

Growth, Xylem Vulnerability to Cavitation and Leaf Cell Response to Dehydration in Tree Seedlings of the Caribbean Dry Forest



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimentation
2.2. Morphometric and Biomass Measurements
2.3. Leaf Gas Exchange
2.4. Wood Density, Stem Water Content and Huber Value
2.5. Pressure–Volume Curves
2.6. Xylem Embolism Vulnerability Curves
2.7. TLP-Based Hydraulic Safety Margin
2.8. Plant Transpiration
2.9. Statistical Analyses
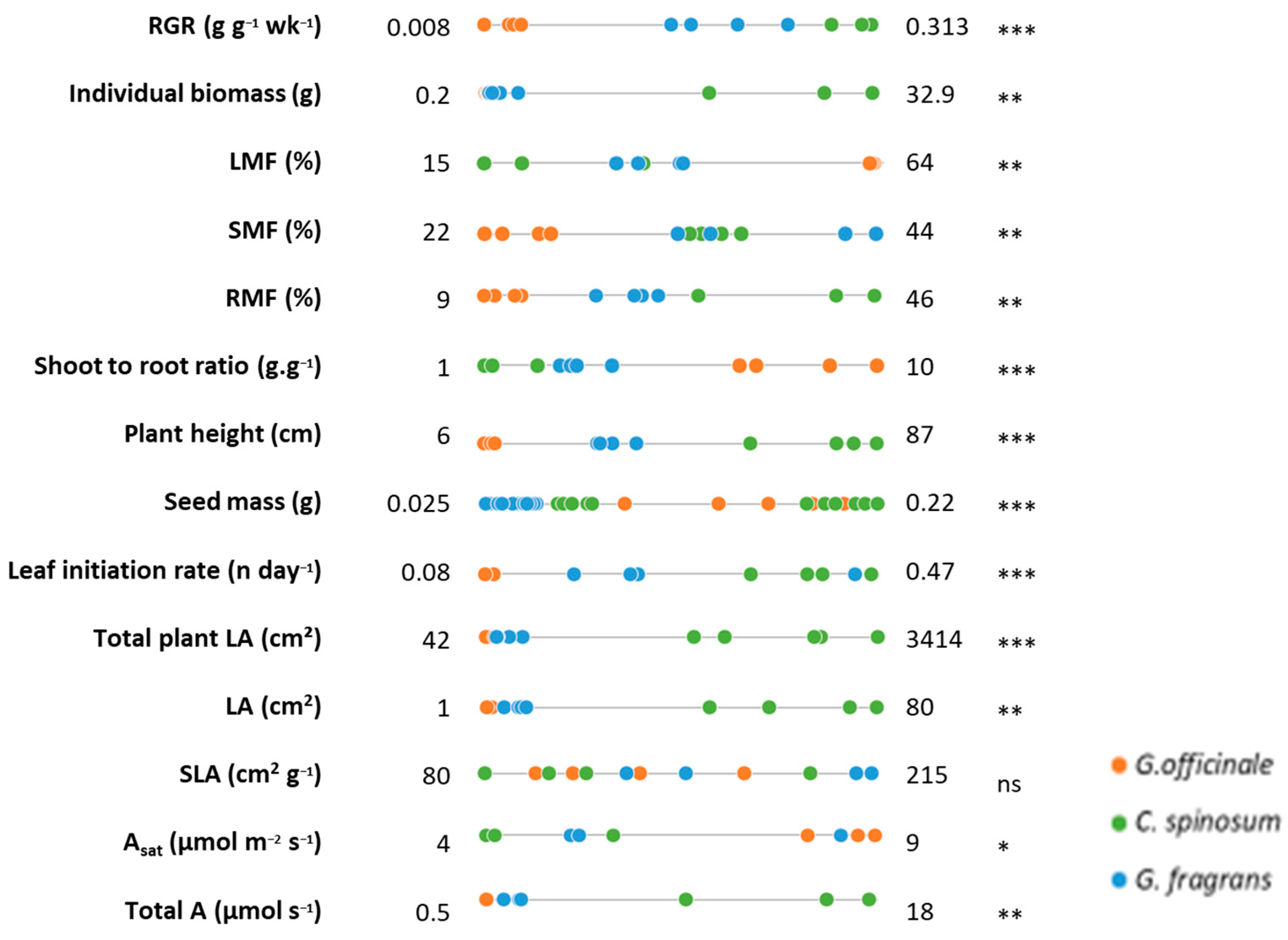
3. Results
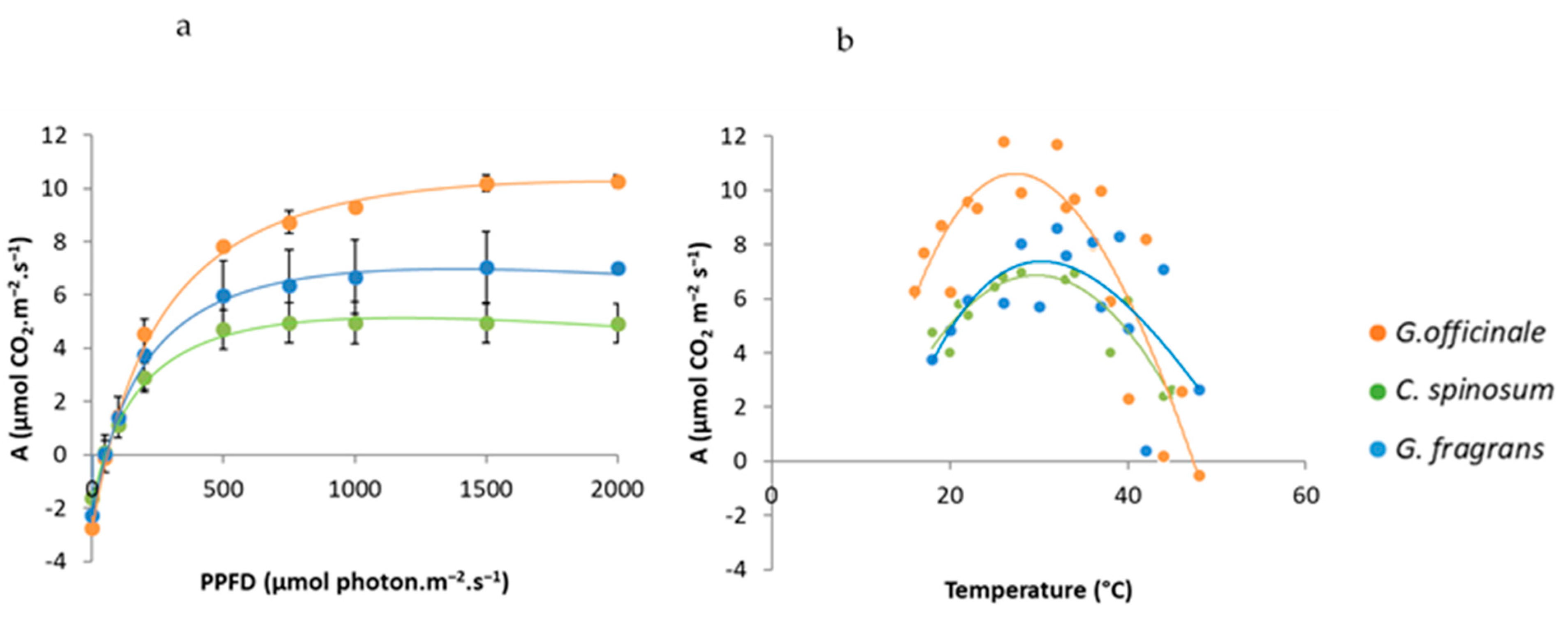
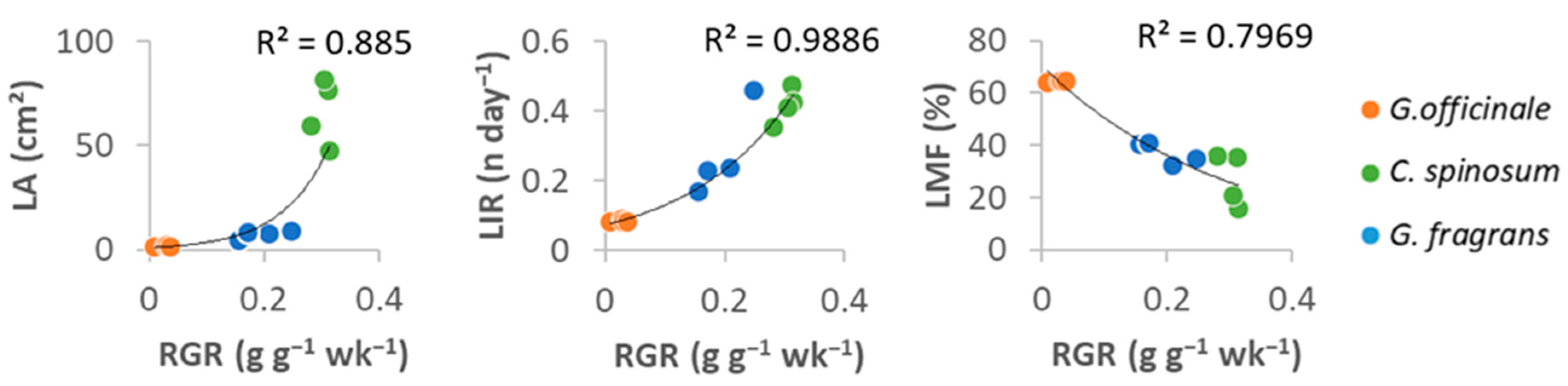
3.1. Growth and Carbon Acquisition
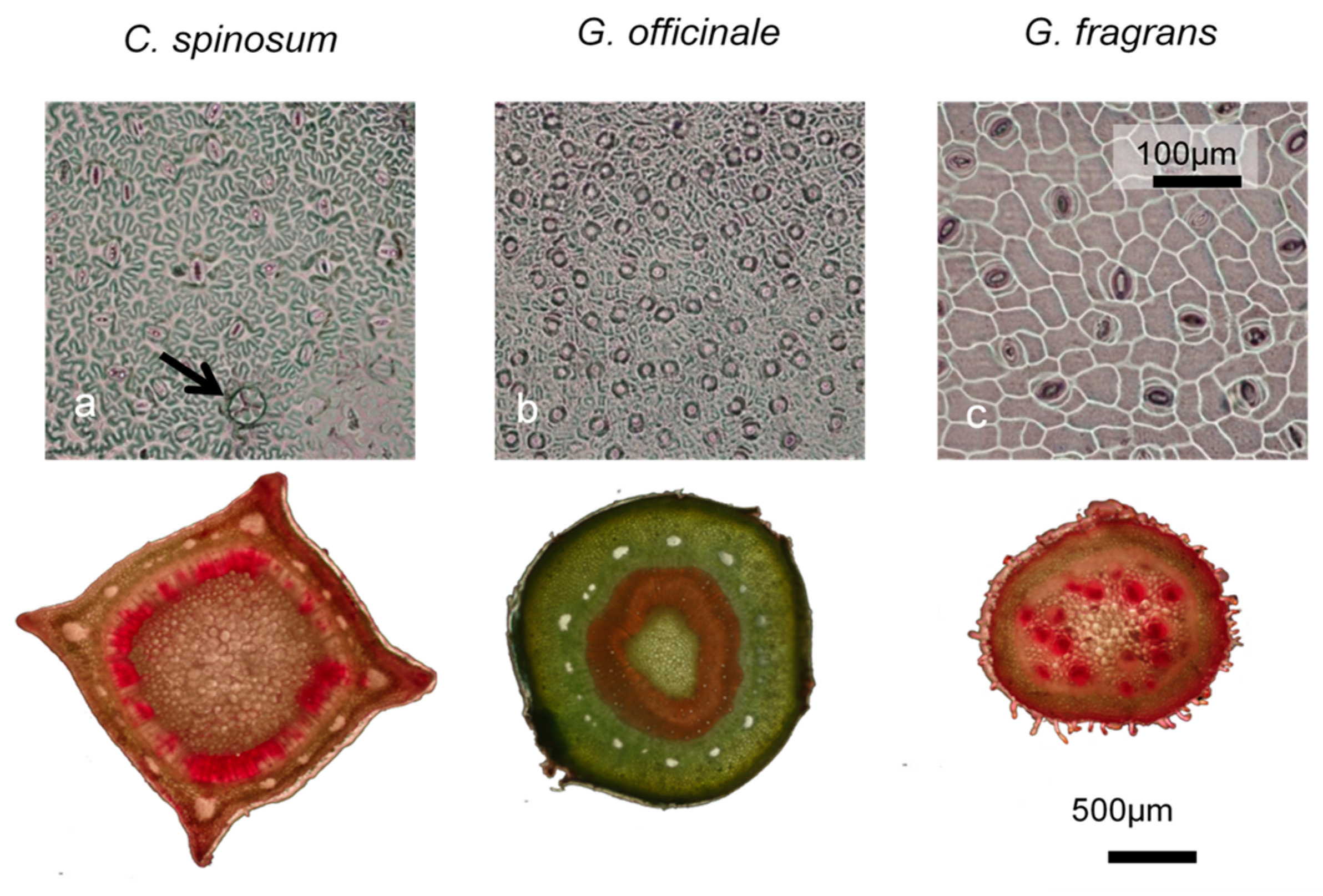
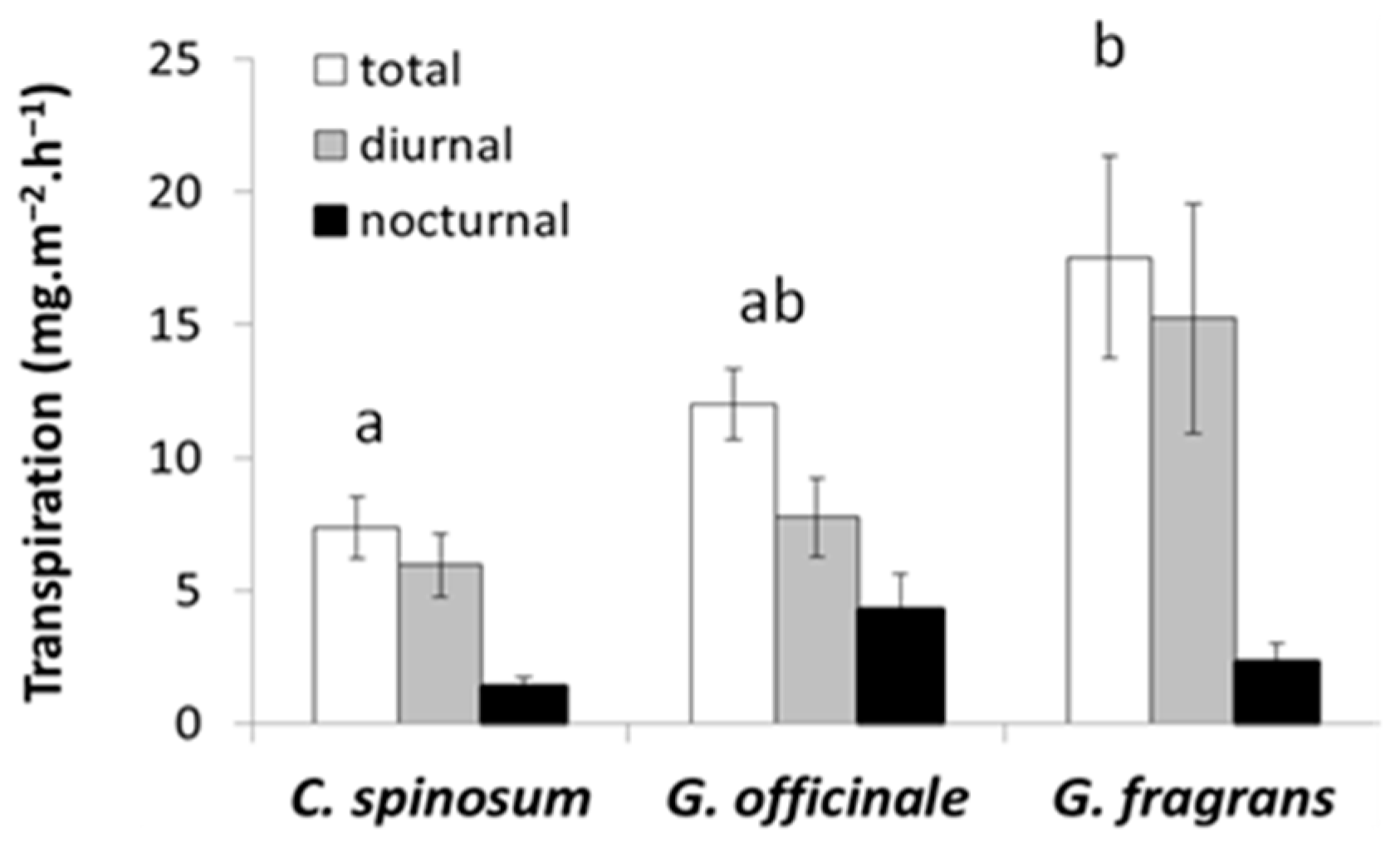
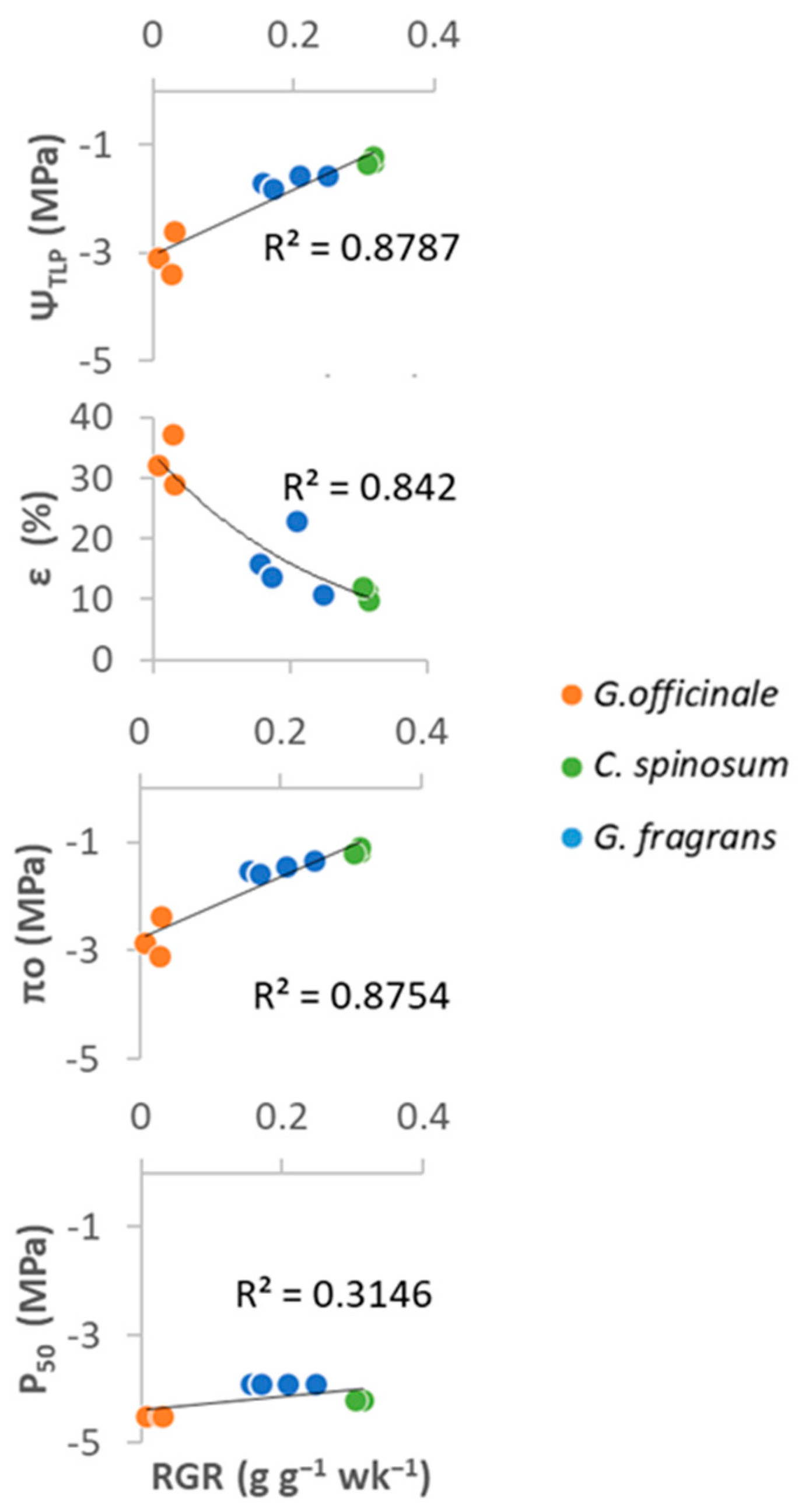
3.2. Water Management
4. Discussion
4.1. Different Growth Strategies in Seedlings of the Caribbean Dry Forest
4.2. Interspecific Differences in Water Use and Drought Tolerance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chazdon, R.L.; Brancalion, P.H.S.; Lamb, D.; Laestadius, L.; Calmon, M.; Kumar, C. A policy-driven knowledge agenda for global forest and landscape restoration. Conserv. Lett. 2017, 10, 125–132. [Google Scholar] [CrossRef]
- Hasnat, G.T.; Hossain, M.K. Global overview of tropical dry forests. In Handbook of Research on the Conservation and Restoration of Tropical Dry Forests; IGI Global: Hershey, PA, USA, 2020; pp. 1–23. [Google Scholar] [CrossRef]
- Dexter, K.G.; Smart, B.; Baldauf, C.; Baker, T.R.; Balinga, M.P.B.; Brienen, R.J.W.; Fauset, S.; Feldpausch, T.R.; Silva, L.F.-D.; Muledi, J.; et al. Floristics and biogeography of vegetation in seasonally dry tropical regions. Int. For. Rev. 2015, 17, 10–32. [Google Scholar] [CrossRef]
- Janzen, D.H. Tropical dry forests. In Biodiversity; National Academy of Sciences/Smithsonian Institution: Washington, DC, USA, 1988; Volume 15, pp. 130–137. [Google Scholar]
- Murphy, P.G.; Lugo, A.E. Ecology of tropical dry forest. Annu. Rev. Ecol. Syst. 1986, 17, 67–88. [Google Scholar] [CrossRef]
- Lamb, D.; Erskine, P.D.; Parrotta, J.A. Restoration of Degraded Tropical Forest Landscapes. Science 2005, 310, 1628–1632. [Google Scholar] [CrossRef]
- Meli, P.; Holl, K.D.; Rey Benayas, J.M.; Jones, H.P.; Jones, P.C.; Montoya, D.; Moreno Mateos, D. A Global Review of Past Land Use, Climate, and Active vs. Passive Restoration Effects on Forest Recovery. PLoS ONE 2017, 12, e0171368. [Google Scholar] [CrossRef]
- UICN. Liste Rouge de la Flore Vasculaire de Guadeloupe; UICN: Paris, France, 2019. [Google Scholar]
- Portillo-Quintero, C.A.; Sánchez-Azofeifa, G.A. Extent and Conservation of Tropical Dry Forests in the Americas. Biol. Conserv. 2010, 143, 144–155. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Okada, K.; Mori, A.S. Reconsidering Biodiversity Hotspots Based on the Rate of Historical Land-Use Change. Biol. Conserv. 2019, 233, 268–275. [Google Scholar] [CrossRef]
- Banda, K.; Delgado, A.; Dexter, K.; Linares, R.; Oliveira, A.; Prado, D.; Pullan, M.; Quintana, C.; Riina, R.; Rodríguez, M.G.M.; et al. Plant diversity patterns in neotropical dry forests and their conservation implications. Science 2016, 353, 1383–1387. [Google Scholar]
- Giorgi, F. Climate Change Hot-Spots. Geophys. Res. Lett. 2006, 33, 101029. [Google Scholar] [CrossRef]
- Allen, K.; Dupuy, J.M.; Gei, M.G.; Hulshof, C.; Medvigy, D.; Pizano, C.; Salgado-Negret, B.; Smith, C.M.; Trierweiler, A.; Van Bloem, S.J.; et al. Will Seasonally Dry Tropical Forests Be Sensitive or Resistant to Future Changes in Rainfall Regimes? Environ. Res. Lett. 2017, 12, 023001. [Google Scholar] [CrossRef]
- Vichot-Llano, A.; Martinez-Castro, D.; Bezanilla-Morlot, A.; Centella-Artola, A.; Giorgi, F. Projected Changes in Precipitation and Temperature Regimes and Extremes over the Caribbean and Central America Using a Multiparameter Ensemble of RegCM4. Int. J. Climatol. 2020, 41, 1328–1350. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global Convergence in the Vulnerability of Forests to Drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef]
- González-M, R.; Posada, J.M.; Carmona, C.P.; Garzón, F.; Salinas, V.; Idárraga-Piedrahita, Á.; Pizano, C.; Avella, A.; López-Camacho, R.; Norden, N.; et al. Diverging Functional Strategies but High Sensitivity to an Extreme Drought in Tropical Dry Forests. Ecol. Lett. 2020, 24, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Poorter, L.; Rozendaal, D.M.A.; Bongers, F.; de Almeida-Cortez, J.S.; Almeyda Zambrano, A.M.; Álvarez, F.S.; Andrade, J.L.; Villa, L.F.A.; Balvanera, P.; Becknell, J.M.; et al. Wet and Dry Tropical Forests Show Opposite Successional Pathways in Wood Density but Converge over Time. Nat. Ecol. Evol. 2019, 3, 928–934. [Google Scholar] [CrossRef]
- Méndez-Alonzo, R.; Paz, H.; Zuluaga, R.C.; Rosell, J.A.; Olson, M.E. Coordinated Evolution of Leaf and Stem Economics in Tropical Dry Forest Trees. Ecology 2012, 93, 2397–2406. [Google Scholar] [CrossRef]
- Hulshof, C.; Martinez-Yrizar, A.; Burquez, A.; Boyle, B. Plant Functional Trait Variation in Tropical Dry Forests: A Review and Synthesis. In Tropical Dry Forests in the Americas; Sanchez-Azofeifa, A., Powers, J.S., Fernandes, G.W., Quesada, M., Eds.; CRC Press: Boca Raton, USA, 2013. [Google Scholar] [CrossRef]
- Khurana, E.K.T.A.; Singh, J.S. Ecology of seed and seedling growth for conservation and restoration of tropical dry forest: A review. Environ. Conserv. 2001, 28, 39–52. [Google Scholar] [CrossRef]
- Poorter, L.; Markesteijn, L. Seedling Traits Determine Drought Tolerance of Tropical Tree Species. Biotropica 2008, 40, 321–331. [Google Scholar] [CrossRef]
- Rojas, A.; Moreno, L.; Melgarejo, L.M.; Rodríguez, M.A. Physiological response of gmelina (Gmelina arborea Roxb.) to hydric conditions of the colombian Caribbean. Agron. Colomb. 2012, 30, 52–58. [Google Scholar]
- Mediavilla, S.; Escudero, A. Stomatal responses to drought of mature trees and seedlings of two co-occurring Mediterranean oaks. For. Ecol. Manag. 2004, 187, 281–294. [Google Scholar] [CrossRef]
- Poorter, L.; Hayashida-Oliver, Y. Effects of Seasonal Drought on Gap and Understorey Seedlings in a Bolivian Moist Forest. J. Trop. Ecol. 2000, 16, 481–498. [Google Scholar] [CrossRef]
- Tyree, M.T.; Engelbrecht, B.M.J.; Vargas, G.; Kursar, T.A. Desiccation Tolerance of Five Tropical Seedlings in Panama. Relationship to a Field Assessment of Drought Performance. Plant Physiol. 2003, 132, 1439–1447. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, K. Tree seedling development in tropical dry abandoned pasture and secondary forest in Costa Rica. J. Veg. Sci. 1993, 4, 95–102. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Zhang, Y.; Yang, J.; Kreidler, N.; Sun, S.-W.; Lin, L.; Hu, Y.-H.; Cao, K.-F.; Sack, L. Drought Tolerance as a Driver of Tropical Forest Assembly: Resolving Spatial Signatures for Multiple Processes. Ecology 2016, 97, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Green, P.T.; Harms, K.E.; Connell, J.H. Nonrandom, Diversifying Processes Are Disproportionately Strong in the Smallest Size Classes of a Tropical Forest. Proc. Natl. Acad. Sci. USA 2014, 111, 18649–18654. [Google Scholar] [CrossRef] [PubMed]
- Akram, M.A.; Zhang, Y.; Wang, X.; Shrestha, N.; Malik, K.; Khan, I.; Ma, W.; Sun, Y.; Li, F.; Ran, J.; et al. Phylogenetic independence in the variations in leaf functional traits among different plant life forms in an arid environment. J. Plant Physiol. 2022, 272, 153671. [Google Scholar] [CrossRef]
- Sobrado, M.A. Cost-Benefit Relationships in Deciduous and Evergreen Leaves of Tropical Dry Forest Species. Funct. Ecol. 1991, 5, 608. [Google Scholar] [CrossRef]
- Eamus, D. Ecophysiological Traits of Deciduous and Evergreen Woody Species in the Seasonally Dry Tropics. Trends Ecol. Evol. 1999, 14, 11–16. [Google Scholar] [CrossRef]
- Gorné, L.D.; Díaz, S.; Minden, V.; Onoda, Y.; Kramer, K.; Muir, C.; Michaletz, S.T.; Lavorel, S.; Sharpe, J.; Jansen, S.; et al. The Acquisitive–Conservative Axis of Leaf Trait Variation Emerges Even in Homogeneous Environments. Ann. Bot. 2020, 129, 709–722. [Google Scholar] [CrossRef]
- Kitajima, K.; Myers, J.A. Seedling ecophysiology: Strategies toward achievement of positive net carbon balance. In Seedling Ecology and Evolution; Allessio Leck, M., Parker, T.V., Simpson, R.L., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 172–188. [Google Scholar]
- Turner, I.M. The Ecology of Trees in the Tropical Rain Forest; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Ribeiro, D.R.; Silva, J.L.A.; do Nascimento, M.T.; Vitória, A.P. Leaf Habits and Their Relationship with Leaf and Wood Traits in Tropical Dry Forests. Trees 2021, 36, 7–24. [Google Scholar] [CrossRef]
- Garkoti, S.C.; Zobel, D.B.; Singh, S.P. Variation in drought response of sal (Shorea robusta) seedlings. Tree Physiol. 2003, 23, 1021–1030. [Google Scholar] [CrossRef]
- Chaturvedi, R.K.; Tripathi, A.; Raghubanshi, A.S.; Singh, J.S. Functional Traits Indicate a Continuum of Tree Drought Strategies across a Soil Water Availability Gradient in a Tropical Dry Forest. For. Ecol. Manag. 2021, 482, 118740. [Google Scholar] [CrossRef]
- Tyree, M.T.; Sperry, J.S. Vulnerability of xylem to cavitation and embolism. Annu. Rev. Plant Phys. Mol. Bio. 1989, 40, 19–38. [Google Scholar] [CrossRef]
- Bartlett, M.K.; Scoffoni, C.; Sack, L. The Determinants of Leaf Turgor Loss Point and Prediction of Drought Tolerance of Species and Biomes: A Global Meta-Analysis. Ecol. Lett. 2012, 15, 393–405. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Klein, T.; Bartlett, M.; Sack, L.; Pellegrini, A.F.A.; Choat, B.; Jansen, S. Meta-Analysis Reveals That Hydraulic Traits Explain Cross-Species Patterns of Drought-Induced Tree Mortality across the Globe. Proc. Natl. Acad. Sci. USA 2016, 113, 5024–5029. [Google Scholar] [CrossRef]
- Cochard, H.; Badel, E.; Herbette, S.; Delzon, S.; Choat, B.; Jansen, S. Methods for Measuring Plant Vulnerability to Cavitation: A Critical Review. J. Exp. Bot. 2013, 64, 4779–4791. [Google Scholar] [CrossRef] [PubMed]
- Maréchaux, I.; Bartlett, M.K.; Sack, L.; Baraloto, C.; Engel, J.; Joetzjer, E.; Chave, J. Drought Tolerance as Predicted by Leaf Water Potential at Turgor Loss Point Varies Strongly across Species within an Amazonian Forest. Funct. Ecol. 2015, 29, 1268–1277. [Google Scholar] [CrossRef]
- Guillemot, J.; Martin-StPaul, N.K.; Bulascoschi, L.; Poorter, L.; Morin, X.; Pinho, B.X.; Maire, G.; Bittencourt, P.; Oliveira, R.S.; Bongers, F.; et al. Small and Slow Is Safe: On the Drought Tolerance of Tropical Tree Species. Glob. Chang. Biol. 2022, 28, 2622–2638. [Google Scholar] [CrossRef]
- Brodribb, T.J.; McAdam, S.A.M. Evolution of the Stomatal Regulation of Plant Water Content. Plant Physiol. 2017, 174, 639–649. [Google Scholar] [CrossRef]
- Sperry, J.S.; Venturas, M.D.; Anderegg, W.R.L.; Mencuccini, M.; Mackay, D.S.; Wang, Y.; Love, D.M. Predicting Stomatal Responses to the Environment from the Optimization of Photosynthetic Gain and Hydraulic Cost. Plant Cell Environ. 2016, 40, 816–830. [Google Scholar] [CrossRef]
- Eller, C.; Barros, F.d.V.; Rowland, L.; Bittencourt, P.; Rowland, L.; Mencuccini, M.; Oliveira, R.S. Xylem Hydraulic Safety and Construction Costs Determine Tropical Tree Growth. Plant Cell Environ. 2018, 41, 548–562. [Google Scholar] [CrossRef]
- Oliveira, R.S.; Eller, C.B.; Barros, F.d.V.; Hirota, M.; Brum, M.; Bittencourt, P. Linking Plant Hydraulics and the Fast–Slow Continuum to Understand Resilience to Drought in Tropical Ecosystems. New Phytol. 2021, 230, 904–923. [Google Scholar] [CrossRef] [PubMed]
- Pineda-García, F.; Paz, H.; Meinzer, F.C. Drought Resistance in Early and Late Secondary Successional Species from a Tropical Dry Forest: The Interplay between Xylem Resistance to Embolism, Sapwood Water Storage and Leaf Shedding. Plant Cell Environ. 2012, 36, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Markesteijn, L.; Poorter, L.; Paz, H.; Sack, L.; Bongers, F. Ecological Differentiation in Xylem Cavitation Resistance Is Associated with Stem and Leaf Structural Traits. Plant Cell Environ. 2010, 34, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Rollet, B. Description des espèces. In Arbres des Petites Antilles Tome 2; O.N.F.: Paris, France, 2010. [Google Scholar]
- Mira, E.; Rousteau, A.; Tournebize, R.; Robert, M.; Evette, A. Evaluating the Suitability of Neotropical Trees and Shrubs for Soil and Water Bioengineering: Survival and Growth of Cuttings from Ten Caribbean Species. Ecol. Eng. 2022, 185, 106808. [Google Scholar] [CrossRef]
- Hunt, R. Plant Growth Curves. The Functional Approach to Plant Growth Analysis; Edward Arnold Ltd.: London, UK, 1982. [Google Scholar]
- de Lobo, F.A.; de Barros, M.P.; Dalmagro, H.J.; Dalmolin, Â.C.; Pereira, W.E.; de Souza, É.C.; Vourlitis, G.L.; Ortíz, C.E. Fitting Net Photosynthetic Light-Response Curves with Microsoft Excel—A Critical Look at the Models. Photosynthetica 2013, 51, 445–456. [Google Scholar] [CrossRef]
- Simpson, W.T. Specific Gravity, Moisture Content, and Density Relationship for Wood; General Technical Report FPL-GTR-76; United States Department of Agriculture: Washington, DC, USA, 1993.
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. Corrigendum to: New Handbook for Standardised Measurement of Plant Functional Traits Worldwide. Aust. J. Bot. 2016, 64, 715. [Google Scholar] [CrossRef]
- Mayr, S.; Cochard, H. A new method for vulnerability analysis of small xylem areas reveals that compression wood of Norway spruce has lower hydraulic safety than opposite wood. Plant Cell Environ. 2003, 26, 1365–1371. [Google Scholar] [CrossRef]
- Schulte, P.J.; Hinckley, T.M. A Comparison of Pressure-Volume Curve Data Analysis Techniques. J. Exp. Bot. 1985, 36, 1590–1602. [Google Scholar] [CrossRef]
- Sperry, J.S.; Donnelly, J.R.; Tyree, M.T. A Method for Measuring Hydraulic Conductivity and Embolism in Xylem. Plant Cell Environ. 1988, 11, 35–40. [Google Scholar] [CrossRef]
- Pammenter, N.W.; Van der Willigen, C.A. Mathematical and Statistical Analysis of the Curves Illustrating Vulnerability of Xylem to Cavitation. Tree Physiol. 1998, 18, 589–593. [Google Scholar] [CrossRef]
- Peters, J.M.R.; López, R.; Nolf, M.; Hutley, L.B.; Wardlaw, T.; Cernusak, L.A.; Choat, B. Living on the Edge: A Continental-scale Assessment of Forest Vulnerability to Drought. Glob. Chang. Biol. 2021, 27, 3620–3641. [Google Scholar] [CrossRef] [PubMed]
- Martin-StPaul, N.; Delzon, S.; Cochard, H. Plant Resistance to Drought Depends on Timely Stomatal Closure. Ecol. Lett. 2017, 20, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; Holbrook, N.M. Stomatal Closure during Leaf Dehydration, Correlation with Other Leaf Physiological Traits. Plant Physiol. 2003, 132, 2166–2173. [Google Scholar] [CrossRef]
- Vargas, G.G.; Brodribb, T.J.; Dupuy, J.M.; González-M, R.; Hulshof, C.M.; Medvigy, D.; Allerton, T.A.P.; Pizano, C.; Salgado-Negret, B.; Schwartz, N.B.; et al. Beyond Leaf Habit: Generalities in Plant Function across 97 Tropical Dry Forest Tree Species. New Phytol. 2021, 232, 148–161. [Google Scholar] [CrossRef]
- Bucci, S.J.; Scholz, F.G.; Goldstein, G.; Meinzer, F.C.; Hinojosa, J.A.; Hoffmann, W.A.; Franco, A.C. Processes preventing nocturnal equilibration between leaf and soil water potential in tropical savanna woody species. Tree Physiol. 2004, 24, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- McDonald, E.P.; Erickson, J.E.; Kruger, E.L. Research Note: Can Decreased Transpiration Limit Plant Nitrogen Acquisition in Elevated CO2? Funct. Plant Biol. 2002, 29, 1115. [Google Scholar] [CrossRef]
- Gates, D.M. Transpiration and leaf temperature. Annu. Rev. Plant Physiol. 1968, 19, 211–238. [Google Scholar] [CrossRef]
- Clifford, S.C.; Arndt, S.K.; Corlett, J.E.; Joshi, S.; Sankhla, N.; Popp, M.; Jones, H.G. The Role of Solute Accumulation, Osmotic Adjustment and Changes in Cell Wall Elasticity in Drought Tolerance in Ziziphus Mauritiana (Lamk.). J. Exp. Bot. 1998, 49, 967–977. [Google Scholar] [CrossRef]
- Stratton, L.; Goldstein, G.; Meinzer, F.C. Stem Water Storage Capacity and Efficiency of Water Transport: Their Functional Significance in a Hawaiian Dry Forest. Plant Cell Environ. 2000, 23, 99–106. [Google Scholar] [CrossRef]
- Aasamaa, K.; Sõber, A.; Rahi, M. Leaf Anatomical Characteristics Associated with Shoot Hydraulic Conductance, Stomatal Conductance and Stomatal Sensitivity to Changes of Leaf Water Status in Temperate Deciduous Trees. Funct. Plant Biol. 2001, 28, 765. [Google Scholar] [CrossRef]
- Brum, M.; Vadeboncoeur, M.A.; Ivanov, V.; Asbjornsen, H.; Saleska, S.; Alves, L.F.; Penha, D.; Dias, J.D.; Aragão, L.E.O.C.; Barros, F.; et al. Hydrological Niche Segregation Defines Forest Structure and Drought Tolerance Strategies in a Seasonal Amazon Forest. J. Ecol. 2018, 107, 318–333. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Name | Unit |
|---|---|---|
| Atot | Total carbon assimilation rate at the scale of the whole plant | µmol s−1 |
| Asat | Light-saturated rate of carbon assimilation | mmol m−2 s−1 |
| Ds | Stomatal density | n m−2 |
| gs | Stomatal conductance | mmol m−2 s−1 |
| Hv | Huber value (sapwood area/leaf area) | (×10−4) cm−2 cm−2 |
| HSM | Hydraulic safety margin | MPa |
| LA | Leaf area unit | cm2 |
| LIR | Leaf initiation rate | n day−1 |
| LMF | Leaf mass fraction (leaf mass/plant mass) | % |
| P50 | Pressure causing 50% loss of conductance | MPa |
| PLC | Percent loss of hydraulic conductivity | % |
| PPFD | Photosynthetic photon flux density | µmol (photon) m−2 s−1 |
| RGR | Relative growth rate | g g−1 wk−1 |
| RMF | Root mass fraction (root mass/plant mass) | % |
| s | Slope of the vulnerability curve | % MPa−1 |
| SLA | Specific leaf area (leaf area/leaf mass) | cm2 g−1 |
| SMF | Stem mass fraction (stem mass/plant mass) | % |
| SWC | Stem water content | % |
| SWD | Stem wood density | g cm−3 |
| ε | Bulk modulus of elasticity | MPa |
| πo | Osmotic potential at full turgor | MPa |
| Ψ | Leaf water potential | MPa |
| ΨTLP | Leaf water potential at the turgor loss point | MPa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mira, E.; Cochard, H.; Evette, A.; Dulormne, M. Growth, Xylem Vulnerability to Cavitation and Leaf Cell Response to Dehydration in Tree Seedlings of the Caribbean Dry Forest. Forests 2023, 14, 697. https://doi.org/10.3390/f14040697
Mira E, Cochard H, Evette A, Dulormne M. Growth, Xylem Vulnerability to Cavitation and Leaf Cell Response to Dehydration in Tree Seedlings of the Caribbean Dry Forest. Forests. 2023; 14(4):697. https://doi.org/10.3390/f14040697
Chicago/Turabian StyleMira, Eléonore, Hervé Cochard, André Evette, and Maguy Dulormne. 2023. "Growth, Xylem Vulnerability to Cavitation and Leaf Cell Response to Dehydration in Tree Seedlings of the Caribbean Dry Forest" Forests 14, no. 4: 697. https://doi.org/10.3390/f14040697
APA StyleMira, E., Cochard, H., Evette, A., & Dulormne, M. (2023). Growth, Xylem Vulnerability to Cavitation and Leaf Cell Response to Dehydration in Tree Seedlings of the Caribbean Dry Forest. Forests, 14(4), 697. https://doi.org/10.3390/f14040697