

Cryopreservation and Cryotolerance Mechanism in Zygotic Embryo and Embryogenic Callus of Oil Palm


 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Cryopreservation Methods
2.3. Tissue Sectioning and Microscopy
2.4. Detection of H2O2 Content and Antioxidant Enzyme Activities
2.5. Genetic Stability Detection
2.6. Transcriptome Analysis
2.7. qPCR Validation
2.8. Statistical Analysis
3. Results
3.1. Effect of Dehydration and Cryopreservation Time on Viability and Germination of ZE
3.2. Effect of Pre-Culture and PVS2 Treatments on the Viability Rate of EC
3.3. Analysis of Cell Viability, H2O2 Content, and Antioxidants Activity
3.4. Genetic Stability of Cryopreserved Material
3.5. Transcriptome Changes
3.6. Response of DEGs in Cryotolerance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khawnium, T.; Te-Chato, S. Simple vitrification protocol for cryopreservation of oil palm using embryogenic culture. J. Agric. Technol. 2011, 7, 519–529. [Google Scholar]
- Corley, R.H.V.; Tinker, P.B. The Oil Palm; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Suranthran, P.; Gantait, S.; Sinniah, U.R.; Subramaniam, S.; Alwee, S.S.R.S.; Roowi, S.H. Effect of loading and vitrification solutions on survival of cryopreserved oil palm polyembryoids. Plant Growth Regul. 2012, 66, 101–109. [Google Scholar] [CrossRef]
- Palanyandy, S.R.; Suranthran, P.; Gantait, S.; Sinniah, U.R.; Subramaniam, S.; Abdul Aziz, M.; Syed Alwee, S.S.R.; Roowi, S.H. In vitro developmental study of oil palm (Elaeis guineensis Jacq.) polyembryoids from cell suspension using scanning electron microscopy. Acta Physiol. Plant. 2013, 35, 1727–1733. [Google Scholar] [CrossRef]
- Weckx, S.; Inzé, D.; Maene, L. Tissue culture of oil palm: Finding the balance between mass propagation and somaclonal variation. Front. Plant Sci. 2019, 10, 722. [Google Scholar] [CrossRef] [PubMed]
- Scherwinski-Pereira, J.E.; da Guedes, R.S.; Fermino, P.C.P.; Silva, T.L.; Costa, F.H.S. Somatic embryogenesis and plant regeneration in oil palm using the thin cell layer technique. Vitr. Cell. Dev. Biol. Plant 2010, 46, 378–385. [Google Scholar] [CrossRef]
- Kong, E.Y.; Biddle, J.; Foale, M.; Adkins, S.W. Cell suspension culture: A potential in vitro culture method for clonal propagation of coconut plantlets via somatic embryogenesis. Ind. Crops Prod. 2020, 147, 112125. [Google Scholar] [CrossRef]
- Kushairi, A.; Tarmizi, A.; Zamzuri, I.; Ong-Abdullah, M.; Samsul Kamal, R.; Ooi, S.; Rajanaidu, N. Production, performance and advances in oil palm tissue culture. In Proceedings of the International Seminar on Advances in Oil Palm Tissue Culture, Yogyakarta, Indonesia, 29 May 2010. [Google Scholar]
- Beulé, T.; Ilbert, P.; Adeoti, K.; Durand-Gasselin, T.; Dumet, D.; Engelmann, F.; Morcillo, F. Recovery of oil palm (Elaeis guineensis Jacq.) somatic embryos cryostored for 20 years. CryoLetters 2018, 39, 60–66. [Google Scholar]
- Corley, R.; Lee, C.; Law, I.; Wong, C. Abnormal flower development in oil palm clones. Planter 1986, 62, 233–240. [Google Scholar]
- Khan, F.S.; Goher, F.; Zhang, D.; Shi, P.; Li, Z.; Htwe, Y.M.; Wang, Y. Is CRISPR/Cas9 a way forward to fast-track genetic improvement in commercial palms? Prospects and limits. Front. Plant Sci. 2022, 13, 1042828. [Google Scholar] [CrossRef]
- Steinmacher, D.A.; Saldanha, C.W.; Clement, C.R.; Guerra, M.P. Cryopreservation of peach palm zygotic embryos. CryoLetters 2007, 28, 13–22. [Google Scholar]
- Norziha, A.; Marhalil, M.; Fadila, A.; Zulkifli, Y.; Maizura, I.; Mohd Din, A.; Rajanaidu, N.; Kushairi, A. Long-term storage of oil palm germplasm zygotic embryo using cryopreservation. J. Oil Palm Res. 2017, 29, 541–547. [Google Scholar]
- Rival, A.; Turquay, P.; Samosir, Y.; Adkins, S.W. Cryopreservation of coconut (Cocos nucifera L.) zygotic embryos does not induce morphological, cytological or molecular changes in recovered seedlings. Planta 2010, 232, 435–447. [Google Scholar]
- Engelmann, F.; Chabrillange, N.; Dussert, S.; Duval, Y. Cryopreservation of zygotic embryos and kernels of oil palm (Elaeis guineensis Jacq.). Seed Sci. Res. 1995, 5, 81–86. [Google Scholar] [CrossRef]
- da Silva, J.A.T.; Engelmann, F. Cryopreservation of oil palm (Elaeis guineensis Jacq.). Cryobiology 2017, 77, 82–88. [Google Scholar] [CrossRef]
- Ahmad Malike, F.; Amiruddin, M.D.; Yaakub, Z.; Marjuni, M.; Abdullah, N.; Abu Bakar, N.A.; Mustaffa, S.; Mohamad, M.M.; Hassan, M.Y.; Abdullah, M.O. Oil palm (Elaeis spp.) breeding in Malaysia. Adv. Plant Breed. Strateg. Ind. Food Crops 2019, 6, 489–535. [Google Scholar]
- Alansi, S.; Al-Qurainy, F.; Nadeem, M.; Khan, S.; Tarroum, M.; Alshameri, A.; Gaafar, A.-R.Z. Cryopreservation: A tool to conserve date palm in Saudi Arabia. Saudi J. Biol. Sci. 2019, 26, 1896–1902. [Google Scholar] [CrossRef] [PubMed]
- Araújo de Oliveira, A.C.; da Silva Lédo, A.; Polek, M.; Krueger, R.; Shepherd, A.; Volk, G.M. Optimization of in vitro germination and cryopreservation conditions for preserving date palm pollen in the USDA National Plant Germplasm System. Plant Cell Tissue Organ Cult. 2021, 144, 223–232. [Google Scholar] [CrossRef]
- Lédo, A.S.; Vendrame, W.A. Coconut micropropagation and cryopreservation. Horticultural Reviews; Wiley-Blackwell: New York, NY, USA, 2021; Volume 48, pp. 307–337. [Google Scholar]
- Bekheet, S.A.; Sota, V.; El-Shabrawi, H.M.; El-Minisy, A.M. Cryopreservation of shoot apices and callus cultures of globe artichoke using vitrification method. J. Genet. Eng. Biotechnol. 2020, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kaviani, B. Conservation of plant genetic resources by cryopreservation. Aust. J. Crop Sci. 2011, 5, 778–800. [Google Scholar]
- Sakai, A.; Engelmann, F. Vitrification, encapsulation-vitrification and droplet-vitrification: A review. CryoLetters 2007, 28, 151–172. [Google Scholar]
- de Freitas, R.T.; Paiva, R.; Sales, T.S.; da Silva, D.P.C.; de Souza, A.C. Cryopreservation of Coffea arabica L. zygotic embryos by vitrification. Not. Bot. Horti Agrobot. Cluj Napoca 2016, 44, 445–451. [Google Scholar] [CrossRef][Green Version]
- Chen, G.-Q.; Ren, L.; Zhang, J.; Reed, B.M.; Zhang, D.; Shen, X.-H. Cryopreservation affects ROS-induced oxidative stress and antioxidant response in Arabidopsis seedlings. Cryobiology 2015, 70, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Quijada-Rivera, M.; Tiznado-Hernández, M.E.; Hernández-Oñate, M.Á.; Vargas-Arispuro, I.; Astorga-Cienfuegos, K.R.; Lazo-Javalera, M.F.; Rivera-Domínguez, M. Transcriptome assessment in’Red Globe’grapevine zygotic embryos during the cooling and warming phase of the cryopreservation procedure. Cryobiology 2022, 110, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Htwe, C.S.S.; Rajkumar, S.; Pathania, P.; Agrawal, A. Transcriptome Profiling during Sequential Stages of Cryopreservation in Banana (Musa AAA cv Borjahaji) Shoot Meristem. Plants 2023, 12, 1165. [Google Scholar] [CrossRef]
- Huang, B.; Zhang, J.-M.; Chen, X.-L.; Xin, X.; Yin, G.-K.; He, J.-J.; Lu, X.-X.; Zhou, Y.-C. Oxidative damage and antioxidative indicators in 48 h germinated rice embryos during the vitrification-cryopreservation procedure. Plant Cell Rep. 2018, 37, 1325–1342. [Google Scholar] [CrossRef]
- Hao, Y.-J.; You, C.-X.; Deng, X.-X. Analysis of ploidy and the patterns of amplified fragment length polymorphism and methylation sensitive amplified polymorphism in strawberry plants recovered from cryopreservation. CryoLetters 2002, 23, 37–46. [Google Scholar] [PubMed]
- Gantait, S.; Sinniah, U.R.; Suranthran, P.; Palanyandy, S.R.; Subramaniam, S. Improved cryopreservation of oil palm (Elaeis guineensis Jacq.) polyembryoids using droplet vitrification approach and assessment of genetic fidelity. Protoplasma 2015, 252, 89–101. [Google Scholar] [CrossRef]
- Das Bhowmik, S.S.; Basu, A.; Sahoo, L. Direct shoot organogenesis from rhizomes of medicinal Zingiber Alpinia calcarata Rosc. and evaluation of genetic stability by RAPD and ISSR markers. J. Crop Sci. Biotechnol. Adv. 2016, 19, 157–165. [Google Scholar] [CrossRef]
- Rao, A.A.; Chaudhury, R.; Kumar, S.; Velu, D.; Saraswat, R.; Kamble, C. Cryopreservation of mulberry germplasm core collection and assessment of genetic stability through ISSR markers. Int. J. Ind. Ent. 2007, 15, 23–33. [Google Scholar]
- Leunufna, S.; Keller, E. Cryopreservation of yams using vitrification modified by including droplet method: Effects of cold acclimation and sucrose. CryoLetters 2005, 26, 93–102. [Google Scholar]
- Liu, Y.-G.; Liu, L.-X.; Wang, L.; Gao, A.-Y. Determination of genetic stability in surviving apple shoots following cryopreservation by vitrification. CryoLetters 2008, 29, 7–14. [Google Scholar] [PubMed]
- Villalobos-Olivera, A.; Ferreira, C.F.; Yanes-Paz, E.; Lorente, G.Y.; Souza, F.V.; Engelmann, F.; Martínez-Montero, M.E.; Lorenzo, J.C. Inter simple sequence repeat (ISSR) markers reveal DNA stability in pineapple plantlets after shoot tip cryopreservation. Vegetos 2022, 35, 360–366. [Google Scholar] [CrossRef]
- Espasandin, F.D.; Brugnoli, E.A.; Ayala, P.G.; Ayala, L.P.; Ruiz, O.A.; Sansberro, P.A. Long-term preservation of Lotus tenuis adventitious buds. Plant Cell Tissue Organ Cult. 2019, 136, 373–382. [Google Scholar] [CrossRef]
- Teixeira, J.; Söndahl, M.; Kirby, E. Somatic embryogenesis from immature inflorescences of oil palm. Plant Cell Rep. 1994, 13, 247–250. [Google Scholar] [CrossRef]
- Eeuwens, C. Mineral requirements for growth and callus initiation of tissue explants excised from mature coconut palms (Cocos nucifera) and cultured in vitro. Physiol. Plant. 1976, 36, 23–28. [Google Scholar] [CrossRef]
- Zhang, D.; Shi, P.; Htwe, Y.M.; Li, Z.; Ihase, L.O.; Mason, A.S.; Sun, X.; Xiao, Y.; Wang, Y. Caffeate may play an important role in the somatic embryogenesis of oil palm (Elaeis guineensis Jacq.). Ind. Crops Prod. 2021, 174, 114143. [Google Scholar] [CrossRef]
- Copeland, L.O.; McDonald, M.B.; Copeland, L.O.; McDonald, M.B. Seed viability and viability testing. In Principles of Seed Science and Technology; Springer: New York, NY, USA, 2001; pp. 124–139. [Google Scholar]
- Kiernan, J. Histological and Histochemical Methods; Scion Publishing Ltd.: Banbury, UK, 2015. [Google Scholar]
- Ting, N.-C.; Jansen, J.; Mayes, S.; Massawe, F.; Sambanthamurthi, R.; Ooi, L.C.-L.; Chin, C.W.; Arulandoo, X.; Seng, T.-Y.; Alwee, S.S.R.S. High density SNP and SSR-based genetic maps of two independent oil palm hybrids. BMC Genom. 2014, 15, 309. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, D.; Shi, P.; Htwe, Y.M.; Yu, Q.; Huang, L.; Zhou, H.; Liu, L.; Wang, Y. Cell wall lignification may be necessary for somatic embryogenesis of areca palm (Areca catechu). Sci. Hort. 2023, 307, 111538. [Google Scholar] [CrossRef]
- Team, R. RStudio: Integrated Development for R; RStudio: Boston, MA, USA, 2015. [Google Scholar]
- Team R.D.C. A Language and Environment for Statistical Computing. 2009. Available online: http://www.R-project.org (accessed on 31 March 2023).
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Folgado, R.; Sergeant, K.; Renaut, J.; Swennen, R.; Hausman, J.-F.; Panis, B. Changes in sugar content and proteome of potato in response to cold and dehydration stress and their implications for cryopreservation. J. Proteom. 2014, 98, 99–111. [Google Scholar] [CrossRef]
- Baust, J.M.; Campbell, L.H.; Harbell, J.W. Best practices for cryopreserving, thawing, recovering, and assessing cells. Vitr. Cell. Dev. Biol. Anim. 2017, 53, 855–871. [Google Scholar] [CrossRef] [PubMed]
- Kaviani, B.; Kulus, D. Cryopreservation of endangered ornamental plants and fruit crops from tropical and subtropical regions. Biology 2022, 11, 847. [Google Scholar] [CrossRef]
- Barraco, G.; Sylvestre, I.; Engelmann, F. Cryopreservation of sugarcane (Saccharum spp.) shoot tips using encapsulation and dropletvitrification. Sci. Hortic. 2011, 130, 320–324. [Google Scholar] [CrossRef]
- Khawniam, T.; Te-Chato, S. Cryopreservation of embryogenic callus of hybrid tenera oil palm by dehydration technique and evaluation of somaclonal variation by SSR marker. J. Agric. Technol. 2012, 8, 2115–2125. [Google Scholar]
- Fang, J.-Y.; Wetten, A.; Hadley, P. Cryopreservation of cocoa (Theobroma cacao L.) somatic embryos for long-term germplasm storage. Plant Sci. 2004, 166, 669–675. [Google Scholar] [CrossRef]
- Fki, L.; Bouaziz, N.; Chkir, O.; Benjemaa-Masmoudi, R.; Rival, A.; Swennen, R.; Drira, N.; Panis, B. Cold hardening and sucrose treatment improve cryopreservation of date palm meristems. Biol. Plant. 2013, 57, 375–379. [Google Scholar] [CrossRef]
- Lynch, P.T.; Siddika, A.; Johnston, J.W.; Trigwell, S.M.; Mehra, A.; Benelli, C.; Lambardi, M.; Benson, E.E. Effects of osmotic pretreatments on oxidative stress, antioxidant profiles and cryopreservation of olive somatic embryos. Plant Sci. 2011, 181, 47–56. [Google Scholar] [CrossRef]
- Niino, T.; Sakai, A.; Yakuwa, H.; Nojiri, K. Cryopreservation of in vitro-grown shoot tips of apple and pear by vitrification. Plant Cell Tissue Organ Cult. 1992, 28, 261–266. [Google Scholar] [CrossRef]
- Dumet, D.; Engelmann, F.; Chabrillange, N.; Duval, Y. Cryopreservation of oil palm (Elaeis guineensis Jacq.) somatic embryos involving a desiccation step. Plant Cell Rep. 1993, 12, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Cai, C.; Wang, R.; Song, S.; Song, J. Cytological and physiological changes in recalcitrant Chinese fan palm (Livistona chinensis) embryos during cryopreservation. Protoplasma 2012, 249, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Zhang, D.; Jiang, X.-N.; Gai, Y.; Wang, W.-M.; Reed, B.M.; Shen, X.-H. Peroxidation due to cryoprotectant treatment is a vital factor for cell survival in Arabidopsis cryopreservation. Plant Sci. 2013, 212, 37–47. [Google Scholar] [CrossRef]
- Ren, L.; Wang, M.-R.; Wang, Q.-C. ROS-induced oxidative stress in plant cryopreservation: Occurrence and alleviation. Planta 2021, 254, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Ren, L.; Chen, G.-Q.; Zhang, J.; Reed, B.M.; Shen, X.-H. ROS-induced oxidative stress and apoptosis-like event directly affect the cell viability of cryopreserved embryogenic callus in Agapanthus praecox. Plant Cell Rep. 2015, 34, 1499–1513. [Google Scholar] [CrossRef] [PubMed]
- Ren, R.; Li, Z.; Zhang, L.; Zhou, H.; Jiang, X.; Liu, Y. Enzymatic and nonenzymatic antioxidant systems impact the viability of cryopreserved Paeonia suffruticosa pollen. Plant Cell Tissue Organ Cult. 2021, 144, 233–246. [Google Scholar] [CrossRef]
- Agrawal, A.; Tyagi, R.; Goswami, R.; Uma, S.; Saraswathi, M.; Durai, P. Cryobanking of banana (Musa sp.) germplasm in India: Evaluation of agronomic and molecular traits of cryopreserved plants. In Proceedings of the I International Symposium on Cryopreservation in Horticultural Species, Leuven, Belgium, 5–8 April 2009; Volume 908, pp. 129–138. [Google Scholar]
- Benson, E.E.; Betsou, F.; Fuller, B.J.; Harding, K.; Kofanova, O. Translating cryobiology principles into trans-disciplinary storage guidelines for biorepositories and biobanks: A concept paper. CryoLetters 2013, 34, 277–312. [Google Scholar] [PubMed]
- Li, B.-Q.; Feng, C.-H.; Wang, M.-R.; Hu, L.-Y.; Volk, G.; Wang, Q.-C. Recovery patterns, histological observations and genetic integrity in Malus shoot tips cryopreserved using droplet-vitrification and encapsulation-dehydration procedures. J. Biotechnol. 2015, 214, 182–191. [Google Scholar] [CrossRef]
- Wang, M.-R.; Bi, W.; Shukla, M.R.; Ren, L.; Hamborg, Z.; Blystad, D.-R.; Saxena, P.K.; Wang, Q.-C. Epigenetic and genetic integrity, metabolic stability, and field performance of cryopreserved plants. Plants 2021, 10, 1889. [Google Scholar] [CrossRef]
- Bakoumé, C.; Wickneswari, R.; Siju, S.; Rajanaidu, N.; Kushairi, A.; Billotte, N. Genetic diversity of the world’s largest oil palm (Elaeis guineensis Jacq.) field genebank accessions using microsatellite markers. Genet. Resour. Crop Evol. 2015, 62, 349–360. [Google Scholar] [CrossRef]
- Khan, F.S.; Gan, Z.-M.; Li, E.-Q.; Ren, M.-K.; Hu, C.-G.; Zhang, J.-Z. Transcriptomic and physiological analysis reveals interplay between salicylic acid and drought stress in citrus tree floral initiation. Planta 2022, 255, 24. [Google Scholar] [CrossRef]
- Gadjev, I.; Vanderauwera, S.; Gechev, T.S.; Laloi, C.; Minkov, I.N.; Shulaev, V.; Apel, K.; Inzé, D.; Mittler, R.; Van Breusegem, F. Transcriptomic footprints disclose specificity of reactive oxygen species signaling in Arabidopsis. Plant Physiol. 2006, 141, 436–445. [Google Scholar] [CrossRef]
- Ren, L.; Zhang, D.; Chen, G.-Q.; Reed, B.M.; Shen, X.-H.; Chen, H.-Y. Transcriptomic profiling revealed the regulatory mechanism of Arabidopsis seedlings response to oxidative stress from cryopreservation. Plant Cell Rep. 2015, 34, 2161–2178. [Google Scholar] [CrossRef] [PubMed]
- Benson, E.E. Cryopreservation of phytodiversity: A critical appraisal of theory & practice. Crit. Rev. Plant Sci. 2008, 27, 141–219. [Google Scholar]
- Guo, X.; Liu, D.; Chong, K. Cold signaling in plants: Insights into mechanisms and regulation. J. Integr. Plant Biol. 2018, 60, 745–756. [Google Scholar] [CrossRef]
- Jasso-Robles, F.I.; Gonzalez, M.E.; Pieckenstain, F.L.; Ramírez-García, J.M.; de la Luz Guerrero-González, M.; Jiménez-Bremont, J.F.; Rodríguez-Kessler, M. Decrease of Arabidopsis PAO activity entails increased RBOH activity, ROS content and altered responses to Pseudomonas. Plant Sci. 2020, 292, 110372. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
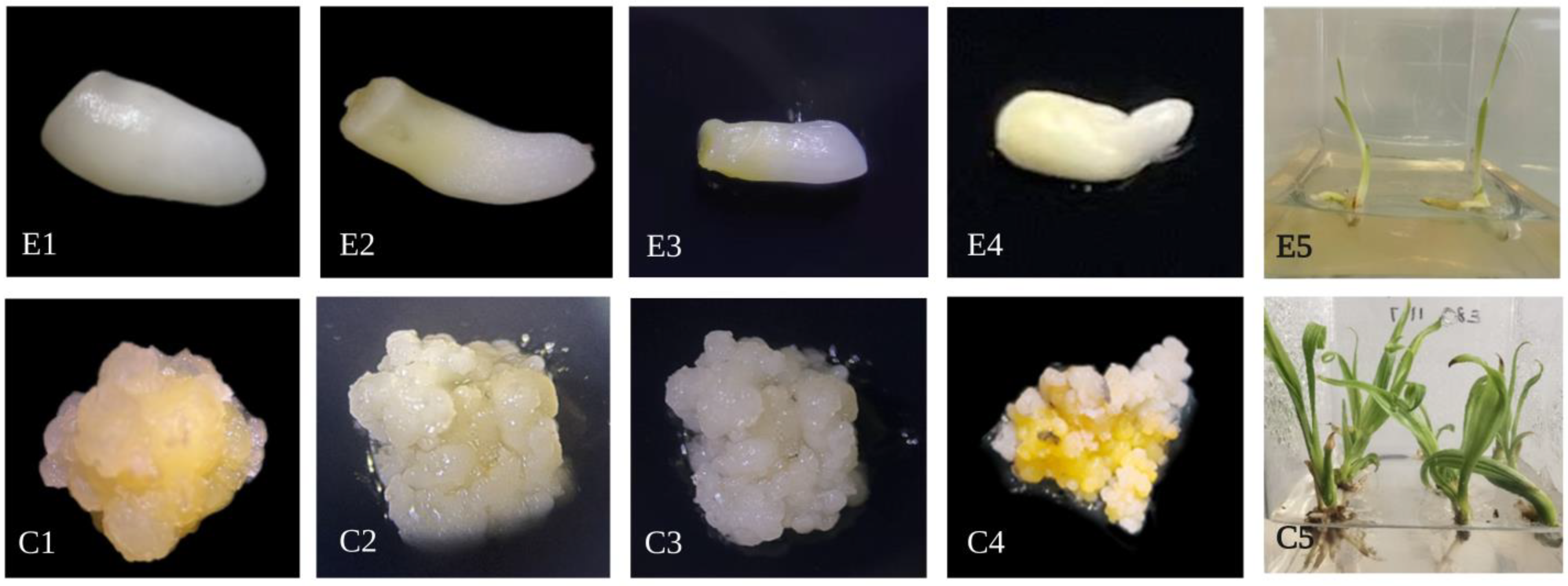{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
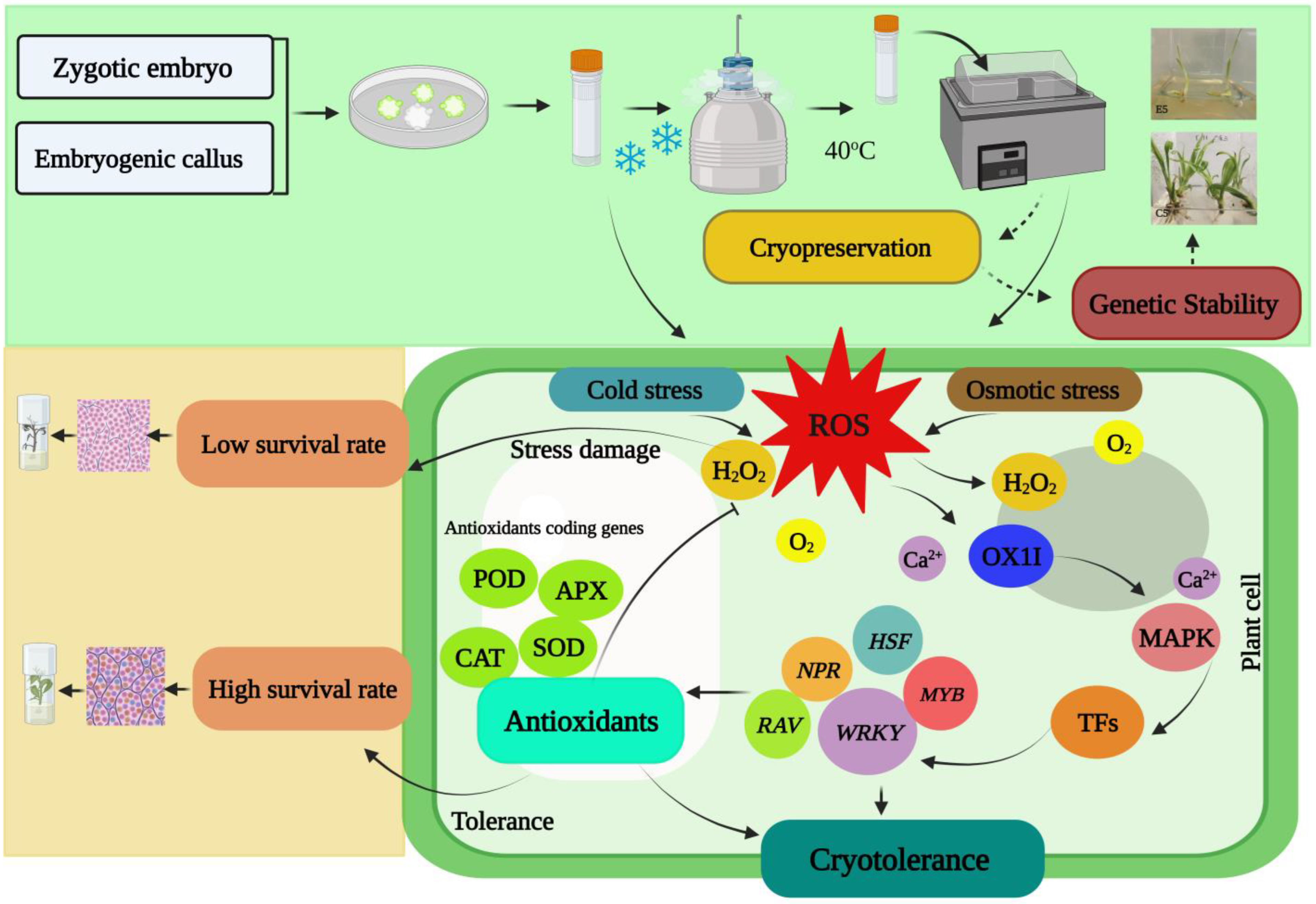{kind=link}
| Dehydrating Time (d) | Viability (%) | Germination (%) |
|---|---|---|
| 0 | 0 | 0 |
| 3 | 96.67 ± 0.03 a | 90.88 ± 0.03 a |
| 4 | 80.00 ± 0.03 b | 69.58 ± 0.04 b |
| 10 | 0 | 0 |
| Cryopreservation Time (d) | Viability (%) | Germination (%) |
|---|---|---|
| 1 | 91.11 ± 0.02 a | 85.56 ± 0.03 a |
| 3 | 90.00 ± 0.03 a | 83.33 ± 0.05 a |
| 7 | 91.11 ± 0.04 a | 83.33 ± 0.03 a |
| 20 | 90.00 ± 0.03 a | 81.11 ± 0.04 a |
| 40 | 90.00 ± 0.03 a | 82.22 ± 0.06 a |
| 60 | 92.22 ± 0.03 a | 84.44 ± 0.02 a |
| +LN/ −LN | PVS2 Treatment Time (min) | Sucrose Concentration (M) | |||
|---|---|---|---|---|---|
| 0.3 | 0.5 | 0.7 | 0.9 | ||
| +LN | 5 | 68.33 ± 0.02 a | 38.33 ± 0.12 | 55.00 ± 0.12 | 25.00 ± 0.04 |
| −LN | 80.53 ± 0.11 | 54.57 ± 0.21 | 73.57 ± 0.05 | 50.54 ± 0.10 | |
| +LN | 10 | 63.33 ± 0.10 a | 43.33 ± 0.08 | 46.67 ± 0.06 | 23.33 ± 0.06 |
| −LN | 77.54 ± 0.14 | 58.57 ± 0.08 | 61.39 ± 0.08 | 50.61 ± 0.03 | |
| +LN | 30 | 46.67 ± 0.05 | 48.33 ± 0.06 | 58.33 ± 0.16 | 21.67 ± 0.06 |
| −LN | 64.48 ± 0.09 | 66.40 ± 0.04 | 68.51 ± 0.12 | 46.38 ± 0.14 | |
| +LN | 60 | 25.00 ± 0.04 | 76.67 ± 0.05 a | 55.00 ± 0.16 | 6.67 ± 0.02 |
| −LN | 53.48 ± 0.12 | 85.56 ± 0.14 | 74.49 ± 0.1 | 20.52 ± 0.12 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, Q.; Shi, P.; Khan, F.S.; Htwe, Y.M.; Zhang, D.; Li, Z.; Wei, X.; Yu, Q.; Zhou, K.; Wang, Y. Cryopreservation and Cryotolerance Mechanism in Zygotic Embryo and Embryogenic Callus of Oil Palm. Forests 2023, 14, 966. https://doi.org/10.3390/f14050966
Wei Q, Shi P, Khan FS, Htwe YM, Zhang D, Li Z, Wei X, Yu Q, Zhou K, Wang Y. Cryopreservation and Cryotolerance Mechanism in Zygotic Embryo and Embryogenic Callus of Oil Palm. Forests. 2023; 14(5):966. https://doi.org/10.3390/f14050966
Chicago/Turabian StyleWei, Qiuen, Peng Shi, Faiza Shafique Khan, Yin Min Htwe, Dapeng Zhang, Zhiying Li, Xueke Wei, Qun Yu, Kaibing Zhou, and Yong Wang. 2023. "Cryopreservation and Cryotolerance Mechanism in Zygotic Embryo and Embryogenic Callus of Oil Palm" Forests 14, no. 5: 966. https://doi.org/10.3390/f14050966
APA StyleWei, Q., Shi, P., Khan, F. S., Htwe, Y. M., Zhang, D., Li, Z., Wei, X., Yu, Q., Zhou, K., & Wang, Y. (2023). Cryopreservation and Cryotolerance Mechanism in Zygotic Embryo and Embryogenic Callus of Oil Palm. Forests, 14(5), 966. https://doi.org/10.3390/f14050966