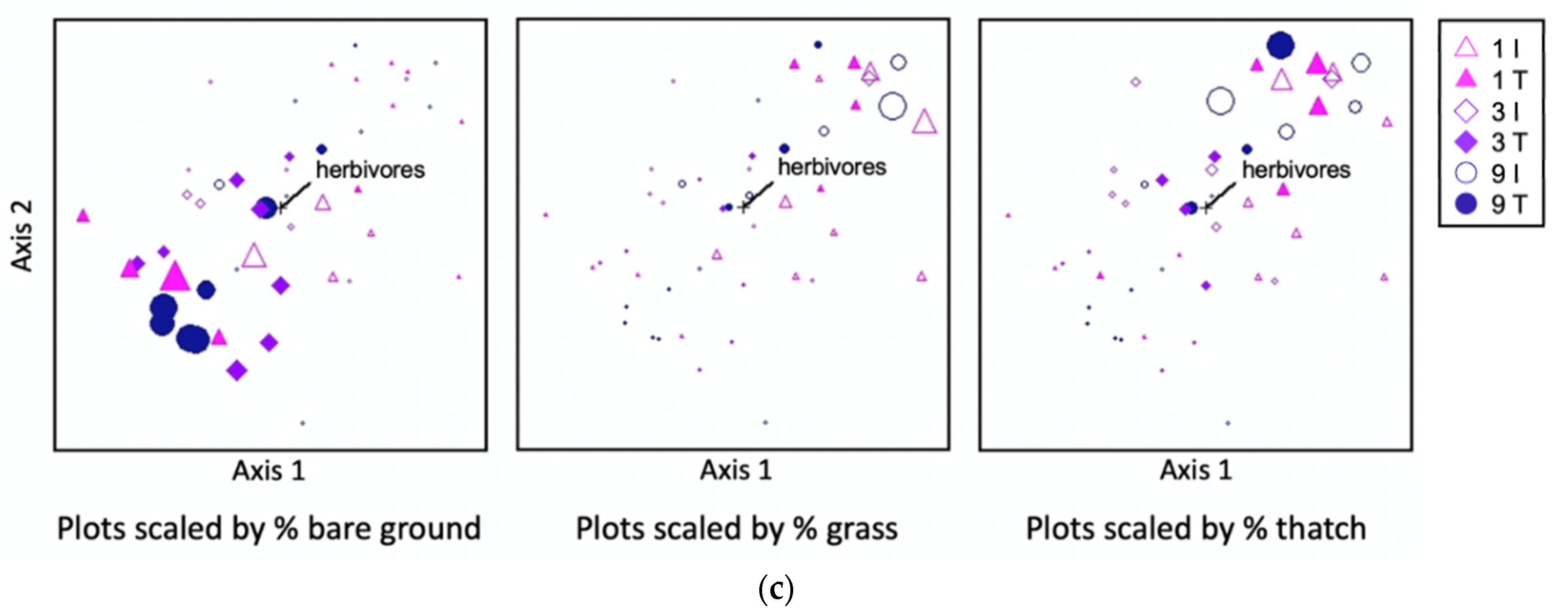
c) Non-metric multidimensional scaling (NMDS) ordination of ground cover, performed using Bray-Curtis distance, with points scaled according to percent bare ground, grass, and thatch (senesced grass) cover, respectively (larger points indicate greater cover by the category indicated below each panel; treatment means and standard errors for each cover class are listed in
Table 4). Herbivore abundance is shown as a vector overlay, indicating the relationship of herbivores with plot ground cover dissimilarity along axes 1 and 2 of the three-dimensional NMDS ordination. The direction of the line away from the center point indicates the direction of the association. I = intertrack, T = track; 1, 3, and 9 refer to a number of feller buncher passes received.
Figure 8.
Abundances of (
a) herbivore nematodes and (
b) nematodes in the family Tylenchidae (one observation of 174 Tylenchidae/g dry soil in a 9-intertrack treatment is not shown). The horizontal line within each box represents the median for that treatment, and the lower and upper bounds of the box indicate the 25th and 75th percentiles, respectively. Whiskers extend to 1.5× the interquartile range from the lower and upper box bounds, and outliers are plotted as dots. (
c) Non-metric multidimensional scaling (NMDS) ordination of ground cover, performed using Bray-Curtis distance, with points scaled according to percent bare ground, grass, and thatch (senesced grass) cover, respectively (larger points indicate greater cover by the category indicated below each panel; treatment means and standard errors for each cover class are listed in
Table 4). Herbivore abundance is shown as a vector overlay, indicating the relationship of herbivores with plot ground cover dissimilarity along axes 1 and 2 of the three-dimensional NMDS ordination. The direction of the line away from the center point indicates the direction of the association. I = intertrack, T = track; 1, 3, and 9 refer to a number of feller buncher passes received.


Table 1.
Feeding group and colonizer-persister (cp) class assignments for nematode taxa at the Cerro Seco study site in Valles Caldera National Preserve.
Table 1.
Feeding group and colonizer-persister (cp) class assignments for nematode taxa at the Cerro Seco study site in Valles Caldera National Preserve.
| Feeding Group(s) | cp Class(es) | Taxon |
|---|
| Bacterivores | cp1 | Panagrolaimidae |
| | | Rhabditidae |
| | cp2 | Cephalobidae |
| | | Plectidae |
| | cp3 | Monhysteridae * |
| | | Prismatolaimidae |
| | | Rhabdolaimidae |
| | | Teratocephalobidae |
| | cp4 | Alaimidae |
| Fungivores | cp2 | Aphelenchidae |
| | | Aphelenchoididae |
| Predators | cp4 | Mononchidae |
| Omnivores/predators/fungivores | cp4/cp5 | Dorylaimida |
| Herbivores | NA | Criconematidae |
| | | Hoplolaimidae |
| | | Hemicyclophoridae |
| | | Trichodoridae |
| | | Other Tylenchoidea (apart from Tylenchidae) |
| Plant associates | cp2 | Tylenchidae |
Table 2.
Non-parametric ANOVA-type analysis of soil penetration resistance and shear strength; ATS = ANOVA-type test statistic; df = numerator degrees of freedom; passes = number of passes applied to the transect segment (1, 3, or 9).
Table 2.
Non-parametric ANOVA-type analysis of soil penetration resistance and shear strength; ATS = ANOVA-type test statistic; df = numerator degrees of freedom; passes = number of passes applied to the transect segment (1, 3, or 9).
| Penetration Resistance | | | |
|---|
| | df | ATS | p |
|---|
| Passes | 1.92 | 0.25 | 0.770 |
| Treatment (track vs. intertrack) | 1.00 | 22.54 | <0.001 *** |
| Passes × Treatment | 1.89 | 2.24 | 0.109 |
| Shear Strength | | | |
| | df | ATS | p |
| Passes | 1.78 | 0.78 | 0.443 |
| Treatment (track vs. intertrack) | 1.00 | 7.77 | 0.005 ** |
| Passes × Treatment | 1.94 | 0.42 | 0.653 |
Table 3.
Non-parametric ANOVA-type analysis of bulk density; ATS = ANOVA-type test statistic; df = numerator degrees of freedom; passes = number of passes applied to the transect segment (1, 3, or 9).
Table 3.
Non-parametric ANOVA-type analysis of bulk density; ATS = ANOVA-type test statistic; df = numerator degrees of freedom; passes = number of passes applied to the transect segment (1, 3, or 9).
| Bulk Density | | | |
|---|
| | df | ATS | p |
|---|
| Passes | 1.99 | 1.21 | 0.298 |
| Treatment (track vs. intertrack) | 1.00 | 15.99 | <0.001 *** |
| Depth | 2.52 | 6.70 | <0.001 *** |
| Passes × Treatment | 1.93 | 2.12 | 0.122 |
| Depth × Treatment | 2.62 | 3.06 | 0.033 * |
| Passes × Depth | 4.69 | 1.91 | 0.094 |
| Passes × Treatment × Depth | 4.45 | 0.45 | 0.795 |
Table 4.
Litter depth, trench depth (depth of the nearest indentation formed by the feller buncher tracks), and ground cover measured in intertrack (I) and track (T) quadrats from the 1, 3, and 9 pass segments of the experimental transects. Ground cover classes are listed in descending order of their mean cover within the intertrack quadrats. Thatch = senesced grass.
Table 4.
Litter depth, trench depth (depth of the nearest indentation formed by the feller buncher tracks), and ground cover measured in intertrack (I) and track (T) quadrats from the 1, 3, and 9 pass segments of the experimental transects. Ground cover classes are listed in descending order of their mean cover within the intertrack quadrats. Thatch = senesced grass.
| | 1 I | 1 T | 3 I | 3 T | 9 I | 9 T | I (Overall) | T (Overall) |
|---|
| | Mean (SE) | Mean (SE) | Mean (SE) | Mean (SE) | Mean (SE) | Mean (SE) | Mean (SE) | Mean (SE) |
|---|
| Litter depth (cm) | 2.7 (0.8) | 2.7 (0.8) | 1.8 (0.3) | 1.9 (0.5) | 3.4 (1) | 1.7 (0.5) | 2.6 (0.4) | 2.1 (0.3) |
| Trench depth (cm) | NA | 5.7 (0.7) | NA | 5 (0.6) | NA | 5.3 (0.9) | NA | 5.3 (0.4) |
| Thatch (%) | 30.6 (7.9) | 26.1 (8.7) | 27.8 (7.2) | 15 (4.8) | 26.1 (10.7) | 22.8 (10.3) | 28.1 (4.8) | 21.3 (4.7) |
| Moss (%) | 2.8 (1.5) | 4.4 (2.6) | 28.3 (6) | 17.2 (7.7) | 30.6 (12) | 8.9 (3.4) | 20.6 (5) | 10.2 (3) |
| Grass (%) | 28.1 (7.7) | 10.8 (3.8) | 9.3 (4.9) | 3.1 (1.3) | 23 (9.6) | 8.3 (3.6) | 20.1 (4.5) | 7.4 (1.9) |
| Pine litter (%) | 20 (7.3) | 11.1 (4.6) | 7.2 (2.9) | 7.8 (3.4) | 8.3 (3.4) | 5.6 (1.8) | 11.9 (3) | 8.1 (2) |
| Bare ground (%) | 17.4 (9) | 30 (10.6) | 6.1 (2.9) | 45.9 (5.1) | 10 (7.8) | 55 (12.5) | 11.2 (4) | 43.6 (5.9) |
| Spruce litter (%) | 0.2 (0.2) | 6.1 (3.5) | 15.6 (5.6) | 8.3 (4.9) | 14.4 (8.8) | 3.3 (2.4) | 10.1 (3.6) | 5.9 (2.1) |
| Lichen (%) | 6.9 (2.4) | 5.7 (4.4) | 4.4 (1.9) | 5 (1.8) | 2.8 (1.2) | 1.1 (1.1) | 4.7 (1.1) | 3.9 (1.6) |
| Forb (%) | 1.4 (1.1) | 1.7 (1.2) | 3 (1.2) | 0.4 (0.3) | 2.8 (2.2) | 2.2 (1.2) | 2.4 (0.9) | 1.4 (0.6) |
| Wood mulch (%) | 3 (1.9) | 10.6 (4.5) | 2.8 (2.2) | 2.2 (1.7) | 1.1 (0.7) | 6.1 (5.5) | 2.3 (1) | 6.3 (2.4) |
| Sticks (%) | 0.6 (0.6) | 0.6 (0.6) | 3.9 (2.3) | 2.7 (1.2) | 1.7 (1.2) | 3.6 (1.4) | 2 (0.9) | 2.3 (0.7) |
| Moss litter (%) | 2.2 (1.5) | 0 (0) | 1.7 (1.7) | 5.1 (3.3) | 0 (0) | 0 (0) | 1.3 (0.7) | 1.7 (1.2) |
| Shrub (%) | 0 (0) | 0 (0) | 0 (0) | 0.4 (0.4) | 0.6 (0.6) | 0 (0) | 0.2 (0.2) | 0.1 (0.1) |
| Scat (%) | 0 (0) | 0 (0) | 0.2 (0.2) | 0.2 (0.2) | 0 (0) | 0 (0) | 0.1 (0.1) | 0.1 (0.1) |
| Forb litter (%) | 0 (0) | 3.3 (3.3) | 0 (0) | 1.1 (1.1) | 0 (0) | 0 (0) | 0 (0) | 1.5 (1.2) |
Table 5.
Multi-response permutation procedure (MRPP) results for ground cover, performed using Bray–Curtis distance. A is the chance-corrected proportion of between-sample distances explainable by the treatment group.
Table 5.
Multi-response permutation procedure (MRPP) results for ground cover, performed using Bray–Curtis distance. A is the chance-corrected proportion of between-sample distances explainable by the treatment group.
| | A | p |
|---|
| Overall group differences | 0.0927 | <0.001 *** |
| 1 pass (intertrack vs. track) | −0.007 | 0.500 |
| 3 passes (intertrack vs. track) | 0.0898 | 0.002 ** |
| 9 passes (intertrack vs. track) | 0.1234 | 0.004 ** |
Table 6.
Non-parametric ANOVA-type analysis of total nematode abundance; ATS = ANOVA-type test statistic; df = numerator degrees of freedom; passes = number of passes applied to the transect segment (1, 3, or 9).
Table 6.
Non-parametric ANOVA-type analysis of total nematode abundance; ATS = ANOVA-type test statistic; df = numerator degrees of freedom; passes = number of passes applied to the transect segment (1, 3, or 9).
| Total Nematodes | | | |
|---|
| | df | ATS | p |
|---|
| Passes | 1.97 | 0.66 | 0.513 |
| Treatment (track vs. intertrack) | 1.00 | 2.04 | 0.154 |
| Passes × Treatment | 1.99 | 1.93 | 0.145 |
Table 7.
Results of non-parametric ANOVA-type analysis of cp2 bacterivore and cp2 fungivore abundances; ATS = ANOVA-type test statistic; df = numerator degrees of freedom; passes = number of passes applied to the transect segment (1, 3, or 9).
Table 7.
Results of non-parametric ANOVA-type analysis of cp2 bacterivore and cp2 fungivore abundances; ATS = ANOVA-type test statistic; df = numerator degrees of freedom; passes = number of passes applied to the transect segment (1, 3, or 9).
| cp2 Bacterivores | | | |
|---|
| | df | ATS | p |
|---|
| Passes | 2.00 | 0.09 | 0.911 |
| Treatment (track vs. intertrack) | 1.00 | 1.84 | 0.175 |
| Passes × Treatment | 1.95 | 3.88 | 0.022 * |
| cp2 Fungivores | | | |
| | df | ATS | p |
| Passes | 1.97 | 1.77 | 0.172 |
| Treatment (track vs. intertrack) | 1.00 | 10.16 | 0.001 ** |
| Passes × Treatment | 1.63 | 0.85 | 0.407 |
Table 8.
Non-parametric ANOVA-type analysis of total cp3, cp4, and cp5 nematodes; cp3 and cp4 bacterivores; and Dorylaimida; ATS = ANOVA-type test statistic; df = numerator degrees of freedom; passes = number of passes applied to the transect segment (1, 3, or 9).
Table 8.
Non-parametric ANOVA-type analysis of total cp3, cp4, and cp5 nematodes; cp3 and cp4 bacterivores; and Dorylaimida; ATS = ANOVA-type test statistic; df = numerator degrees of freedom; passes = number of passes applied to the transect segment (1, 3, or 9).
| Total cp3, cp4, and cp5 Nematodes | | | |
|---|
| | df | ATS | p |
|---|
| Passes | 1.93 | 1.49 | 0.227 |
| Treatment (track vs. intertrack) | 1.00 | 3.40 | 0.065 |
| Passes × Treatment | 1.92 | 3.77 | 0.025 * |
| cp3 and cp4 Bacterivores | | | |
| | df | ATS | p |
| Passes | 1.77 | 0.26 | 0.747 |
| Treatment (track vs. intertrack) | 1.00 | 5.49 | 0.019 * |
| Passes × Treatment | 1.84 | 4.92 | 0.009 ** |
| Dorylaimida | | | |
| | df | ATS | p |
| Passes | 2.00 | 1.99 | 0.137 |
| Treatment (track vs. intertrack) | 1.00 | 3.21 | 0.073 |
| Passes × Treatment | 1.84 | 3.93 | 0.023 * |
Table 9.
Non-parametric ANOVA-type analysis of herbivore nematodes and nematodes in the family Tylenchidae. ATS = ANOVA-type test statistic; df = numerator degrees of freedom; passes = number of passes applied to the transect segment (1, 3, or 9).
Table 9.
Non-parametric ANOVA-type analysis of herbivore nematodes and nematodes in the family Tylenchidae. ATS = ANOVA-type test statistic; df = numerator degrees of freedom; passes = number of passes applied to the transect segment (1, 3, or 9).
| Herbivores | | | |
|---|
| | df | ATS | p |
|---|
| Passes | 1.80 | 2.65 | 0.077 |
| Treatment (track vs. intertrack) | 1.00 | 0.60 | 0.439 |
| Passes × Treatment | 1.63 | 1.01 | 0.353 |
| Tylenchidae | | | |
| | df | ATS | p |
| Passes | 1.98 | 0.25 | 0.779 |
| Treatment (track vs. intertrack) | 1.00 | 2.17 | 0.141 |
| Passes × Treatment | 1.69 | 0.45 | 0.606 |



 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}