
Forest Tent Caterpillar Outbreaks Drive Change in Ant Communities in Boreal Forests

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Experimental Design
2.2. Experimental Design
2.3. Environmental Variables
2.4. Ant Survey
2.5. Survey of Other Arthropods
2.6. Statistical Analysis
3. Results
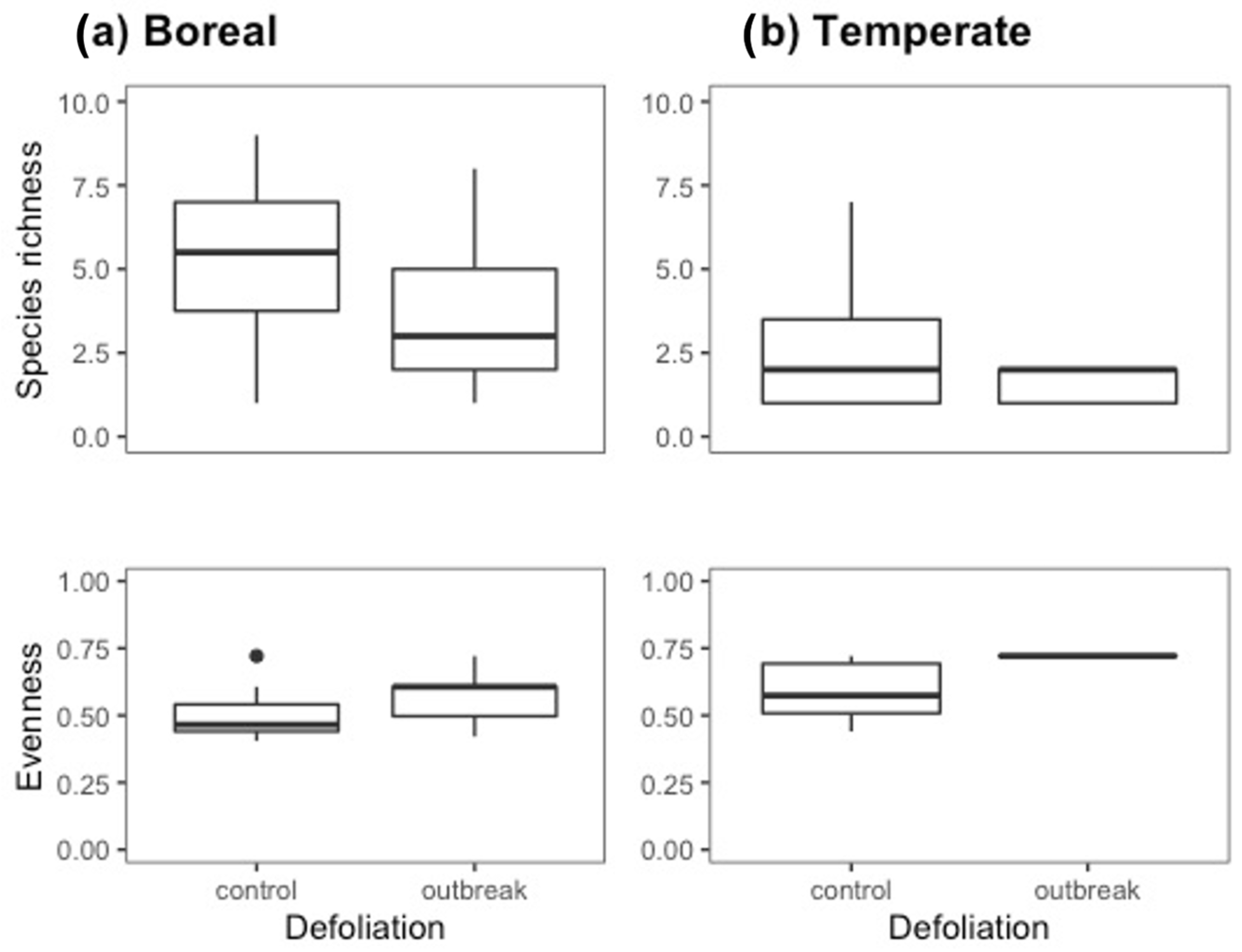
3.1. Species Richness and Evenness
3.2. Species Composition
Ordination
3.3. Beta Diversity
3.4. Indicator Species
4. Discussion
4.1. Ant Community Responses to Disturbance
4.2. Indicator Species
4.3. Prey, Predators, and Outbreaks
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aakala, T.; Remy, C.C.; Arseneault, D.; Morin, H.; Girardin, M.P.; Gennaretti, F.; Navarro, L.; Kuosmanen, N.; Ali, A.A.; Boucher, É.; et al. Millennial-scale disturbance history of the boreal zone. In Boreal Forests in the Face of Climate Change: Sustainable Management; Girona, M.M., Morin, H., Gauthier, S., Bergeron, Y., Eds.; Advances in Global Change Research; Springer International Publishing: Cham, Switzerland, 2023; pp. 53–87. ISBN 978-3-031-15988-6. [Google Scholar]
- Montoro Girona, M.; Navarro, L.; Morin, H. A Secret Hidden in the Sediments: Lepidoptera Scales. Front. Ecol. Evol. 2018, 6, 2. [Google Scholar] [CrossRef]
- Navarro, L.; Morin, H.; Bergeron, Y.; Girona, M.M. Changes in spatiotemporal patterns of 20th century spruce budworm outbreaks in eastern Canadian boreal forests. Front. Plant Sci. 2018, 9, 1905. [Google Scholar] [CrossRef]
- Lavoie, J.; Montoro Girona, M.; Grosbois, G.; Morin, H. Does the type of silvicultural practice influence spruce budworm defoliation of seedlings? Ecosphere 2021, 12, e03506. [Google Scholar] [CrossRef]
- De Grandpré, L.; Marchand, M.; Kneeshaw, D.D.; Paré, D.; Boucher, D.; Bourassa, S.; Gervais, D.; Simard, M.; Griffin, J.M.; Pureswaran, D.S. Defoliation-induced changes in foliage quality may trigger broad-scale insect outbreaks. Commun. Biol. 2022, 5, 463. [Google Scholar] [CrossRef] [PubMed]
- Caron, A.; Jarry, J.J.; Despland, E. Early instar mortality of a forest pest caterpillar: Which mortality sources increase during an outbreak crash? Entomol. Exp. Appl. 2022, 170, 268–276. [Google Scholar] [CrossRef]
- Swank, W.T.; Waide, J.B.; Crossley, D.A.; Todd, R.L. Insect defoliation enhances nitrate export from forest ecosystems. Oecologia 1981, 51, 297–299. [Google Scholar] [CrossRef]
- Hunter, M.D. Insect population dynamics meets ecosystem ecology: Effects of herbivory on soil nutrient dynamics. Agric. For. Entomol. 2001, 3, 77–84. [Google Scholar] [CrossRef]
- Metcalfe, D.B.; Asner, G.P.; Martin, R.E.; Silva Espejo, J.E.; Huasco, W.H.; Farfán Amézquita, F.F.; Carranza-Jimenez, L.; Galiano Cabrera, D.F.; Baca, L.D.; Sinca, F.; et al. Herbivory makes major contributions to ecosystem carbon and nutrient cycling in tropical forests. Ecol. Lett. 2014, 17, 324–332. [Google Scholar] [CrossRef]
- Metcalfe, D.B.; Crutsinger, G.M.; Kumordzi, B.B.; Wardle, D.A. Nutrient fluxes from insect herbivory increase during ecosystem retrogression in boreal forest. Ecology 2016, 97, 124–132. [Google Scholar] [CrossRef]
- Lovett, G.M.; Christenson, L.M.; Groffman, P.M.; Jones, C.G.; Hart, J.E.; Mitchell, M.J. Insect Defoliation and Nitrogen Cycling in Forests. BioScience 2002, 52, 335. [Google Scholar] [CrossRef]
- Gravel, D.; Albouy, C.; Thuiller, W. The meaning of functional trait composition of food webs for ecosystem functioning. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150268. [Google Scholar] [CrossRef]
- Barnes, A.D.; Jochum, M.; Lefcheck, J.S.; Eisenhauer, N.; Scherber, C.; O’Connor, M.I.; de Ruiter, P.; Brose, U. Energy flux: The link between multitrophic biodiversity and ecosystem functioning. Trends Ecol. Evol. 2018, 33, 186–197. [Google Scholar] [CrossRef]
- Kristensen, J.A.; Metcalfe, D.B.; Rousk, J. The biogeochemical consequences of litter transformation by insect herbivory in the Subarctic: A microcosm simulation experiment. Biogeochemistry 2018, 138, 323–336. [Google Scholar] [CrossRef]
- Lovett, G.M.; Ruesink, A.E. Carbon and nitrogen mineralization from decomposing gypsy moth frass. Oecologia 1995, 104, 133–138. [Google Scholar] [CrossRef]
- Debaly, Z.M.; Marchand, P.; Girona, M.M. Autoregressive models for time series of random sums of positive variables: Application to tree growth as a function of climate and insect outbreak. Ecol. Model. 2022, 471, 110053. [Google Scholar] [CrossRef]
- Odum, E.P. The Strategy of Ecosystem Development. Ekistics 1970, 29, 234–238. [Google Scholar]
- Calderón-Sanou, I.; Münkemüller, T.; Zinger, L.; Schimann, H.; Yoccoz, N.G.; Gielly, L.; Foulquier, A.; Hedde, M.; Ohlmann, M.; Roy, M.; et al. Cascading effects of moth outbreaks on subarctic soil food webs. Sci. Rep. 2021, 11, 15054. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Caruso, T. Soil microbial community responses to climate extremes: Resistance, resilience and transitions to alternative states. Philos. Trans. R. Soc. B 2020, 375, 20190112. [Google Scholar] [CrossRef]
- De Ruiter, P.C.; Neutel, A.-M.; Moore, J.C. Energetics, patterns of interaction strengths, and stability in real ecosystems. Science 1995, 269, 1257–1260. [Google Scholar] [CrossRef] [PubMed]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.E.; Holt, R.D.; Jackson, J.B. Trophic downgrading of planet Earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef]
- Saravesi, K.; Aikio, S.; Wäli, P.R.; Ruotsalainen, A.L.; Kaukonen, M.; Huusko, K.; Suokas, M.; Brown, S.P.; Jumpponen, A.; Tuomi, J. Moth outbreaks alter root-associated fungal communities in subarctic mountain birch forests. Microb. Ecol. 2015, 69, 788–797. [Google Scholar] [CrossRef]
- Vindstad, O.P.L.; Schultze, S.; Jepsen, J.U.; Biuw, E.M.; Kapari, L.T.; Sverdrup-Thygeson, A.; Ims, R.A. Numerical responses of saproxylic beetles to rapid increases in dead wood availability following geometrid moth outbreaks in sub-arctic mountain birch forest. PLoS ONE 2014, 9, e99624. [Google Scholar] [CrossRef]
- Sandén, H.; Mayer, M.; Stark, S.; Sandén, T.; Nilsson, L.O.; Jepsen, J.U.; Wäli, P.R.; Rewald, B. Moth outbreaks reduce decomposition in subarctic forest soils. Ecosystems 2020, 23, 151–163. [Google Scholar] [CrossRef]
- Hoffmann, B.D.; Andersen, A.N. Responses of ants to disturbance in Australia, with particular reference to functional groups. Austral Ecol. 2003, 28, 444–464. [Google Scholar] [CrossRef]
- Jurgensen, M.F.; Finér, L.; Domisch, T.; Kilpeläinen, J.; Punttila, P.; Ohashi, M.; Niemelä, P.; Sundström, L.; Neuvonen, S.; Risch, A.C. Organic mound-building ants: Their impact on soil properties in temperate and boreal forests. J. Appl. Entomol. 2008, 132, 266–275. [Google Scholar] [CrossRef]
- Risch, A.C.; Jurgensen, M.F. Ants in the soil system-a hydrological, chemical and biological approach. J. Appl. Entomol. 2008, 132, 265. [Google Scholar] [CrossRef]
- Domisch, T.; Finér, L.; Neuvonen, S.; Niemelä, P.; Risch, A.C.; Kilpeläinen, J.; Ohashi, M.; Jurgensen, M.F. Foraging activity and dietary spectrum of wood ants (Formica rufa group) and their role in nutrient fluxes in boreal forests. Ecol. Entomol. 2009, 34, 369–377. [Google Scholar] [CrossRef]
- Del Toro, I.; Ribbons, R.R.; Pelini, S.L. The little things that run the world revisited: A review of ant-mediated ecosystem services and disservices (Hymenoptera: Formicidae). Myrmecol. News 2012, 17, 133–146. [Google Scholar]
- Folgarait, P.J. Ant biodiversity and its relationship to ecosystem functioning: A review. Biodivers. Conserv. 1998, 7, 1221–1244. [Google Scholar] [CrossRef]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Wardle, D.A.; Hyodo, F.; Bardgett, R.D.; Yeates, G.W.; Nilsson, M.-C. Long-term aboveground and belowground consequences of red wood ant exclusion in boreal forest. Ecology 2011, 92, 645–656. [Google Scholar] [CrossRef]
- Laine, K.J.; Niemelä, P. The influence of ants on the survival of mountain birches during an Oporinia autumnata (Lep., Geometridae) outbreak. Oecologia 1980, 47, 39–42. [Google Scholar] [CrossRef]
- Punttila, P.; Niemelä, P.; Karhu, K. The impact of wood ants (Hymenoptera: Formicidae) on the structure of invertebrate community on mountain birch (Betula pubescens ssp. czerepanovii). In Proceedings of the Annales Zoologici Fennici; JSTOR. Finnish Zoological and Botanical Publishing Board: Helsinki, Finland, 2004; pp. 429–446. [Google Scholar]
- Finér, L.; Jurgensen, M.F.; Domisch, T.; Kilpeläinen, J.; Neuvonen, S.; Punttila, P.; Risch, A.C.; Ohashi, M.; Niemelä, P. The Role of Wood Ants (Formica rufa group) in Carbon and Nutrient Dynamics of a Boreal Norway Spruce Forest Ecosystem. Ecosystems 2013, 16, 196–208. [Google Scholar] [CrossRef]
- Andersen, A.N. Responses of ant communities to disturbance: Five principles for understanding the disturbance dynamics of a globally dominant faunal group. J. Anim. Ecol. 2019, 88, 350–362. [Google Scholar] [CrossRef]
- Kaukonen, M.; Ruotsalainen, A.L.; Wäli, P.R.; Männistö, M.K.; Setälä, H.; Saravesi, K.; Huusko, K.; Markkola, A. Moth herbivory enhances resource turnover in subarctic mountain birch forests? Ecology 2013, 94, 267–272. [Google Scholar] [CrossRef]
- Mikola, J.; Yeates, G.W.; Barker, G.M.; Wardle, D.A.; Bonner, K.I. Effects of defoliation intensity on soil food-web properties in an experimental grassland community. Oikos 2001, 92, 333–343. [Google Scholar] [CrossRef]
- Pitman, R.M.; Vanguelova, E.I.; Benham, S.E. The effects of phytophagous insects on water and soil nutrient concentrations and fluxes through forest stands of the Level II monitoring network in the UK. Sci. Total Environ. 2010, 409, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Chapman, S.K.; Hart, S.C.; Cobb, N.S.; Whitham, T.G.; Koch, G.W. Insect herbivory increases litter quality and decomposition: An extension of the acceleration hypothesis. Ecology 2003, 84, 2867–2876. [Google Scholar] [CrossRef]
- Moya-Laraño, J.; Wise, D.H. Direct and indirect effects of ants on a forest-floor food web. Ecology 2007, 88, 1454–1465. [Google Scholar] [CrossRef] [PubMed]
- Çakır, M. The negative effect of wood ants (Formica rufa) on microarthropod density and soil biological quality in a semi-arid pine forest. Pedobiologia 2019, 77, 150593. [Google Scholar] [CrossRef]
- Karhu, K.J.; Neuvonen, S. Wood ants and a geometrid defoliator of birch: Predation outweighs beneficial effects through the host plant. Oecologia 1998, 113, 509–516. [Google Scholar] [CrossRef]
- Way, M.J.; Khoo, K.C. Role of Ants in Pest Management. Annu. Rev. Entomol. 1992, 37, 479–503. [Google Scholar] [CrossRef]
- Tilman, D. Cherries, Ants and Tent Caterpillars: Timing of Nectar Production in Relation to Susceptibility of Caterpillars to Ant Predation. Ecology 1978, 59, 686–692. [Google Scholar] [CrossRef]
- Gösswald, K. Die Waldameise. Band 2. Die Waldameise in Okosystem Wald, ihr Nutzen und ihre Hege; Aula Verlag: Totnes, UK, 1990; Volume 510S. [Google Scholar]
- Carvalho, K.S.; Vasconcelos, H.L. Forest fragmentation in central Amazonia and its effects on litter-dwelling ants. Biol. Conserv. 1999, 91, 151–157. [Google Scholar] [CrossRef]
- Maeto, K.; Sato, S. Impacts of forestry on ant species richness and composition in warm-temperate forests of Japan. For. Ecol. Manag. 2004, 187, 213–223. [Google Scholar] [CrossRef]
- Palladini, J.D.; Jones, M.G.; Sanders, N.J.; Jules, E.S. The recovery of ant communities in regenerating temperate conifer forests. For. Ecol. Manag. 2007, 242, 619–624. [Google Scholar] [CrossRef]
- Ewers, R.M.; Boyle, M.J.; Gleave, R.A.; Plowman, N.S.; Benedick, S.; Bernard, H.; Bishop, T.R.; Bakhtiar, E.Y.; Chey, V.K.; Chung, A.Y. Logging cuts the functional importance of invertebrates in tropical rainforest. Nat. Commun. 2015, 6, 6836. [Google Scholar] [CrossRef] [PubMed]
- Punttila, P.; Haila, Y.; Niemelä, J.; Pajunen, T. Ant communities in fragments of old-growth taiga and managed surroundings. In Proceedings of the Annales Zoologici Fennici; JSTOR; Finnish Zoological and Botanical Publishing Board: Helsinki, Finland, 1994; pp. 131–144. [Google Scholar]
- Véle, A.; Holuša, J.; Horák, J. Ant abundance increases with clearing size. J. For. Res. 2016, 21, 110–114. [Google Scholar] [CrossRef]
- Grevé, M.E.; Hager, J.; Weisser, W.W.; Schall, P.; Gossner, M.M.; Feldhaar, H. Effect of forest management on temperate ant communities. Ecosphere 2018, 9, e02303. [Google Scholar] [CrossRef]
- Farji-Brener, A. Why are leaf-cutting ants more common in early secondary forests than in old-growth tropical forests? An evaluation of the palatable forage hypothesis. Oikos 2001, 92, 169–177. [Google Scholar] [CrossRef]
- Yamamoto, S.-I. Forest gap dynamics and tree regeneration. J. For. Res. 2000, 5, 223–229. [Google Scholar] [CrossRef]
- Punttila, P.; Haila, Y.; Pajunen, T.; Tukia, H. Colonisation of clearcut forests by ants in the southern Finnish taiga: A quantitative survey. Oikos 1991, 61, 250–262. [Google Scholar] [CrossRef]
- Schall, P.; Gossner, M.M.; Heinrichs, S.; Fischer, M.; Boch, S.; Prati, D.; Jung, K.; Baumgartner, V.; Blaser, S.; Böhm, S. The impact of even-aged and uneven-aged forest management on regional biodiversity of multiple taxa in European beech forests. J. Appl. Ecol. 2018, 55, 267–278. [Google Scholar] [CrossRef]
- Fitzgerald, T. The Tent Caterpillars; Cornell University Press: Ithaca, NY, USA, 1995. [Google Scholar]
- Moulinier, J. Impacts de la défoliation par la livrée des forêts sur la mortalité du peuplier faux-tremble et la dynamique forestière post-épidémie en forêt boréale. Ph.D. Thesis, Université du Québec en Abitibi-Témiscamingue, Rouyn-Noranda, QC, Canada, 2013. [Google Scholar]
- Lach, L.; Parr, C.; Abbott, K. Ant Ecology; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Montllor, C.B.; Bernays, E.A. Invertebrate Predators and Caterpillar Foraging; Chapman and Hall: New York, NY, USA, 1993. [Google Scholar]
- Piñol, J.; Espadaler, X.; Cañellas, N.; MARTÍNEZ-VILALTA, J.; Barrientos, J.A.; Sol, D. Ant versus bird exclusion effects on the arthropod assemblage of an organic citrus grove. Ecol. Entomol. 2010, 35, 367–376. [Google Scholar] [CrossRef]
- Clark, R.E.; Farkas, T.E.; Lichter-Marck, I.; Johnson, E.R.; Singer, M.S. Multiple interaction types determine the impact of ant predation of caterpillars in a forest community. Ecology 2016, 97, 3379–3388. [Google Scholar] [CrossRef]
- Green, G.; Sullivan, C. Ants Attacking Larvae of the Forest Tent Caterpillar, Malacosoma disstria Hbn. (Lepidoptera: Lasiocampidae). Can. Entomol. 1950, 82, 94–195. [Google Scholar] [CrossRef]
- Despland, E.; Lessard, J.-P. Social predation by ants as a mortality source for an arboreal gregarious forest pest. Basic Appl. Ecol. 2022, 59, 82–91. [Google Scholar] [CrossRef]
- Rosumek, F.B.; Silveira, F.A.O.; Neves, F.D.S.; Barbosa, N.P.D.U.; Diniz, L.; Oki, Y.; Pezzini, F.; Fernandes, G.W.; Cornelissen, T. Ants on plants: A meta-analysis of the role of ants as plant biotic defenses. Oecologia 2009, 160, 537–549. [Google Scholar] [CrossRef]
- Romero, G.Q.; Antiqueira, P.A.; Koricheva, J. A meta-analysis of predation risk effects on pollinator behaviour. PLoS ONE 2011, 6, e20689. [Google Scholar] [CrossRef]
- Moreira, X.; Mooney, K.A.; Zas, R.; Sampedro, L. Bottom-up effects of host-plant species diversity and top-down effects of ants interactively increase plant performance. Proc. R. Soc. B Biol. Sci. 2012, 279, 4464–4472. [Google Scholar] [CrossRef]
- Cerdá, X.; Dejean, A. 3. Predation by ants on arthropods and other animals. In Predation in the Hymenoptera: An Evolutionary Perspective; Transworld Research Network: Trivandrum, India, 2011. [Google Scholar]
- MFFP. Aires Infestées par la Livrée des Forêts au Québec en 2016; Gouvernement du Québec, Direction de la protection des Forêts: Québec, QC, Canada, 2016; p. 10. [Google Scholar]
- MFFP. Aires Infestées par la Livrée des Forêts au Québec en 2017; Gouvernement du Québec, Direction de la protection des forêts: Québec, QC, Canada, 2017; p. 14. [Google Scholar]
- Bergeron, Y. Species and stand dynamics in the mixed woods of Quebec’s southern boreal forest. Ecology 2000, 81, 1500–1516. [Google Scholar] [CrossRef]
- Agriculture Canada Expert Committee on Soil Survey. The Canadian System of Soil Classification (Second Edition). Available online: https://sis.agr.gc.ca/cansis/publications/manuals/1987-cssc-ed2/index.html (accessed on 3 November 2022).
- Vincent, J.-S.; Hardy, L. L’évolution et l’extension des lacs glaciaires Barlow et Ojibway en territoire québécois. Géographie Phys. Quat. 1977, 31, 357–372. [Google Scholar] [CrossRef]
- MFFP. Aires Infestées par la Livrée des Forêts au Québec en 2015; Gouvernement du Québec, Direction de la protection des Forêts: Québec, QC, Canada, 2015; p. 8. [Google Scholar]
- MFFP. Aires Infestées par la Livrée des Forêts au Québec en 2018; Gouvernement du Québec, Direction de la protection des Forêts: Québec, QC, Canada, 2019; p. 14. [Google Scholar]
- Morin, H.; Laprise, D.; Bergeron, Y. Chronology of spruce budworm outbreaks near Lake Duparquet, Abitibi region, Quebec. Can. J. For. Res. 1993, 23, 1497–1506. [Google Scholar] [CrossRef]
- Dubuc, J.-F. La Performance Biologique et le Comportement Alimentaire de trois espèces de Lépidoptères Après Défoliation du Bouleau Blanc (Betula papyrifera marsh.); Université du Québec à Montréal: Montreal, QC, Canada, 1996. [Google Scholar]
- Environment Canada. National Climate Data and Information Archive; Environment Canada: Ottawa, ON, Canada, 2017. [Google Scholar]
- Lajoie, P.G. Étude pédologique des comtés de Hull, Labelle et Papineau; Québec Ottawa Ministère Iagriculture: Ottawa, ON, Canada, 1967. [Google Scholar]
- Bestelmeyer, B.T.; Agosti, D.; Alonso, L.E.; Brandão, C.R.F.; Brown, W.L.; Delabie, J.H.; Silvestre, R. Field techniques for the study of ground-dwelling ant: An overview, description, and evaluation. In Ants: Standard Methods for Measuring and Monitoring Biodiversity; Smithsonian Institution Press: Washington, DC, USA, 2000. [Google Scholar]
- Mahon, M.B.; Campbell, K.U.; Crist, T.O. Effectiveness of Winkler Litter Extraction and Pitfall Traps in Sampling Ant Communities and Functional Groups in a Temperate Forest. Environ. Entomol. 2017, 46, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Francœur, A. Les fourmis de la forêt boréale du Québec (Formicidae, Hymenoptera). Nat. Can. 2001, 125, 8. [Google Scholar]
- Ellison, A.M.; Gotelli, N.J.; Farnsworth, E.J.; Alpert, G.D. A Field Guide to the Ants of New England; Yale University Press: London, UK, 2012; ISBN 978-0-300-16930-0. [Google Scholar]
- Hopkin, S.P. A Key to the Collembola (Springtails) of Britain and Ireland; FSC Publications: Shrewsbury, UK, 2007. [Google Scholar]
- Fjellberg, A. The Collembola of Fennoscandia and Denmark. Part 1: Poduromorpha Fauna. In Entomologica Scandinavica; Brill Academic: Leiden, The Netherlands, 1998. [Google Scholar]
- Fjellberg, A. The Collembola of Fennoscandia and Denmark, Part II: Entomobryomorpha and Symphypleona; Brill: Leiden, The Netherlands, 2007. [Google Scholar]
- Christiansen, K.; Bellinger, P. The Collembola of North America North of the Rio Grande. A Taxonomic Analysis. Part 1. Introduction. General. Families Poduridae and Hypogastruridae. Part 2. Families Onychiuridae and Isotomidae. Part 3. Family Entomobrydae. Part 4. Families Neelidae and Sminthuridae. Glossary. Bibliography. Index; Grinnell College: Grinnell, IA, USA, 1980. [Google Scholar]
- Koudji, E.G.; Despland, E.; Caron, A.; Handa, I.T. Soil Springtail Communities Are Resilient to Forest Tent Caterpillar Defoliation in Quebec Mixed Hardwood Forests. Forests, 2023; submitted. [Google Scholar]
- Evans, E. Influence of weather on predator/prey relations: Stinkbugs and tent caterpillars. J. N. Y. Entomol. Soc. 1982, 90, 241–246. [Google Scholar]
- Fitzgerald, T.; Costa, J. Collective behavior in social caterpillars. In Information Processing in Social Insects; Birkhäuser: Basel, Switzerland, 1999; pp. 379–400. [Google Scholar]
- McClure, M.; Despland, E. Collective foraging patterns of field colonies of Malacosoma disstria caterpillars. Can. Entomol. 2010, 142, 473–480. [Google Scholar] [CrossRef]
- Ronnås, C.; Larsson, S.; Pitacco, A.; Battisti, A. Effects of colony size on larval performance in a processionary moth. Ecol. Entomol. 2010, 35, 436–445. [Google Scholar] [CrossRef]
- Cobbold, C.A.; Lewis, M.A.; Lutscher, F.; Roland, J. How parasitism affects critical patch-size in a host–parasitoid model: Application to the forest tent caterpillar. Theor. Popul. Biol. 2005, 67, 109–125. [Google Scholar] [CrossRef]
- Sorensen, T.A. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons. Biol. Skar. 1948, 5, 1–34. [Google Scholar]
- Jaccard, P. Étude comparative de la distribution florale dans une portion des Alpes et des Jura. Bull. Soc. Vaud. Sci. Nat. 1901, 37, 547–579. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 1.17-4. Acesso Em 2010, 23, 2010. Available online: http://cran.r-project.org> (accessed on 5 December 2022).
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Caceres, M.; Jansen, F. Package ‘indicspecies’. Indicators 2016, 8. [Google Scholar]
- Team R Development Core. A Language and Environment for Statistical Computing; The R Foundation for Statistical Computing, Department of Agronomy, Faculty: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 2 February 2023).
- Broza, M.; Izhaki, I. Post-fire arthropod assemblages in Mediterranean forest soils in Israel. Int. J. Wildland Fire 1997, 7, 317–325. [Google Scholar] [CrossRef]
- Gardner, S.M.; Cabido, M.R.; Valladares, G.R.; Diaz, S. The influence of habitat structure on arthropod diversity in Argentine semi-arid Chaco forest. J. Veg. Sci. 1995, 6, 349–356. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Van Der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Bowman, W.D.; Kaufmann, R.; Schmidt, S.K. A temporal approach to linking aboveground and belowground ecology. Trends Ecol. Evol. 2005, 20, 634–641. [Google Scholar] [CrossRef]
- Hedlund, K.; Griffiths, B.; Christensen, S.; Scheu, S.; Setälä, H.; Tscharntke, T.; Verhoef, H. Trophic interactions in changing landscapes: Responses of soil food webs. Basic Appl. Ecol. 2004, 5, 495–503. [Google Scholar] [CrossRef]
- Anderson, K.E.; Russell, J.A.; Moreau, C.S.; Kautz, S.; Sullam, K.E.; Hu, Y.I.; Basinger, U.; Mott, B.M.; Buck, N.; Wheeler, D.E. Highly similar microbial communities are shared among related and trophically similar ant species. Mol. Ecol. 2012, 21, 2282–2296. [Google Scholar] [CrossRef]
- Souza, R.F.; Anjos, D.V.; Carvalho, R.; Del-Claro, K. Availability of food and nesting-sites as regulatory mechanisms for the recovery of ant diversity after fire disturbance. Sociobiology 2015, 62, 1–9. [Google Scholar] [CrossRef]
- Frizzo, T.L.; Campos, R.I.; Vasconcelos, H.L. Contrasting effects of fire on arboreal and ground-dwelling ant communities of a Neotropical savanna. Biotropica 2012, 44, 254–261. [Google Scholar] [CrossRef]
- Silveira, J.M.; Louzada, J.; Barlow, J.; Andrade, R.; Mestre, L.; Solar, R.; Lacau, S.; Cochrane, M.A. A multi-taxa assessment of biodiversity change after single and recurrent wildfires in a Brazilian Amazon forest. Biotropica 2016, 48, 170–180. [Google Scholar] [CrossRef]
- Vasconcelos, H.L.; Maravalhas, J.B.; Cornelissen, T. Effects of fire disturbance on ant abundance and diversity: A global meta-analysis. Biodivers. Conserv. 2017, 26, 177–188. [Google Scholar] [CrossRef]
- Calizza, E.; Rossi, L.; Careddu, G.; Sporta Caputi, S.; Costantini, M.L. Species richness and vulnerability to disturbance propagation in real food webs. Sci. Rep. 2019, 9, 19331. [Google Scholar] [CrossRef]
- Cooke, B.J.; Sturtevant, B.R.; Robert, L.-E. The Forest Tent Caterpillar in Minnesota: Detectability, Impact, and Cycling Dynamics. Forests 2022, 13, 601. [Google Scholar] [CrossRef]
- Andersen, A.N.; Penman, T.D.; Debas, N.; Houadria, M. Ant community responses to experimental fire and logging in a eucalypt forest of south-eastern Australia. For. Ecol. Manag. 2009, 258, 188–197. [Google Scholar] [CrossRef]
- York, A. Long-term effects of frequent low-intensity burning on ant communities in coastal blackbutt forests of southeastern Australia. Austral Ecol. 2000, 25, 83–98. [Google Scholar] [CrossRef]
- Parr, C.L.; Robertson, H.G.; Biggs, H.C.; Chown, S.L. Response of African savanna ants to long-term fire regimes. J. Appl. Ecol. 2004, 41, 630–642. [Google Scholar] [CrossRef]
- Maravalhas, J.; Vasconcelos, H.L. Revisiting the pyrodiversity–biodiversity hypothesis: Long-term fire regimes and the structure of ant communities in a N eotropical savanna hotspot. J. Appl. Ecol. 2014, 51, 1661–1668. [Google Scholar] [CrossRef]
- Johns, R.C.; Bowden, J.J.; Carleton, D.R.; Cooke, B.J.; Edwards, S.; Emilson, E.J.S.; James, P.M.A.; Kneeshaw, D.; MacLean, D.A.; Martel, V.; et al. A Conceptual Framework for the Spruce Budworm Early Intervention Strategy: Can Outbreaks be Stopped? Forests 2019, 10, 910. [Google Scholar] [CrossRef]
- Lessard, J.-P.; Dunn, R.R.; Sanders, N.J. Temperature-mediated coexistence in temperate forest ant communities. Insectes Sociaux 2009, 56, 149–156. [Google Scholar] [CrossRef]
- Fotso Kuate, A.; Hanna, R.; Tindo, M.; Nanga, S.; Nagel, P. Ant Diversity in Dominant Vegetation Types of Southern Cameroon. Biotropica 2015, 47, 94–100. [Google Scholar] [CrossRef]
- Leal, I.R.; Filgueiras, B.K.C.; Gomes, J.P.; Iannuzzi, L.; Andersen, A.N. Effects of habitat fragmentation on ant richness and functional composition in Brazilian Atlantic forest. Biodivers. Conserv. 2012, 21, 1687–1701. [Google Scholar] [CrossRef]
- Solar, R.R.D.C.; Barlow, J.; Andersen, A.N.; Schoereder, J.H.; Berenguer, E.; Ferreira, J.N.; Gardner, T.A. Biodiversity consequences of land-use change and forest disturbance in the Amazon: A multi-scale assessment using ant communities. Biol. Conserv. 2016, 197, 98–107. [Google Scholar] [CrossRef]
- Fairweather, A.D.; Lewis, J.H.; Hunt, L.; McAlpine, D.F.; Smith, M.A. Ants (Hymenoptera: Formicidae) of Rockwood Park, New Brunswick: An Assessment of Species Richness and Habitat. Northeast. Nat. 2020, 27, 576. [Google Scholar] [CrossRef]
- Milford, E.R. Ant communities in flooded and unflooded riparian forest of the middle Rio Grande. Southwest. Nat. 1999, 44, 278–286. [Google Scholar]
- Ellison, A.M.; Farnsworth, E.J.; Gotelli, N.J. Ant diversity in pitcher-plant bogs of Massachusetts. Northeast. Nat. 2002, 9, 267–284. [Google Scholar] [CrossRef]
- Francoeur, A. Deux nouvelles fourmis néarctiques: Leptothorax retractus et L. sphagnicolus (Formicidae, Hymenoptera). Can. Entomol. 1986, 118, 1151–1164. [Google Scholar] [CrossRef]
- Francoeur, A. Révision taxonomique des espèces néarctiques du groupe Fusca, genre Formica (Formicidae, Hymenoptera). Mem. Soc. ent. Queb. 3. 316 pp. 1979. Formicoidea. Can. Its Insect Fauna Mem. Ent. Soc. Can. 1973, 108, 502–503. [Google Scholar]
- Francoeur, A. Extension de l’aire connue de la fourmi Myrmica quebecensis (Formicidae, Hymenoptera). 2011. Available online: https://www.erudit.org/fr/revues/natcan/2018-v142-n1-natcan03265/1042014ar.pdf (accessed on 2 February 2023).
- Oberg, E.; Del Toro, I.; Pelini, S. Characterization of the thermal tolerances of forest ants of New England. Insectes Sociaux 2012, 59, 167–174. [Google Scholar] [CrossRef]
- Wheeler, G.C.; Wheeler, J. The Ants of North Dakota; University of North Dakota Grand Forks: Grand Forks, ND, USA, 1963. [Google Scholar]
- MacKay, W.P.; Mackay, E. The Ants of New Mexico (Hymenoptera: Formicidae); Edwin Mellen Press: Lewiston, NY, USA, 2002. [Google Scholar]
- Sirois, L. Impact of fire on Picea mariana and Pinus banksiana seedlings in subarctic lichen woodlands. J. Veg. Sci. 1993, 4, 795–802. [Google Scholar] [CrossRef]
- Rosengren, R. The interaction between red wood ants, Cinara aphids, and pines. A ghost of mutualism past? In Ant-Plant Interactions; Oxford University Press: Oxford, UK, 1991; pp. 80–91. [Google Scholar]
- Parmentier, T.; Dekoninck, W.; Wenseleers, T. A highly diverse microcosm in a hostile world: A review on the associates of red wood ants (Formica rufa group). Insectes Sociaux 2014, 61, 229–237. [Google Scholar] [CrossRef]
- Kilpeläinen, J.; Finér, L.; Niemelä, P.; Domisch, T.; Neuvonen, S.; Ohashi, M.; Risch, A.C.; Sundström, L. Carbon, nitrogen and phosphorus dynamics of ant mounds (Formica rufa group) in managed boreal forests of different successional stages. Appl. Soil Ecol. 2007, 36, 156–163. [Google Scholar] [CrossRef]
- Serttaş, A.; Bakar, Ö.; Alkan, U.M.; Yılmaz, A.; Yolcu, H.I.; Ipekdal, K. Nest Survival and Transplantation Success of Formica rufa (Hymenoptera: Formicidae) Ants in Southern Turkey: A Predictive Approach. Forests 2020, 11, 533. [Google Scholar] [CrossRef]
- Stockan, J.A.; Rao, S.; Pakeman, R. Nesting preferences of the threatened wood ant Formica exsecta (Hymenoptera: Formicidae); implications for conservation in Scotland. J. Insect Conserv. 2010, 14, 269–276. [Google Scholar] [CrossRef]
- Rosengren, R.; Fortelius, W.; Lindström, K.; Luther, A. Phenology and causation of nest heating and thermoregulation in red wood ants of the Formica rufa group studied in coniferous forest habitats in southern Finland. In Proceedings of the Annales Zoologici Fennici; JSTOR; Finnish Academy of Sciences, Societas Scientiarum Fennica, Societas pro Fauna et Flora Fennica and Societas Biologica Fennica Vanamo: Helsinki, Finland, 1987; pp. 147–155. [Google Scholar]
- Weseloh, R.M. Simulation of predation by ants based on direct observations of attacks on gypsy moth larvae. Can. Èntomol. 1989, 121, 1069–1076. [Google Scholar] [CrossRef]
- Mokadam, C. Native and Non-Native Ant Impacts on Native Fungi. Ph.D. Thesis, Buffalo State College, Buffalo, NY, USA, 2021. [Google Scholar]
- Sabelis, M.W. Predatory arthropods. Nat. Enemies Popul. Biol. Predat. Parasites Dis. 1992, 225–264. [Google Scholar] [CrossRef]
- Settle, W.H.; Ariawan, H.; Astuti, E.T.; Cahyana, W.; Hakim, A.L.; Hindayana, D.; Lestari, A.S. Managing tropical rice pests through conservation of generalist natural enemies and alternative prey. Ecology 1996, 77, 1975–1988. [Google Scholar] [CrossRef]
- Symondson, W.; Sunderland, K.; Greenstone, M. Can Generalist Predators Be Effective Biocontrol Agents. Annu. Rev. Entomol. 2002, 47, 561–594. [Google Scholar] [CrossRef]
- Yang, L.H. Interactions between a detrital resource pulse and a detritivore community. Oecologia 2006, 147, 522–532. [Google Scholar] [CrossRef]
- Karban, R. Increased reproductive success at high densities and predator satiation for periodical cicadas. Ecology 1982, 63, 321–328. [Google Scholar] [CrossRef]
- Sweeney, B.W.; Vannote, R.L. Population synchrony in mayflies: A predator satiation hypothesis. Evolution 1982, 36, 810–821. [Google Scholar] [CrossRef]
- Williams, K.S.; Smith, K.G.; Stephen, F.M. Emergence of 13-Yr periodical cicadas (Cicadidae: Magicicada): Phenology, mortality, and predators satiation. Ecology 1993, 74, 1143–1152. [Google Scholar] [CrossRef]
- Yang, L.H.; Edwards, K.F.; Byrnes, J.E.; Bastow, J.L.; Wright, A.N.; Spence, K.O. A meta-analysis of resource pulse–consumer interactions. Ecol. Monogr. 2010, 80, 125–151. [Google Scholar] [CrossRef]
- Eveleigh, E.S.; McCann, K.S.; McCarthy, P.C.; Pollock, S.J.; Lucarotti, C.J.; Morin, B.; McDougall, G.A.; Strongman, D.B.; Huber, J.T.; Umbanhowar, J.; et al. Fluctuations in density of an outbreak species drive diversity cascades in food webs. Proc. Natl. Acad. Sci. USA 2007, 104, 16976–16981. [Google Scholar] [CrossRef]
- Kristensen, J.Å.; Rousk, J.; Metcalfe, D.B. Below-ground responses to insect herbivory in ecosystems with woody plant canopies: A meta-analysis. J. Ecol. 2020, 108, 917–930. [Google Scholar] [CrossRef]
- Banschbach, V.S.; Herbers, J.M. Nest Movements and Population Spatial Structure of the Forest Ant Myrmica Punctiventris (Hymenoptera: Formicidae). Ann. Entomol. Soc. Am. 1999, 92, 414–423. [Google Scholar] [CrossRef]
- Francoeur, A. The Ant Fauna near the Tree-Line in Northern Québec (Formicidae, Hymenoptera). Nordicana 1983, 47, 177–180. [Google Scholar]
- Francoeur, A. Ants (Hymenoptera: Formicidae) of the Yukon. 1997. Available online: https://www.researchgate.net/publication/237749861_Ants_Hymenoptera_Formicidae_of_the_Yukon (accessed on 2 February 2023).




{kind=link}
{kind=link}
{kind=link}
| Boreal Forest | Temperate Forest | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| df | F | R2 | p-Value | df | F | R2 | p-Value | ||
| Defoliation history | 1 | 5.3914 | 0.09096 | 0.002 | Defoliation history | 1 | 1.4536 | 0.10198 | 0.249 |
| Canopy openness | 1 | 0.9813 | 0.01656 | 0.434 | Canopy openness | 1 | 0.3802 | 0.02667 | 0.830 |
| Predators | 1 | 4.0690 | 0.06865 | 0.004 | Predators | 1 | 0.7809 | 0.05478 | 0.567 |
| Collembola (abundance) | 1 | 0.5300 | 0.00894 | 0.764 | Collembola (abundance) | 1 | 1.6389 | 0.11498 | 0.181 |
| Collembola (species richness) | 1 | 1.2997 | 0.02193 | 0.283 | |||||
| Residuals | 47 | 0.79296 | 14 | 0.70158 | |||||
| Groups | Species | A | B | IVI * | p-Value |
|---|---|---|---|---|---|
| Control | Camponotus herculeanus | 0.7812 | 0.5714 | 0.668 | 0.002 |
| Myrmica sp1 | 0.9146 | 0.4286 | 0.626 | 0.003 | |
| Myrmica detritinodis | 0.8893 | 0.3214 | 0.535 | 0.011 | |
| Formica sp1 | 1.0000 | 0.2857 | 0.535 | 0.004 | |
| Outbreak | Formica integra | 1.0 | 0.2 | 0.447 | 0.024 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caron, A.-S.; Koudji, E.G.; Handa, I.T.; Montoro Girona, M.; Despland, E. Forest Tent Caterpillar Outbreaks Drive Change in Ant Communities in Boreal Forests. Forests 2023, 14, 1147. https://doi.org/10.3390/f14061147
Caron A-S, Koudji EG, Handa IT, Montoro Girona M, Despland E. Forest Tent Caterpillar Outbreaks Drive Change in Ant Communities in Boreal Forests. Forests. 2023; 14(6):1147. https://doi.org/10.3390/f14061147
Chicago/Turabian StyleCaron, Anne-Sophie, Essivi Gagnon Koudji, Ira Tanya Handa, Miguel Montoro Girona, and Emma Despland. 2023. "Forest Tent Caterpillar Outbreaks Drive Change in Ant Communities in Boreal Forests" Forests 14, no. 6: 1147. https://doi.org/10.3390/f14061147
APA StyleCaron, A.-S., Koudji, E. G., Handa, I. T., Montoro Girona, M., & Despland, E. (2023). Forest Tent Caterpillar Outbreaks Drive Change in Ant Communities in Boreal Forests. Forests, 14(6), 1147. https://doi.org/10.3390/f14061147