
Forest Gaps Slow the Humification Process of Fir (Abies faxoniana Rehder & E.H.Wilson) Twig Litter during Eight Years of Decomposition in an Alpine Forest

 , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Sample Collection and Chemical Analysis
2.4. Data Calculations and Statistical Analyses Statistical Analyses and Calculations
3. Results
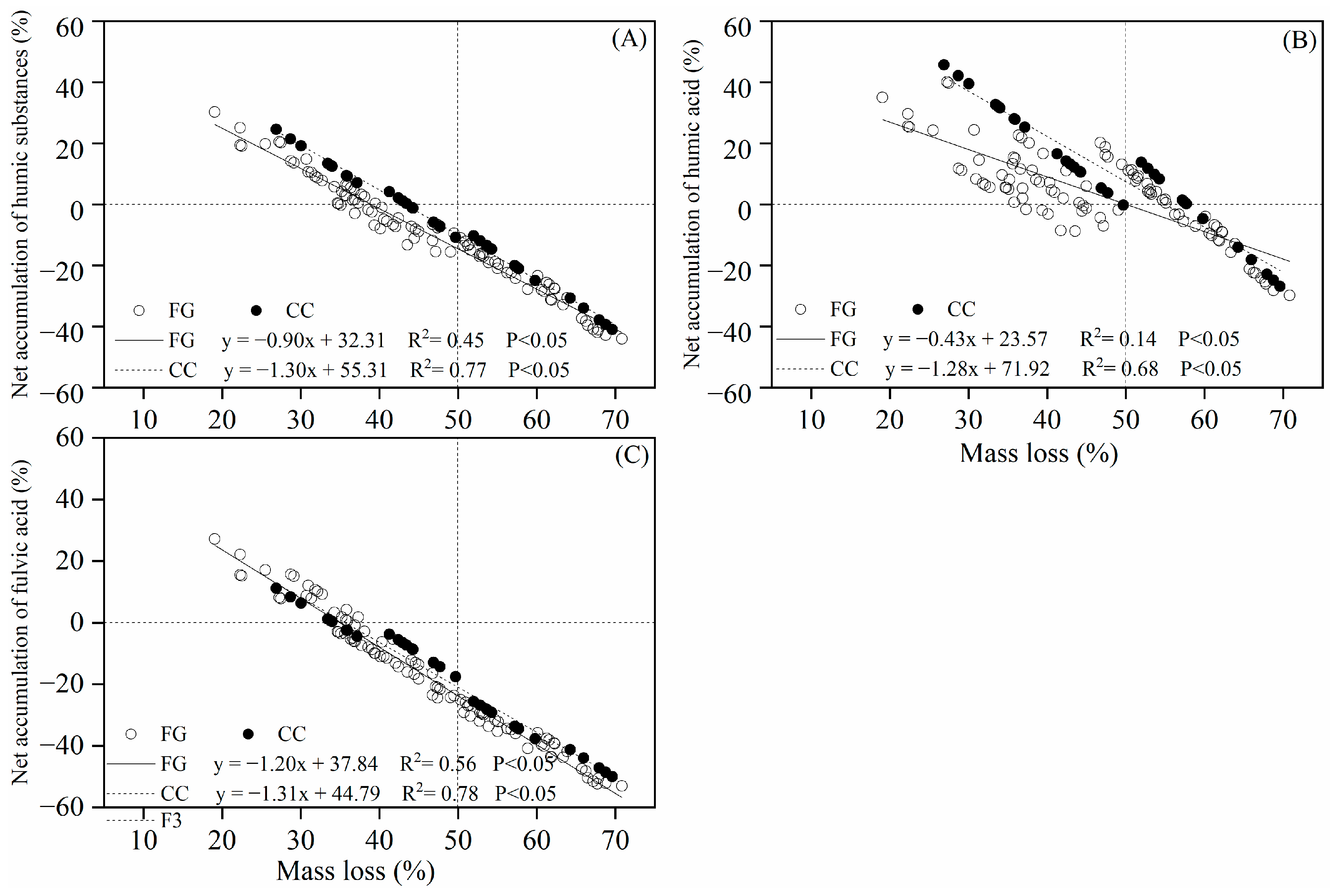
3.1. Accumulation of Humus
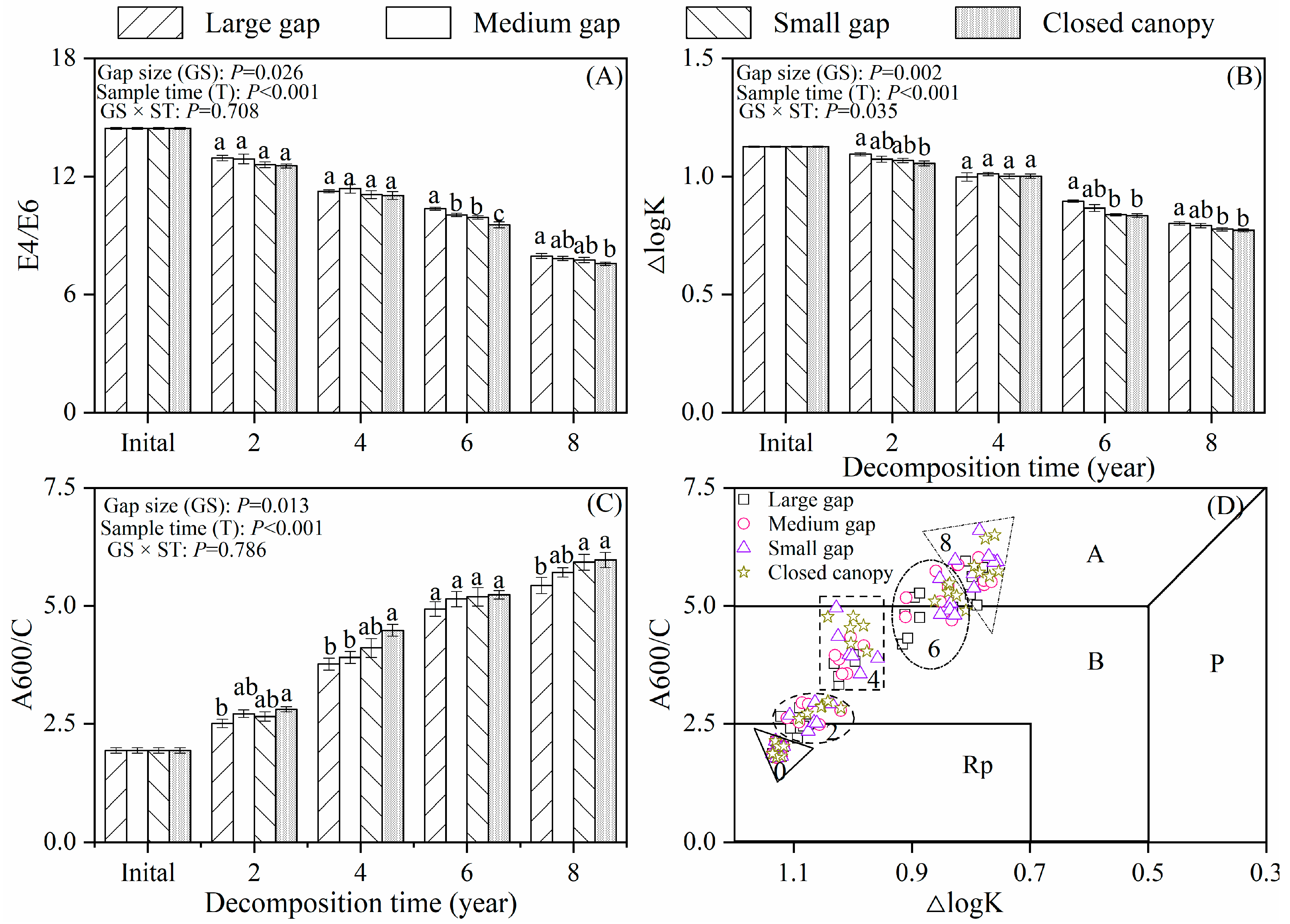
3.2. Humification Degree and Humification Rate
3.3. Optical Properties of Extracted Alkaline Solutions (Humic Acid-like)
3.4. Controlling Factors
4. Discussion
4.1. Effect of Forest Gaps on Twig Litter Humification Accumulation
4.2. Temporal Dynamics of Twig Litter Humification
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Decomposition Time (Year) | Large Gap | Medium Gap | Small Gap | Closed Canopy |
|---|---|---|---|---|---|
| Carbon content (g/kg) | Initial | 441.88 ± 10.77 | 441.88 ± 10.77 | 441.88 ± 10.77 | 441.88 ± 10.77 |
| 2 | 455.47 ± 2.92b | 460.29 ± 5.59b | 463.63 ± 11.28b | 493.28 ± 0.93a | |
| 4 | 463.55 ± 6.63a | 458.15 ± 8.39a | 451.90 ± 14.82a | 461.47 ± 6.36a | |
| 6 | 417.10 ± 10.99a | 395.82 ± 10.46ab | 391.96 ± 11.00ab | 382.73 ± 7.71b | |
| 8 | 350.27 ± 9.84a | 356.40 ± 44.56a | 382.59 ± 24.88a | 396.22 ± 37.34a | |
| Nitrogen content (g/kg) | Initial | 8.11 ± 0.16 | 8.11 ± 0.16 | 8.11 ± 0.16 | 8.11 ± 0.16 |
| 2 | 6.86 ± 0.36a | 6.39 ± 0.20a | 7.53 ± 0.72a | 6.45 ± 0.35a | |
| 4 | 6.00 ± 0.42a | 4.76 ± 0.35b | 4.6 ± 0.24bc | 3.72 ± 0.08c | |
| 6 | 6.66 ± 0.28ab | 6.13 ± 0.30b | 6.67 ± 0.21ab | 7.19 ± 0.36a | |
| 8 | 10.41 ± 0.44a | 9.24 ± 0.64ab | 8.44 ± 0.42b | 9.99 ± 0.27ab | |
| Phosphorus content (g/kg) | Initial | 0.67 ± 0.03 | 0.67 ± 0.03 | 0.67 ± 0.03 | 0.67 ± 0.03 |
| 2 | 0.68 ± 0.04a | 0.71 ± 0.09a | 0.62 ± 0.03a | 0.58 ± 0.04a | |
| 4 | 0.27 ± 0.01a | 0.24 ± 0.00ab | 0.25 ± 0.00ab | 0.23 ± 0.00b | |
| 6 | 0.55 ± 0.02bc | 0.68 ± 0.02a | 0.60 ± 0.01ab | 0.54 ± 0.01c | |
| 8 | 0.47 ± 0.00a | 0.47 ± 0.00a | 0.47 ± 0.00a | 0.47 ± 0.00a | |
| Lignin content (%) | Initial | 32.40 ± 0.00 | 32.40 ± 0.00 | 32.40 ± 0.00 | 32.40 ± 0.00 |
| 2 | 35.82 ± 0.60b | 36.70 ± 0.13 b | 39.38 ± 0.39a | 38.70 ± 0.32a | |
| 4 | 43.44 ± 1.50a | 44.28 ± 0.75a | 39.59 ± 0.86b | 42.61 ± 0.58ab | |
| 6 | 44.96 ± 0.58ab | 45.32 ± 1.09a | 44.80 ± 1.07ab | 43.12 ± 0.60b | |
| 8 | 41.28 ± 1.55ab | 39.66 ± 0.68b | 43.83 ± 0.64a | 39.44 ± 0.92b | |
| Cellulose content (%) | Initial | 23.81 ± 0.00 | 23.81 ± 0.00 | 23.81 ± 0.00 | 23.81 ± 0.00 |
| 2 | 21.96 ± 0.68b | 23.58 ± 1.24b | 21.00 ± 1.59a | 22.06 ± 0.76ab | |
| 4 | 18.83 ± 0.85ab | 18.33 ± 0.47a | 19.51 ± 0.39ab | 17.21 ± 1.31b | |
| 6 | 17.22 ± 0.90ab | 16.72 ± 1.04a | 17.96 ± 0.90ab | 16.32 ± 0.80b | |
| 8 | 13.59 ± 0.23ab | 12.72 ± 0.89ab | 13.79 ± 0.60a | 11.48 ± 0.64b | |
| C/N | Initial | 54.46 ± 0.87 | 54.46 ± 0.87 | 54.46 ± 0.87 | 54.46 ± 0.87 |
| 2 | 66.73 ± 2.96a | 72.13 ± 1.66a | 62.4 ± 4.33a | 66.97 ± 4.05a | |
| 4 | 77.84 ± 4.16c | 97.06 ± 6.05b | 98.57 ± 2.45b | 120.99 ± 5.31a | |
| 6 | 63.38 ± 3.00ab | 65.97 ± 3.45a | 57.8 ± 2.06ab | 54.72 ± 3.04b | |
| 8 | 33.74 ± 1.39b | 35.23 ± 3.56ab | 47.29 ± 5.77a | 42.34 ± 3.16ab | |
| C/P | Initial | 657.42 ± 21.36 | 657.42 ± 21.36 | 657.42 ± 21.36 | 657.42 ± 21.36 |
| 2 | 678.57 ± 36.23a | 672.24 ± 78.15a | 749.54 ± 22.77a | 865.20 ± 69.57a | |
| 4 | 1724.22 ± 46.61c | 1919.53 ± 16.03ab | 1837.05 ± 54.31bc | 2012.04 ± 16.65a | |
| 6 | 776.92 ± 43.14a | 574.08 ± 14.98b | 649.81 ± 24.70ab | 711.37 ± 19.50a | |
| 8 | 747.19 ± 20.32a | 698.8 ± 101.45a | 843.48 ± 67.52a | 902.88 ± 79.67a | |
| N/P | Initial | 12.07 ± 0.37 | 12.07 ± 0.37 | 12.07 ± 0.37 | 12.07 ± 0.37 |
| 2 | 10.18 ± 0.39ab | 9.29 ± 0.94b | 12.10 ± 0.69ab | 12.93 ± 0.70a | |
| 4 | 22.22 ± 0.62a | 19.94 ± 1.31ab | 18.68 ± 0.9ab | 16.70 ± 0.79b | |
| 6 | 12.34 ± 0.83ab | 9.13 ± 0.54c | 11.05 ± 0.28b | 13.39 ± 0.82a | |
| 8 | 22.20 ± 0.91a | 19.75 ± 1.38ab | 18.05 ± 0.88b | 21.3 ± 0.61ab | |
| Lignin/N | Initial | 39.96 ± 0.76 | 39.96 ± 0.76 | 39.96 ± 0.76 | 39.96 ± 0.76 |
| 2 | 51.90 ± 1.36a | 57.38 ± 1.42a | 52.90 ± 3.98a | 52.31 ± 2.66a | |
| 4 | 71.36 ± 1.41b | 93.34 ± 6.16ab | 85.36 ± 1.6ab | 110.93 ± 4.26a | |
| 6 | 66.93 ± 3.42ab | 72.31 ± 3.26a | 66.06 ± 1.76ab | 59.44 ± 3.98b | |
| 8 | 38.86 ± 0.76b | 42.94 ± 1.99b | 51.73 ± 2.51a | 39.00 ± 0.74b |
References
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The microbial efficiency-matrix stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant in-puts form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Berg, B.; Mcclaugherty, C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Zhou, Y.; Wang, L.F.; Chen, Y.M.; Zhang, J.; Xu, Z.F.; Guo, L.; Wang, L.X.; You, C.M.; Tan, B.; Zhang, L.; et al. Temporal dynamics of mixed litter humification in an alpine treeline ecotone. Sci. Total Environ. 2022, 803, 150122. [Google Scholar] [CrossRef] [PubMed]
- Ponge, J.F.; Jabiol, B.; Gégout, J.C. Geology and climate conditions affect more humus forms than forest canopies at large scale in temperate forests. Geoderma 2011, 162, 187–195. [Google Scholar] [CrossRef]
- Larionova, A.A.; Maltseva, A.N.; Lopes de Gerenyu, V.O.; Kvitkina, A.K.; Bykhovets, S.S.; Zolotareva, B.N.; Kudeyarov, V.N. Effect of temperature and moisture on the mineralization and humification of leaf litter in a model incubation experiment. Eurasian Soil Sci. 2017, 50, 422–431. [Google Scholar] [CrossRef]
- Waez-Mousavi, S.M. Humus systems in the Caspian Hyrcanian temperate forests. Appl. Soil Ecol. 2018, 123, 664–667. [Google Scholar] [CrossRef]
- Liu, Q.; Zhuang, L.Y.; Yin, R.; Ni, X.Y.; You, C.M.; Yue, K.; Tan, B.; Liu, Y.; Zhang, L.; Xu, Z.F. Root diameter controls the accumulation of humic substances in decomposing root litter. Geoderma 2019, 348, 68–75. [Google Scholar] [CrossRef]
- Berg, B. Decomposing litter; limit values; humus accumulation, locally and regionally. Appl. Soil Ecol. 2018, 123, 494–508. [Google Scholar] [CrossRef]
- Ponge, J.F. Plant-soil feedbacks mediated by humus forms: A review. Soil Biol. Biochem. 2013, 57, 1048–1060. [Google Scholar] [CrossRef]
- Feng, J.G.; He, K.Y.; Zhang, Q.F.; Han, M.G.; Zhu, B. Changes in plant inputs alter soil carbon and microbial communities in forest ecosystems. Glob. Chang. Biol. 2022, 28, 3426–3440. [Google Scholar] [CrossRef]
- Muscolo, A.; Bagnato, S.; Sidari, M.; Mercurio, R. A review of the roles of forest canopies gaps. J. For. Res. 2014, 25, 725–736. [Google Scholar] [CrossRef]
- Markiewicz, M.; Hulisz, P.; Charzyński, P.; Piernik, A. Characteristics of soil organic matter of edifisols–An example of techno humus system. Appl. Soil Ecol. 2018, 123, 509–512. [Google Scholar] [CrossRef]
- Ni, X.Y.; Yang, W.Q.; Tan, B.; Li, H.; He, J.; Xu, L.Y.; Wu, F.Z. Forest gaps slow the sequestration of soil organic matter: A humification experiment with six foliar litters in an alpine forest. Sci. Rep. 2016, 6, 19744. [Google Scholar] [CrossRef]
- Tan, Y.; Yang, K.J.; Xu, Z.F.; Zhang, L.; Li, H.; You, C.M.; Tan, B. The Contributions of Soil Fauna to the Accumulation of Humic Substances during Litter Humification in Cold Forests. Forests 2022, 13, 1235. [Google Scholar] [CrossRef]
- De Graaff, M.A.; Classen, A.T.; Castro, H.F.; Schadt, C.W. Labile soil carbon inputs mediate the soil microbial community composition and plant residue decomposition rates. New Phytol. 2010, 188, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Amolikondori, A.; Abrari Vajari, K.; Feizian, M.; Di Iorio, A. Influences of forest gaps on soil physico-chemical and biological properties in an oriental beech (Fagus orientalis L.) stand of Hyrcanian forest, north of Iran. Iforest-Biogeosci. For. 2020, 13, 124. [Google Scholar] [CrossRef]
- Wang, D.Y.; Ni, X.Y.; Guo, H.R.; Dai, W.Y. Alpine Litter Humification and Its Response to Reduced Snow Cover: Can More Carbon Be Sequestered in Soils? Forests 2022, 13, 897. [Google Scholar] [CrossRef]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2017, 2, 17105. [Google Scholar] [CrossRef]
- Bagnato, S.; Marziliano, P.A.; Sidari, M.; Mallamaci, C.; Marra, F.; Muscolo, A. Effects of gap size and cardinal directions on natural regeneration, growth dynamics of trees outside the gaps and soil properties in European beech forests of southern Italy. Forests 2021, 12, 1563. [Google Scholar] [CrossRef]
- Sorouri, B.; Allison, S.D. Microbial extracellular enzyme activity with simulated climate change. Elem. Sci. Anth. 2022, 10, 00076. [Google Scholar] [CrossRef]
- Ni, X.Y.; Yang, W.Q.; Liao, S.; Li, H.; Tan, B.; Yue, K.; Xu, Z.F.; Zhang, L.; Wu, F.Z. Rapid release of labile components limits the accumulation of humic substances in decomposing litter in an alpine forest. Ecosphere 2018, 9, e02434. [Google Scholar] [CrossRef]
- He, W.; Wu, F.Z.; Zhang, D.J.; Yang, W.Q.; Tan, B.; Zhao, Y.Y.; Wu, Q.Q. The effects of forest gaps on cellulose degradation in the foliar litter of two shrub species in an alpine fir forest. Plant Soil 2015, 393, 109–122. [Google Scholar] [CrossRef]
- Wu, A.M.; Yin, R.; Xu, Z.F.; Zhang, L.; You, C.M.; Liu, Y.; Li, H.; Wang, L.X.; Liu, S.N.; Zhang, Y.Q.; et al. Forest gaps slow lignin and cellulose degradation of fir (Abies faxoniana) twig litter in an alpine forest. Geoderma 2022, 424, 116010. [Google Scholar] [CrossRef]
- Ni, X.Y.; Yang, W.Q.; Tan, B.; He, J.; Xu, L.Y.; Li, H.; Wu, F.Z. Accelerated foliar litter humification in forest gaps: Dual feedbacks of carbon sequestration during winter and the growing season in an alpine forest. Geoderma 2015, 24, 136–144. [Google Scholar] [CrossRef]
- Yang, J.P.; Zhang, Y.; Liang, Z.Y.; Yue, K.; Fu, C.K.; Ni, X.Y.; Wu, F.Z. The optical properties in alkali-soluble fractions extracted from newly shed litters in a subalpine forest. J. Soils Sediments 2020, 20, 1276–1284. [Google Scholar] [CrossRef]
- Wang, L.F.; Zhang, J.; He, R.L.; Chen, Y.M.; Yang, L.; Zheng, H.F.; Li, H.J.; Xiao, J.J.; Liu, Y. Impacts of soil fauna on lignin and cellulose degradation in litter decomposition across an alpine forest-tundra ecotone. Eur. J. Soil Biol. 2018, 87, 53–60. [Google Scholar] [CrossRef]
- Bokhorst, S.; Metcalfe, D.B.; Wardle, D.A. Reduction in snow depth negatively affects decomposers but impact on decomposition rates is substrate dependent. Soil Biol. Biochem. 2013, 62, 157–164. [Google Scholar] [CrossRef]
- Thackeray, C.W.; Derksen, C.; Fletcher, C.G.; Hall, A. Snow and climate: Feedbacks, drivers, and indices of change. Curr. Clim. Chang. Rep. 2019, 5, 322–333. [Google Scholar] [CrossRef]
- Prescott, C.E. Litter decomposition: What controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 2010, 101, 133–149. [Google Scholar] [CrossRef]
- Miltner, A.; Bombach, P.; Schmidt-Brücken, B.; Kästner, M. SOM genesis: Microbial biomass as a significant source. Biogeochemistry 2012, 111, 41–55. [Google Scholar] [CrossRef]
- Chen, M.; Zhu, X.; Zhao, C.; Yu, P.; Abulaizi, M.; Jia, H. Rapid microbial community evolution in initial Carex litter decomposition stages in Bayinbuluk alpine wetland during the freeze-thaw period. Ecol. Indic. 2021, 121, 107180. [Google Scholar] [CrossRef]
- Lou, T.; Wang, X.J.; He, K.P.; Yang, Z. Ohotodegradation of humic substance and its effects on the environmental pollutant behavior. J. Ecol. Environ. Sci. 2014, 23, 716–720. [Google Scholar] [CrossRef]
- Senga, Y.; Naruoka, C.; Moriai, S.; Nohara, S. Characterizing the transformation of aquatic humic substances exposed to ultraviolet radiation using excitation-emission matrix fluorescence spectroscopy and PARAFAC. Inland Waters 2018, 8, 505–511. [Google Scholar] [CrossRef]
- Campbell, E.E.; Parton, W.J.; Soong, J.L.; Paustian, K.; Hobbs, N.T.; Cotrufo, M.F. Using litter chemistry controls on microbial processes to partition litter carbon fluxes with the Litter Decomposition and Leaching (LIDEL) model. Soil Biol. Biochem. 2016, 100, 160–174. [Google Scholar] [CrossRef]
- Liu, Q.; Zhuang, L.Y.; Ni, X.Y.; You, C.M.; Yang, W.Q.; Wu, F.Z.; Tan, B.; Yue, K.; Liu, Y.; Xu, Z.F. Nitrogen additions stimulate litter humification in a subtropical forest, southwestern China. Sci. Rep. 2018, 8, 17525. [Google Scholar] [CrossRef] [PubMed]
- Mikutta, R.; Turner, S.; Schippers, A.; Gentsch, N.; Meyer-Stüve, S.; Condron, L.M.; Peltzer, D.A.; Richardson, S.G.; Eger, S.; Hempel, G.; et al. Microbial and abiotic controls on mineral-associated organic matter in soil profiles along an ecosystem gradient. Sci. Rep. 2019, 9, 10294. [Google Scholar] [CrossRef]
- Wickings, K.; Grandy, A.S.; Reed, S.C.; Cleveland, C.C. The origin of litter chemical complexity during decomposition. Ecol. Lett. 2012, 15, 1180–1188. [Google Scholar] [CrossRef]
- Trap, J.; Bureau, F.; Perez, G.; Aubert, M. PLS-regressions highlight litter quality as the major predictor of humus form shift along forest maturation. Soil Biol. Biochem. 2013, 57, 969–971. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Haddix, M.L.; Kroeger, M.E.; Stewart, C.E. The role of plant input physical-chemical properties, and microbial and soil chemical diversity on the formation of particulate and mineral-associated organic matter. Soil Biol. Biochem. 2022, 168, 108648. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Córdova, S.C.; Olk, D.C.; Dietzel, R.N.; Mueller, K.E.; Archontouilis, S.V.; Castellano, M.J. Plant litter quality affects the accumulation rate, composition, and stability of mineral-associated soil organic matter. Soil Biol. Biochem. 2018, 125, 115–124. [Google Scholar] [CrossRef]
- Singh, L.; Thakur, D.; Sharma, M.K.; Chawla, A. Dynamics of leaf litter decomposition in the timberline zone of western Himalaya. Acta Oecol. 2021, 111, 103715. [Google Scholar] [CrossRef]
- Fu, C.K.; Yang, W.Q.; Tan, B.; Xu, Z.F.; Zhang, Y.; Yang, J.P.; Ni, X.Y.; Wu, F.Z. Seasonal dynamics of litterfall in a sub-alpine spruce-fir forest on the eastern Tibetan Plateau: Allometric scaling relationships based on one year of observations. Forests 2017, 8, 314. [Google Scholar] [CrossRef]
- Tan, B.; Zhang, J.; Yang, W.Q.; Yin, R.; Xu, Z.F.; Liu, Y.; Zhang, L.; Li, H.; You, C.M. Forest gaps retard carbon and nutrient release from twig litter in alpine forest ecosystems. Eur. J. For. Res. 2020, 139, 53–65. [Google Scholar] [CrossRef]
- IUSS Working Group. World reference base for soil resources: International soil classification system for naming soils and creating legends for soil maps. In World Soil Resources Reports; IUSS Working Group, FAO: Rome, Italy, 2015. [Google Scholar]
- Ni, X.Y.; Berg, B.; Yang, W.Q.; Li, H.; Liao, S.; Tan, B.; Yue, K.; Xu, Z.F.; Zhang, L.; Wu, F.Z. Formation of forest gaps accelerates C, N and P release from foliar litter during 4 years of decomposition in an alpine forest. Biogeochemistry 2018, 139, 321–335. [Google Scholar] [CrossRef]
- Brokaw, N.V. The definition of treefall gap and its effect on measures of forest dynamics. Biotropica 1982, 14, 158–160. [Google Scholar] [CrossRef]
- Kwiatkowska-Malina, J. Qualitative and quantitative soil organic matter estimation for sustainable soil management. J. Soils Sediments 2018, 18, 2801–2812. [Google Scholar] [CrossRef]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef]
- Ikeya, K.; Sleighter, R.L.; Hatcher, P.G.; Watanabe, A. Characterization of the chemical composition of soil humic acids using Fourier transform ion cyclotron resonance mass spectrometry. Geochim. Cosmochim. Acta 2015, 153, 169–182. [Google Scholar] [CrossRef]
- Ikeya, K.; Watanabe, A. Direct expression of an index for the degree of humification of humic acids using organic carbon concentration. Soil Sci. Plant Nutr. 2003, 49, 47–53. [Google Scholar] [CrossRef]
- Swift, R.S. Organic matter characterization. In Methods of Soil Analysis. Part 3. Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; Soil Science Society of America, Inc.: Madison, WI, USA, 1996. [Google Scholar]
- MacCarthy, P. The principles of humic substances. Soil Sci. 2001, 166, 738–751. [Google Scholar] [CrossRef]
- Lu, R.K. Soil and Agro-Chemical Analytical Methods; Agricultural Science and Technology Press: Beijing, China, 1999. [Google Scholar]
- Vanderbilt, K.L.; White, C.S.; Hopkins, O.; Craig, J.A. Aboveground decomposition in arid environments: Results of a long-term study in central New Mexico. J. Arid Environ. 2008, 72, 696–709. [Google Scholar] [CrossRef]
- Khatami, S.; Deng, Y.; Tien, M.; Hatcher, P.G. Lignin contribution to aliphatic constituents of humic acids through fungal degradation. J. Environ. Qual. 2019, 48, 1565–1570. [Google Scholar] [CrossRef]
- Wang, L.F.; Chen, Y.M.; Zhou, Y.; Zheng, H.F.; Xu, Z.F.; Tan, B.; You, C.M.; Zhang, L.; Li, H.; Guo, L.; et al. Litter chemical traits strongly drove the carbon fractions loss during decomposition across an alpine treeline ecotone. Sci. Total Environ. 2021, 753, 142287. [Google Scholar] [CrossRef] [PubMed]
- Tamura, M.; Tharayil, N. Plant litter chemistry and microbial priming regulate the accrual, composition and stability of soil carbon in invaded ecosystems. New Phytol. 2014, 203, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Baptist, F.; Yoccoz, N.G.; Choler, P. Direct and indirect control by snow cover over decomposition in alpine tundra along a snowmelt gradient. Plant Soil. 2010, 328, 397–410. [Google Scholar] [CrossRef]
- Chertov, O.G.; Komarov, A.S.; Nadporozhskaya, M.A. Analysis of the dynamics of plant residue mineralization and humification in soil. Eurasian Soil Sci. 2007, 40, 140. [Google Scholar] [CrossRef]
- Gavazov, K.; Canarini, A.; Jassey, V.E.; Mills, R.; Richter, A.; Sundqvist, M.K.; Väisänengh, M.; Walkerij, T.W.; Wardlek, D.A.; Dorrepaal, E. Plant-microbial linkages underpin carbon sequestration in contrasting mountain tundra vegetation types. Soil Biol. Biochem. 2022, 165, 108530. [Google Scholar] [CrossRef]
- Hessen, D.O.; Ågren, G.I.; Anderson, T.R.; Elser, J.J.; De Ruiter, P.C. Carbon sequestration in ecosystems: The role of stoichiometry. Ecology 2004, 85, 1179–1192. [Google Scholar] [CrossRef]
- Yu, G.C.; Zhao, H.B.; Chen, J.; Zhang, T.L.; Cai, Z.L.; Zhou, G.Y.; Li, Z.X.; Qiu, Z.J.; Wu, Z.M. Soil microbial community dynamics mediate the priming effects caused by in situ decomposition of fresh plant residues. Sci. Total Environ. 2020, 737, 139708. [Google Scholar] [CrossRef]
- Vives-Peris, V.; de Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef]
- Tian, K.; Kong, X.S.; Yuan, L.H.; Lin, H.; He, Z.H.; Yao, B.; Ji, Y.L.; Yang, J.B.; Sun, S.C.; Tian, X.J. Priming effect of litter mineralization: The role of root exudate depends on its interactions with litter quality and soil condition. Plant Soil. 2019, 440, 457–471. [Google Scholar] [CrossRef]
- Canessa, R.; van den Brink, L.; Saldaña, A.; Rios, R.S.; Hättenschwiler, S.; Mueller, C.W.; Prater, M.I.; Bader, M.Y. Relative effects of climate and litter traits on decomposition change with time, climate and trait variability. J. Ecol. 2021, 109, 447–458. [Google Scholar] [CrossRef]
- Almeida, L.F.; Souza, I.F.; Hurtarte, L.C.; Teixeira, P.P.C.; Inagaki, T.M.; Silva, I.R.; Mueller, C.W. Forest litter constraints on the pathways controlling soil organic matter formation. Soil Biol. Biochem. 2021, 163, 108447. [Google Scholar] [CrossRef]
- Han, M.G.; Tang, M.; Shi, B.K.; Jin, G.Z. Effect of canopies gap size on soil respiration in a mixed broadleaved-Korean pine forest: Evidence from biotic and abiotic factors. Eur. J. Soil Biol. 2019, 99, 103194. [Google Scholar] [CrossRef]
- Ding, Z.; Liu, X.; Gong, L.; Chen, X.; Zhao, J.; Chen, W. Response of litter decomposition and the soil environment to one-year nitrogen addition in a Schrenk spruce forest in the Tianshan Mountains, China. Sci. Rep. 2022, 12, 648. [Google Scholar] [CrossRef]
- Luo, R.; Kuzyakov, Y.; Zhu, B.; Qiang, W.; Zhang, Y.; Pang, X.Y. Phosphorus addition decreases plant lignin but increases microbial necromass contribution to soil organic carbon in a subalpine forest. Glob. Chang. Biol. 2022, 28, 4194–4210. [Google Scholar] [CrossRef]
- Ueno, A.; Shimizu, S.; Tamamura, S.; Okuyama, H.; Naganuma, T.; Kaneko, K. Anaerobic decomposition of humic substances by Clostridium from the deep subsurface. Sci. Rep. 2016, 6, 18990. [Google Scholar] [CrossRef]
- Fontaine, S.; Barot, S.; Barré, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef]
- Grinhut, T.; Hadar, Y.; Chen, Y. Degradation and transformation of humic substances by saprotrophic fungi: Processes and mechanisms. Fungal Biol. Rev. 2007, 21, 179–189. [Google Scholar] [CrossRef]
- Wang, Q.W.; Robson, T.M.; Pieristè, M.; Kenta, T.; Zhou, W.M.; Kurokawa, H. Canopies structure and phenology modulate the impacts of solar radiation on C and N dynamics during litter decomposition in a temperate forest. Sci. Total Environ. 2022, 820, 153185. [Google Scholar] [CrossRef]
- Adair, E.C.; Parton, W.J.; Del Grosso, S.J.; Silver, W.L.; Harmon, M.E.; Hall, S.A.; Burke, I.C.; Hart, S.C. Simple three-pool model accurately describes patterns of long-term litter decomposition in diverse climates. Glob. Chang. Biol. 2008, 14, 2636–2660. [Google Scholar] [CrossRef]
- Chertov, O.; Kuzyakov, Y.; Priputina, I.; Frolov, P.; Shanin, V.; Grabarnik, P. Modelling the Rhizosphere Priming Effect in Combination with Soil Food Webs to Quantify Interaction between Living Plant, Soil Biota and Soil Organic Matter. Plants 2022, 11, 2605. [Google Scholar] [CrossRef] [PubMed]
- Talbot, J.M.; Treseder, K.K. Interactions among lignin, cellulose, and nitrogen drive litter chemistry-decay relationships. Ecology 2012, 93, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, T.; Li, C.; Song, F. Differences in microbial community and metabolites in litter layer of plantation and original Korean Pine forests in North temperate zone. Microorganisms 2020, 8, 2023. [Google Scholar] [CrossRef]
- Phillips, L.A.; Ward, V.; Jones, M.D. Ectomycorrhizal fungi contribute to soil organic matter cycling in sub-boreal forests. ISME J. 2014, 8, 699–713. [Google Scholar] [CrossRef]
- Song, C.H.; Zhang, Y.L.; Xia, X.F.; Qi, H.; Li, M.X.; Pan, H.W.; Xi, B.D. Effect of inoculation with a microbial consortium that degrades organic acids on the composting efficiency of food waste. Microb. Biotechnol. 2018, 11, 1124–1136. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, L.F.; Chen, Y.M.; Zhang, J.; Liu, Y. Litter stoichiometric traits have stronger impact on humification than environment conditions in an alpine treeline ecotone. Plant Soil 2020, 453, 545–556. [Google Scholar] [CrossRef]
- Kögel-Knabner, I. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter. Soil Biol. Biochem. 2002, 34, 139–162. [Google Scholar] [CrossRef]
- Wolfe, E.R.; Ballhorn, D.J. Do Foliar Endophytes Matter in Litter Decomposition? Microorganisms 2020, 8, 446. [Google Scholar] [CrossRef]
- Sun, T.; Mao, Z.; Dong, L.; Hou, L.; Song, Y.; Wang, X. Further evidence for slow decomposition of very fine roots using two methods: Litterbags and intact cores. Plant Soil 2013, 366, 633–646. [Google Scholar] [CrossRef]
- Peng, Y.; Vesterdal, L.; Peñuelas, J.; Peguero, G.; Wu, Q.Q.; Heděnec, P.; Yue, K.; Wu, F.Z. Soil fauna effects on litter decomposition are better predicted by fauna communities within litterbags than by ambient soil fauna communities. Plant Soil. 2023, 1–11. [Google Scholar] [CrossRef]







| Variables | Gap Size (GS) | Sampling Time (T) | GS × ST | ||||||
|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | |
| Accumulation of humic substances | 2 | 4.790 | 0.011 * | 4 | 105.457 | <0.001 ** | 8 | 1.038 | 0.406 |
| Accumulation of humic acid | 2 | 7.772 | 0.001 ** | 4 | 38.379 | <0.001 ** | 8 | 1.460 | 0.202 |
| Accumulation of fulvic acid | 2 | 2.732 | 0.071 | 4 | 198.146 | <0.001 ** | 8 | 2.421 | 0.033 * |
| Humic acid to fulvic acid ratio | 2 | 0.164 | 0.849 | 4 | 55.822 | <0.001 ** | 8 | 1.054 | 0.404 |
| Humification degree | 2 | 3.222 | 0.045 * | 4 | 350.901 | <0.001 ** | 8 | 0.577 | 0.794 |
| Humification rate | 2 | 0.715 | 0.493 | 4 | 176.572 | <0.001 ** | 8 | 2.395 | 0.039 * |
| E4/E6 | 2 | 3.841 | 0.026 * | 4 | 1037.761 | <0.001 ** | 8 | 0.680 | 0.708 |
| ΔlogK | 2 | 6.863 | 0.002 ** | 4 | 700.596 | <0.001 ** | 8 | 2.218 | 0.035 * |
| A600/C | 2 | 4.612 | 0.013 * | 4 | 435.932 | <0.001 ** | 8 | 0.586 | 0.786 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, A.; You, C.; Yin, R.; Xu, Z.; Zhang, L.; Liu, Y.; Li, H.; Wang, L.; Xu, L.; Xu, H.; et al. Forest Gaps Slow the Humification Process of Fir (Abies faxoniana Rehder & E.H.Wilson) Twig Litter during Eight Years of Decomposition in an Alpine Forest. Forests 2023, 14, 868. https://doi.org/10.3390/f14050868
Wu A, You C, Yin R, Xu Z, Zhang L, Liu Y, Li H, Wang L, Xu L, Xu H, et al. Forest Gaps Slow the Humification Process of Fir (Abies faxoniana Rehder & E.H.Wilson) Twig Litter during Eight Years of Decomposition in an Alpine Forest. Forests. 2023; 14(5):868. https://doi.org/10.3390/f14050868
Chicago/Turabian StyleWu, Aomiao, Chengming You, Rui Yin, Zhenfeng Xu, Li Zhang, Yang Liu, Han Li, Lixia Wang, Lin Xu, Hongwei Xu, and et al. 2023. "Forest Gaps Slow the Humification Process of Fir (Abies faxoniana Rehder & E.H.Wilson) Twig Litter during Eight Years of Decomposition in an Alpine Forest" Forests 14, no. 5: 868. https://doi.org/10.3390/f14050868
APA StyleWu, A., You, C., Yin, R., Xu, Z., Zhang, L., Liu, Y., Li, H., Wang, L., Xu, L., Xu, H., Hou, G., Liu, S., & Tan, B. (2023). Forest Gaps Slow the Humification Process of Fir (Abies faxoniana Rehder & E.H.Wilson) Twig Litter during Eight Years of Decomposition in an Alpine Forest. Forests, 14(5), 868. https://doi.org/10.3390/f14050868