
Development of an Efficient and Rapid Micropropagation Protocol for In Vitro Multiplication of Maerua crassifolia Forssk


 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Surface Sterilization
2.2. In Vitro Germination of Seeds
2.3. Cultivation Media, Growth Regulators, and Culture Conditions
2.4. Rooting
2.5. Acclimatization
2.6. Flow Cytometric (FCM) Analysis
2.7. Data Analysis
3. Results
3.1. Effect of Cytokinins on the Morphogenetic Response of Hypocotyls
3.2. Synergistic Effect of Cytokinins and Auxins on the Morphogenetic Response of Hypocotyls
3.3. Rooting of the In Vitro Generated Shoots
3.4. Acclimatization
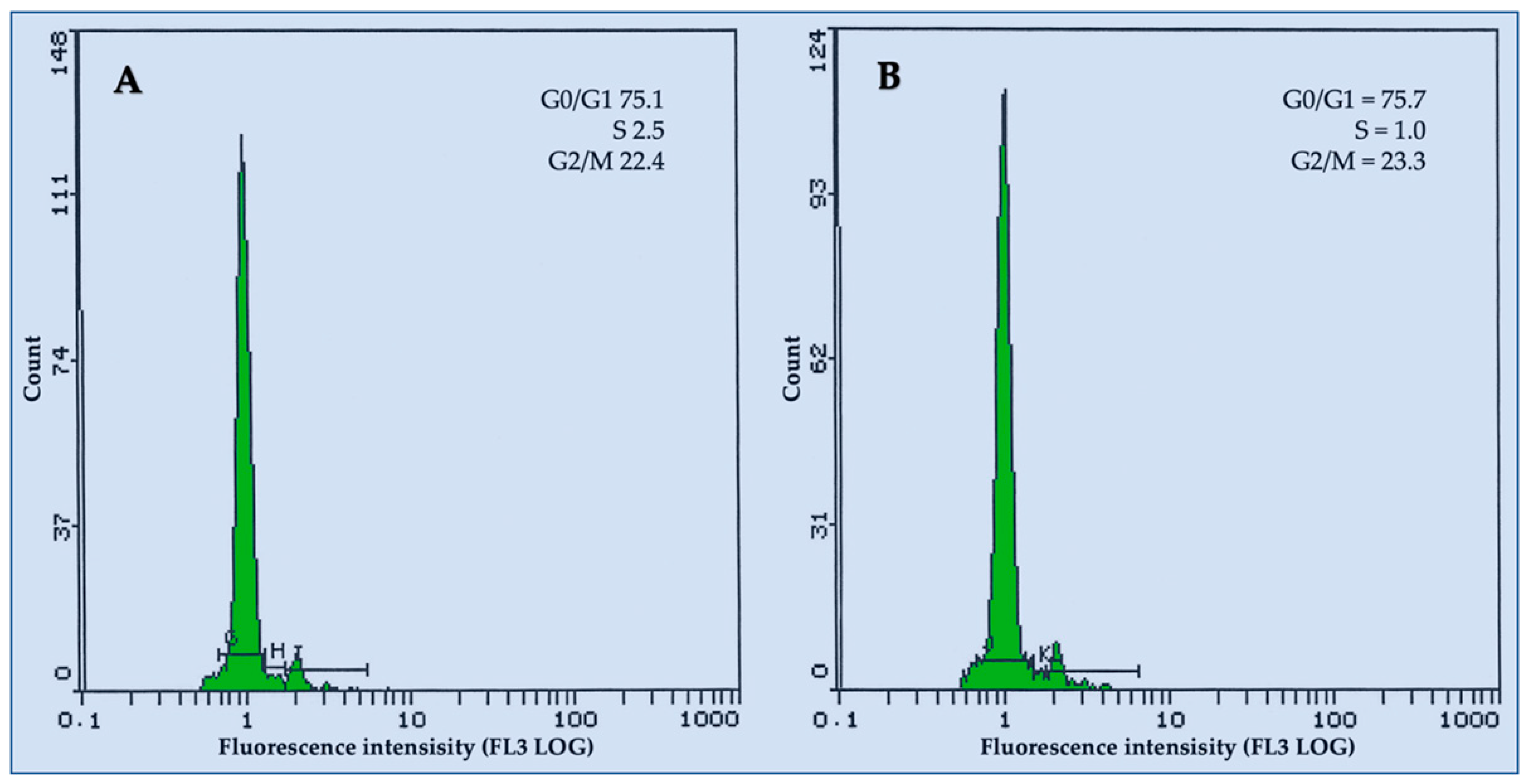
3.5. Flow Cytometric Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Elaidarous, A.A.; Osman, H.E.; Galal, T.M.; El-Morsy, M.H. Vegetation–environment relationship and floristic diversity of Wadi Al-Sharaea, Makkah Province, Saudi Arabia. Rend. Lincei. Sci. Fis. Nat. 2022, 33, 169–184. [Google Scholar] [CrossRef]
- Yonbawi, A.R.; Abdallah, H.M.; Alkhilaiwi, F.A.; Koshak, A.E.; Heard, C.M. Anti-Proliferative, Cytotoxic and Antioxidant Properties of the Methanolic Extracts of Five Saudi Arabian Flora with Folkloric Medicinal Use: Aizoon canariense, Citrullus colocynthis, Maerua crassifolia, Rhazya stricta and Tribulus macropterus. Plants 2021, 10, 2073. [Google Scholar] [CrossRef] [PubMed]
- Volpato, G.; Kourková, P.; Zelený, V. Healing war wounds and perfuming exile: The use of vegetal, animal, and mineral products for perfumes, cosmetics, and skin healing among Sahrawi refugees of Western Sahara. J. Ethnobiol. Ethnomed. 2012, 8, 49. [Google Scholar] [CrossRef] [PubMed]
- Christian, A.G.; Akanimo, E.G.; Ahunna, A.G.; Nwakaego, E.M.; Chimsorom, C.K. Antimalarial potency of the methanol leaf extract of Maerua crassifolia Forssk (Capparaceae). Asian Pac. J. Trop. Dis. 2014, 4, 35–39. [Google Scholar] [CrossRef]
- Akuodor, G.C.; Essien, A.D.; Akpan, J.L.; Chilaka, K.C.; Uwaezuoke, N.J.I.; Nwadike, K.I.; Nwobodo, N.N.; Ezeokpo, B.C. Analgesic, antiinflammatory and antipyretic activities of methanolic leaf extract of Maerua crassifolia. J. Coast. Life Med. 2016, 4, 225–230. [Google Scholar] [CrossRef]
- Kingsley Chimsorom Ckilaka, G.C.A.; Akpan, J.L.; Ogiji, E.D.; Eze, C.O.; Ezeokpo, B.C. Antibacterial and antioxidant activities of methanolic leaf extract of Maerua crassifolia. J. Appl. Pharm. Sci. 2015, 5, 147–150. [Google Scholar] [CrossRef]
- Chaib, F.; Sahki, R.; Sabaou, N.; Rached, W.; Bennaceur, M. Phytochemical investigation and biological activities of some Saharan plants from Hoggar. J. Agric. Sci. 2015, 7, 163–173. [Google Scholar] [CrossRef]
- Qahtan, A.A.; Faisal, M.; Alatar, A.A.; Abdel-Salam, E.M. High-Frequency Plant Regeneration, Genetic Uniformity, and Flow Cytometric Analysis of Regenerants in Ruta chalepensis L. Plants 2021, 10, 2820. [Google Scholar] [CrossRef]
- Bakhtiar, Z.; Mirjalili, M.H.; Sonboli, A. In vitro callus induction and micropropagation of Thymus persicus (Lamiaceae), an endangered medicinal plant. Crop Breed. Appl. Biotechnol. 2016, 16, 48–54. [Google Scholar] [CrossRef]
- Roy, A.; Kundu, K.; Saxena, G.; Kumar, L.; Bharadvaja, N. Effect of different media and growth hormones on shoot multiplication of in vitro grown Centella asiatica accessions. Adv. Tech. Biol. Med. 2016, 4, 1–4. [Google Scholar] [CrossRef]
- Hussain, S.A.; Anis, M.; Alatar, A.A. Efficient In Vitro Regeneration System for Tecoma stans L., Using Shoot Tip and Assessment of Genetic Fidelity Among Regenerants. Proc. Natl. Acad. Sci. India Sect. Biol. Sci. 2020, 90, 171–178. [Google Scholar] [CrossRef]
- Hussain, S.A.; Ahmad, N.; Anis, M.; Hakeem, K.R. Development of an efficient micropropagation system for Tecoma stans (L.) Juss. ex Kunth using thidiazuron and effects on phytochemical constitution. Vitr. Cell. Dev. Biol. Plant 2019, 55, 442–453. [Google Scholar] [CrossRef]
- Saad, A.A.M.; Asalam, A.A. Culture of leaves and immature fruits of Maerua crassifolia in vitro. Int. J. Inf. Res. Rev. 2015, 2, 496–498. [Google Scholar]
- Al-Qurainy, F.; Nadeem, M.; Khan, S.; Alansi, S.; Tarroum, M.; Al-Ameri, A.; Gaafar, A.; Alshameri, A. Micropropagation and evaluation of genetic fidelity of Maerua oblongifolia (FORSSK.) A. RICH: A rare medicinal plant from Saudi Arabia. Fresenius Environ. Bull. 2018, 27, 165–171. [Google Scholar]
- Rathore, M.S.; Shekhawat, N.S. Micropropagation of Maerua oblongifolia: A rare ornamental from semi arid regions of Rajasthan, India. J. Dev. Biol. Tissue Eng. 2011, 3, 92–98. [Google Scholar]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Galbraith, D.W.; Harkins, K.R.; Maddox, J.M.; Ayres, N.M.; Sharma, D.P.; Firoozabady, E. Rapid Flow Cytometric Analysis of the Cell Cycle in Intact Plant Tissues. Science 1983, 220, 1049. [Google Scholar] [CrossRef]
- Liu, R.; Xue, Y.; Ci, H.; Gao, J.; Wang, S.; Zhang, X. Establishment of highly efficient plant regeneration of Paeonia ostii ‘Fengdan’ through optimization of callus, adventitious shoot, and rooting induction. Hortic. Plant J. 2022, 8, 777–786. [Google Scholar] [CrossRef]
- de Oliveira, L.S.; Brondani, G.E.; Molinari, L.V.; Dias, R.Z.; Teixeira, G.L.; Gonçalves, A.N.; de Almeida, M. Optimal cytokinin/auxin balance for indirect shoot organogenesis of Eucalyptus cloeziana and production of ex vitro rooted micro-cuttings. J. For. Res. 2022, 33, 1573–1584. [Google Scholar] [CrossRef]
- Naaz, A.; Hussain, S.A.; Naz, R.; Anis, M.; Alatar, A.A. Successful plant regeneration system via de novo organogenesis in Syzygium cumini (L.) Skeels: An important medicinal tree. Agrofor. Syst. 2019, 93, 1285–1295. [Google Scholar] [CrossRef]
- Magyar-Tábori, K.; Dobránszki, J.; Teixeira da Silva, J.A.; Bulley, S.M.; Hudák, I. The role of cytokinins in shoot organogenesis in apple. Plant Cell Tiss. Org. Cult. 2010, 101, 251–267. [Google Scholar] [CrossRef]
- Tomasiak, A.; Zhou, M.; Betekhtin, A. Buckwheat in Tissue Culture Research: Current Status and Future Perspectives. Int. J. Mol. Sci. 2022, 23, 2298. [Google Scholar] [CrossRef]
- Ebrahimzadeh, M.H.; Naseri, M.; Younesikelaki, F.S.; Yusefi Javan, I.; Swamy, N.R. Influence of Plant Growth Regulators on Callus Induction, Silymarin Production and Antioxidant Activity in Silybum marianum L. Gaertn. by Tissue Culture. Int. J. Hortic. Sci. Technol. 2023, 10, 223–236. [Google Scholar]
- Doğru, S.M.; Balkaya, A.; Kurtar, E.S. In vitro Micropropagation of Maintainer White Head Cabbage Lines Using Cotyledon and Hypocotyl Explants. Black Sea J. Agric. 2022, 5, 180–188. [Google Scholar] [CrossRef]
- Bedir, H.; Ari, E.; Vural, G.E.; Seguí-Simarro, J.M. Effect of the genotype, explant source and culture medium in somatic embryogenesis and organogenesis in Vaccaria hispanica (Mill.) Rauschert. Plant Cell Tiss. Org. Cult. 2022, 150, 329–343. [Google Scholar] [CrossRef]
- George, E.F.; Hall, M.A.; Klerk, G.-J.D. Plant Growth Regulators I: Introduction; Auxins, their Analogues and Inhibitors. In Plant Propagation by Tissue Culture: Volume 1. The Background; George, E.F., Hall, M.A., Klerk, G.-J.D., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 175–204. [Google Scholar] [CrossRef]
- Hesami, M.; Daneshvar, M.H.; Yoosefzadeh-Najafabadi, M.; Alizadeh, M. Effect of plant growth regulators on indirect shoot organogenesis of Ficus religiosa through seedling derived petiole segments. J. Genet. Eng. Biotechnol. 2018, 16, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.A.; Nathar, V.N. In Vitro Method of High-Frequency Plant Regeneration Through Internodal Callus of Ruta graveolens L. In Medicinal Plants: Biodiversity, Sustainable Utilization and Conservation; Khasim, S.M., Long, C., Thammasiri, K., Lutken, H., Eds.; Springer: Singapore, 2020; pp. 761–768. [Google Scholar] [CrossRef]
- Patricia, D.; Stephen, B.; John, A. Shoot organogenesis from leaf discs of the African ginger (Mondia whitei (Hook.f.) Skeels), an endangered medicinal plant. Vitr. Cell. Dev. Biol. Plant 2021, 57, 493–498. [Google Scholar] [CrossRef]
- Aslam, M.M.; Karanja, J.K.; Zhang, Q.; Lin, H.; Xia, T.; Akhtar, K.; Liu, J.; Miao, R.; Xu, F.; Xu, W. In Vitro Regeneration Potential of White Lupin (Lupinus albus) from Cotyledonary Nodes. Plants 2020, 9, 318. [Google Scholar] [CrossRef]
- Estrella-Maldonado, H.; Talavera-May, C.R.; Fuentes Ortìz, G.; Desjardins, Y.; Santamarìa, J.M. Rhizogenesis on in-vitro plantlets of Carica papaya L.: Identification and expression profiling of transcription repressors of response to auxin (Aux/IAA) and auxin response factor (ARF) genes. Vitr. Int. Symp. Papaya 2019, 1250, 153–158. [Google Scholar] [CrossRef]
- Fogaça, C.M.; Fett-Neto, A.G. Role of auxin and its modulators in the adventitious rooting of Eucalyptus species differing in recalcitrance. Plant Growth Regul. 2005, 45, 1–10. [Google Scholar] [CrossRef]
- Ludwig-Müller, J. Indole-3-butyric acid in plant growth and development. Plant Growth Regul. 2000, 32, 219–230. [Google Scholar] [CrossRef]
- Pasqual, M.; Pio, L.A.S.; Oliveira, A.C.L.; Soares, J.D.R. Flow cytometry applied in tissue culture. In Recent Advances in Plant In Vitro Culture; Leva, A., Laura, M., Rinaldi, R., Eds.; InTech: Rijeka, Croatia, 2012; pp. 109–122. [Google Scholar]
- Singh, S.R.; Dalal, S.; Singh, R.; Dhawan, A.K.; Kalia, R.K. Evaluation of genetic fidelity of in vitro raised plants of Dendrocalamus asper (Schult. & Schult. F.) Backer ex K. Heyne using DNA-based markers. Acta Physiol. Plant 2013, 35, 419–430. [Google Scholar] [CrossRef]
- Thiem, B.; Kikowska, M.; Krawczyk, A.; Więckowska, B.; Sliwinska, E. Phenolic acid and DNA contents of micropropagated Eryngium planum L. Plant Cell Tiss. Org. Cult. 2013, 114, 197–206. [Google Scholar] [CrossRef]
- Nybom, H.; Weising, K.; Rotter, B. DNA fingerprinting in botany: Past, present, future. Investig. Genet. 2014, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Faisal, M.; Alatar, A.A.; El-Sheikh, M.A.; Abdel-Salam, E.M.; Qahtan, A.A. Thidiazuron induced in vitro morphogenesis for sustainable supply of genetically true quality plantlets of Brahmi. Ind. Crops Prod. 2018, 118, 173–179. [Google Scholar] [CrossRef]
- Brito, G.; Lopes, T.; Loureiro, J.; Rodriguez, E.; Santos, C. Assessment of genetic stability of two micropropagated wild olive species using flow cytometry and microsatellite markers. Trees 2010, 24, 723–732. [Google Scholar] [CrossRef]
- Liu, L.-S.; Li, R.; Zhao, Y.; Wen, C.-L.; Ren, S.; Guo, Y.-D. High efficiency regeneration and genetic stability analysis of somatic clones of Gynura bicolor DC. Afr. J. Biotechnol. 2011, 10, 10380–10386. [Google Scholar]
- Konar, S.; Karmakar, J.; Ray, A.; Adhikari, S.; Bandyopadhyay, T.K. Regeneration of plantlets through somatic embryogenesis from root derived calli of Hibiscus sabdariffa L. (Roselle) and assessment of genetic stability by flow cytometry and ISSR analysis. PLoS ONE 2018, 13, e0202324. [Google Scholar] [CrossRef]
- Jena, S.; Ray, A.; Sahoo, A.; Sahoo, S.; Dash, B.; Kar, B.; Nayak, S. Rapid plant regeneration in industrially important Curcuma zedoaria revealing genetic and biochemical fidelity of the regenerants. 3 Biotech. 2019, 10, 17. [Google Scholar] [CrossRef]
- Conde, P.; Loureiro, J.; Santos, C. Somatic embryogenesis and plant regeneration from leaves of Ulmus minor Mill. Plant Cell Rep. 2004, 22, 632–639. [Google Scholar] [CrossRef]
- Loureiro, J.; Capelo, A.; Brito, G.; Rodriguez, E.; Silva, S.; Pinto, G.; Santos, C. Micropropagation of Juniperus phoenicea from adult plant explants and analysis of ploidy stability using flow cytometry. Biol. Plant. 2007, 51, 7–14. [Google Scholar] [CrossRef]
- Oliveira, L.S.; Xavier, A.; Otoni, W.C.; Campos, J.M.S.; Viccini, L.F.; Takahashi, E.K. Assessment of genetic stability of micropropagated Eucalyptus globulus Labill hybrid clones by means of flow cytometry and microsatellites markers. Rev. Árvore 2017, 41, e410114. [Google Scholar] [CrossRef][Green Version]
- Huang, H.; Wei, Y.; Zhai, Y.; Ouyang, K.; Chen, X.; Bai, L. High frequency regeneration of plants via callus-mediated organogenesis from cotyledon and hypocotyl cultures in a multipurpose tropical tree (Neolamarkia Cadamba). Sci. Rep. 2020, 10, 4558. [Google Scholar] [CrossRef] [PubMed]
- Ulvrova, T.; Vitamvas, J.; Cepkova, P.H.; Eliasova, K.; Janovska, D.; Bazant, V.; Viehmannova, I. Micropropagation of an ornamental shrub Disanthus cercidifolius Maxim. and assessment of genetic fidelity of regenerants using ISSR and flow cytometry. Plant Cell Tiss. Org. Cult. 2021, 144, 555–566. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cytokinins (μM) | Response (%) 1 | Mean No. of Shoots | Mean Shoot Length (cm) | |
|---|---|---|---|---|
| BA | Kin | |||
| 0.0 | 0.0 | 0.0 ± 0.0 f | 0.0 ± 0.0 e | 0.0 ± 0.0 g |
| 2.5 | - | 42.62 ± 0.93 d | 2.26 ± 0.41 d | 1.88 ± 0.066 ef |
| 5.0 | - | 58.68 ± 1.35 b | 4.68 ± 0.54 b | 2.28 ± 0.12 bd |
| 7.5 | - | 71.26 ± 0.66 a | 6.32 ± 0.38 a | 2.58 ± 0.11 a |
| 10.0 | - | 50.84 ± 1.07 c | 3.86 ± 0.57 bc | 2.41 ± 0.11 ab |
| - | 2.5 | 36.64 ± 1.29 e | 1.84 ± 0.38 d | 1.82 ± 0.06 f |
| - | 5.0 | 43.92 ± 1.36 d | 3.88 ± 0.39 bc | 2.04 ± 0.09 df |
| - | 7.5 | 59.28 ± 1.24 b | 4.52 ± 0.55 b | 2.34 ± 0.13 abc |
| - | 10.0 | 41.46 ± 0.75 d | 2.86 ± 0.39 cd | 2.12 ± 0.13 cde |
| NAA | IBA | Response (%) 1 | Mean No. of Shoots | Mean Shoot Length (cm) |
|---|---|---|---|---|
| 0.0 | 0.0 | 71.2 ± 0.66 e | 6.32 ± 0.38 c | 2.58 ± 0.11 cd |
| 0.1 | - | 73.22 ± 1.16 d | 6.94 ± 0.42 c | 2.42 ± 0.09 de |
| 0.5 | - | 79.68 ± 0.83 b | 9.94 ± 0.49 b | 2.78 ± 0.11 bc |
| 1.0 | - | 85.72 ± 0.78 a | 13.92 ± 0.41 a | 3.16 ± 0.08 a |
| 2.0 | - | 76.62 ± 0.51 c | 9.10 ± 0.46 b | 2.61 ± 0.09 cd |
| - | 0.1 | 70.46 ± 0.51 e | 6.52 ± 0.51 c | 2.18 ± 0.10 e |
| - | 0.5 | 75.28 ± 0.37 c | 7.46 ± 0.50 c | 2.56 ± 0.05 cd |
| - | 1.0 | 80.98 ± 0.56 b | 10.08 ± 0.55 b | 2.92 ± 0.09 ab |
| - | 2.0 | 75.42 ± 0.51 c | 8.91 ± 0.43 b | 2.44 ± 0.18 de |
| Auxins (μM) | Response (%) 1 | No. of Roots/Microshoots | Root Length (cm) | |
|---|---|---|---|---|
| IBA | NAA | |||
| 0.0 | 0.0 | 0.0 ± 0.0 f | 0.0 ± 0.0 d | 0.0 ± 0.0 e |
| 0.5 | - | 83.32 ± 0.88 c | 4.2 ± 0.37 bc | 2.18 ± 0.20 d |
| 1.0 | - | 94.19 ± 0.74 a | 6.8 ± 0.66 a | 4.34 ± 0.25 a |
| 2.0 | - | 85.83 ± 0.67 b | 4.4 ± 0.51 bc | 3.78 ± 0.20 b |
| - | 0.5 | 67.28 ± 1.07 e | 3.2 ± 0.374 c | 1.96 ± 0.14 d |
| - | 1.0 | 83.96 ± 0.56 bc | 4.8 ± 0.374 b | 2.88 ± 0.33 c |
| - | 2.0 | 76.73 ± 0.97 d | 3.8 ± 0.374 bc | 2.28 ± 0.14 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alatar, A.A.; Qahtan, A.A.; Abdel-Salam, E.M.; Faisal, M.; El-Sheikh, M.A. Development of an Efficient and Rapid Micropropagation Protocol for In Vitro Multiplication of Maerua crassifolia Forssk. Forests 2023, 14, 1160. https://doi.org/10.3390/f14061160
Alatar AA, Qahtan AA, Abdel-Salam EM, Faisal M, El-Sheikh MA. Development of an Efficient and Rapid Micropropagation Protocol for In Vitro Multiplication of Maerua crassifolia Forssk. Forests. 2023; 14(6):1160. https://doi.org/10.3390/f14061160
Chicago/Turabian StyleAlatar, Abdulrahman A., Ahmed A. Qahtan, Eslam M. Abdel-Salam, Mohammad Faisal, and Mohamed A. El-Sheikh. 2023. "Development of an Efficient and Rapid Micropropagation Protocol for In Vitro Multiplication of Maerua crassifolia Forssk" Forests 14, no. 6: 1160. https://doi.org/10.3390/f14061160
APA StyleAlatar, A. A., Qahtan, A. A., Abdel-Salam, E. M., Faisal, M., & El-Sheikh, M. A. (2023). Development of an Efficient and Rapid Micropropagation Protocol for In Vitro Multiplication of Maerua crassifolia Forssk. Forests, 14(6), 1160. https://doi.org/10.3390/f14061160