

Efficient Virus-Induced Gene Silencing in Ilex dabieshanensis Using Tobacco Rattle Virus

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growing Conditions
2.2. Sequence Analysis of IdChlH
2.3. pTRV2-IdChlH Vector Construction
2.4. Agroinfiltration of the Ilex Plant
2.5. Expression Analysis by qRT-PCR
2.6. Statistical Analysis
3. Results
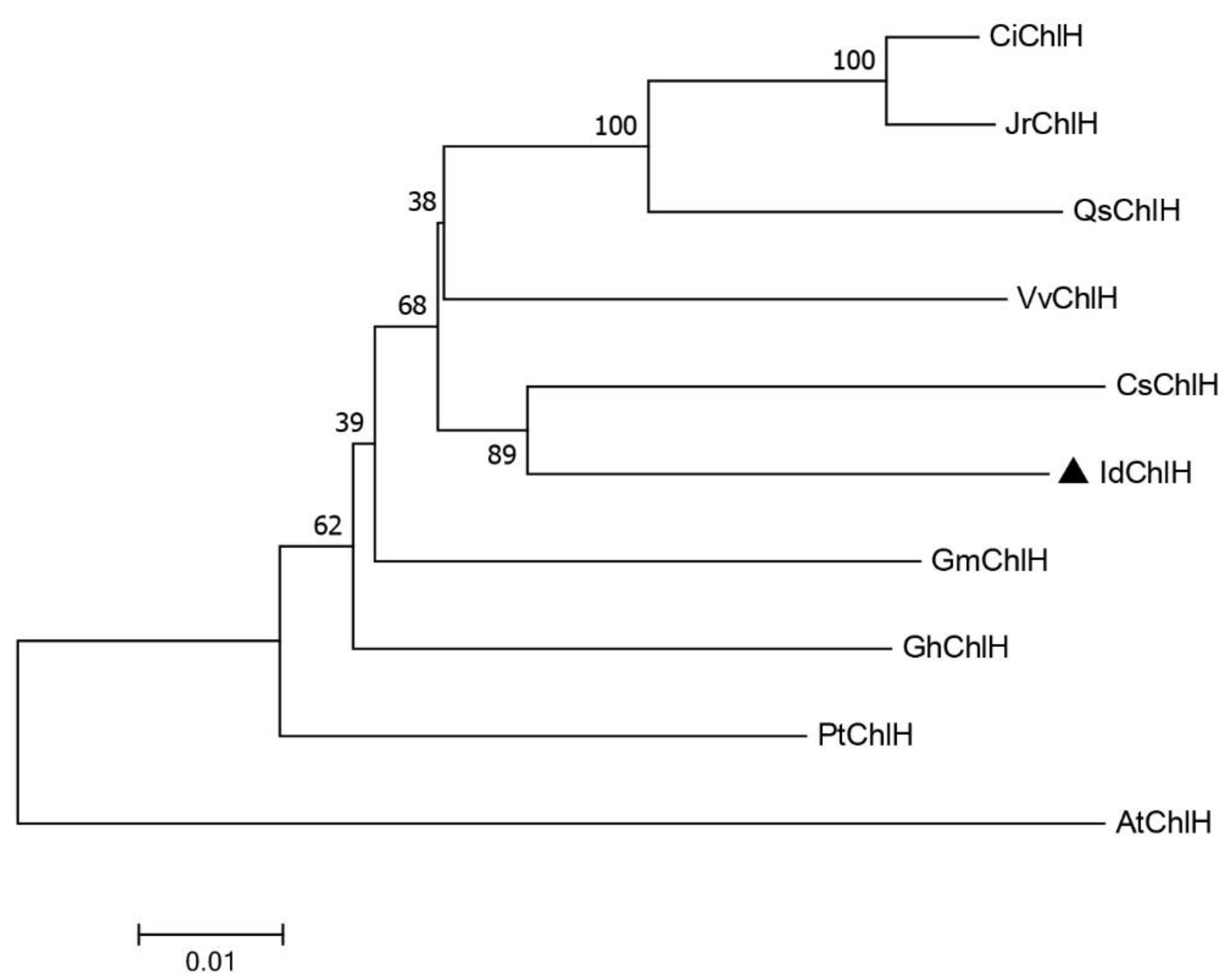
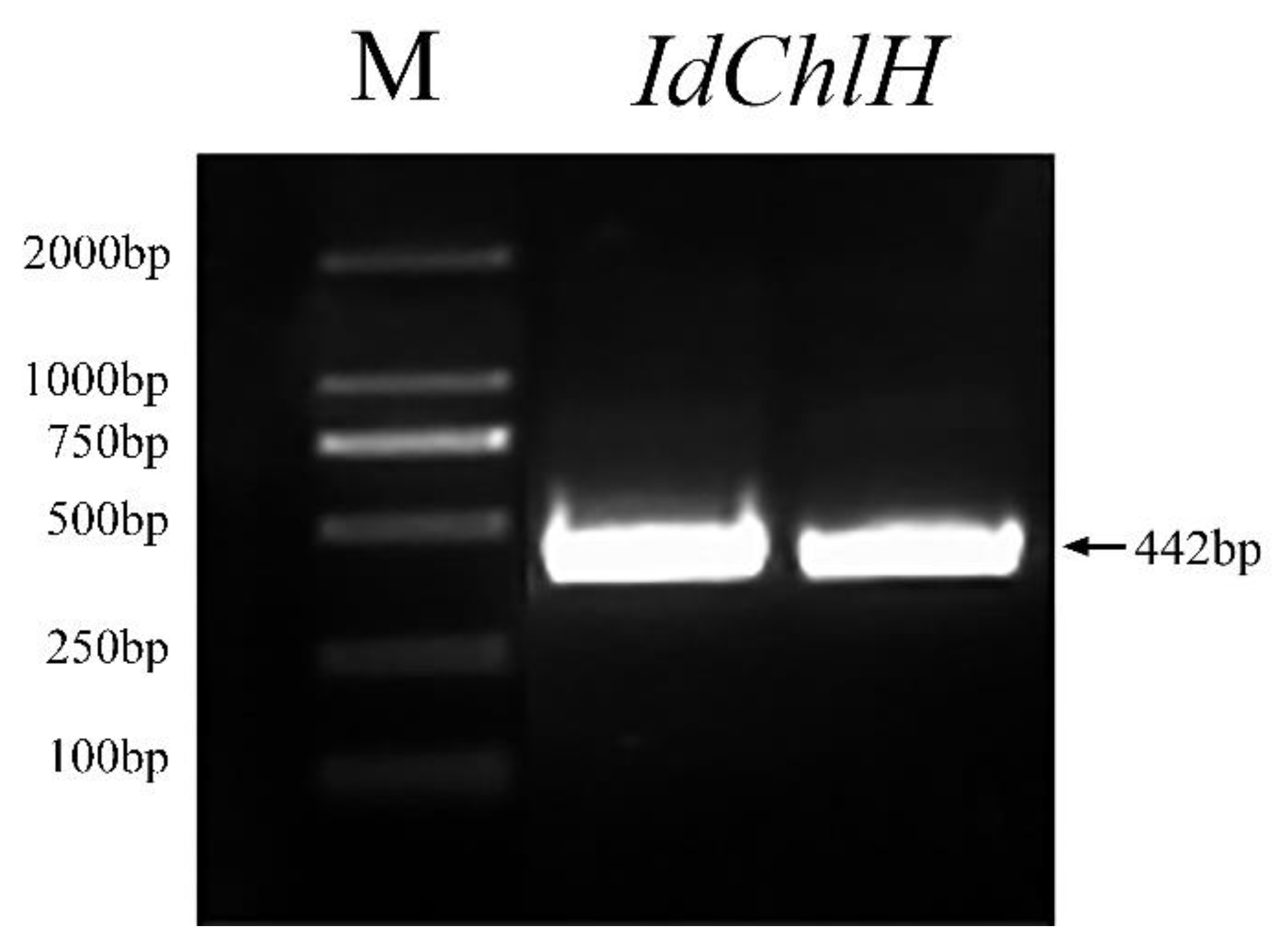
3.1. IdChlH Sequence Characteristics
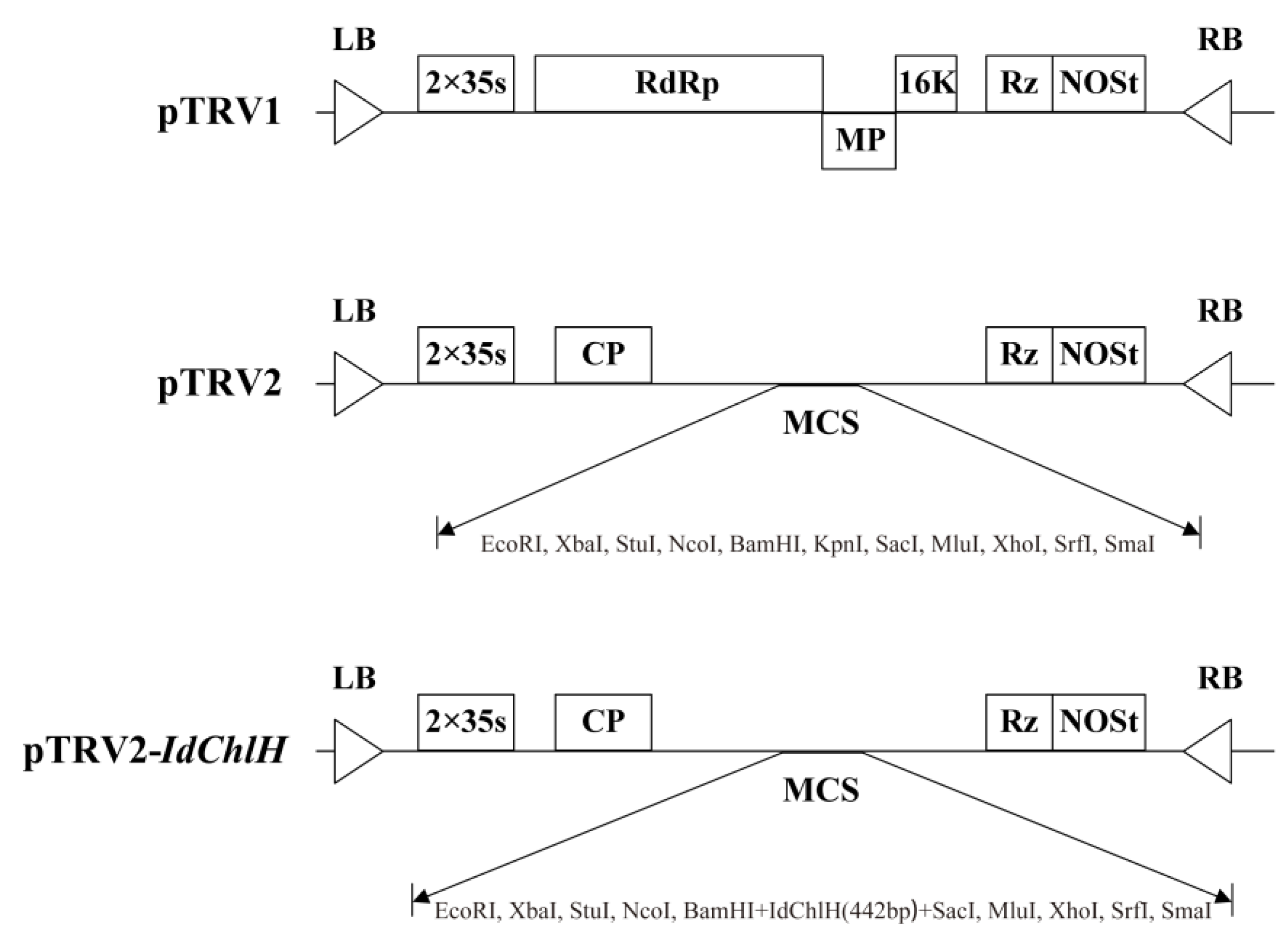
3.2. Construction of the pTRV2-IdChlH Vector
3.3. VIGS-Mediated Silencing of the IdChlH Gene in I. dabieshanensis
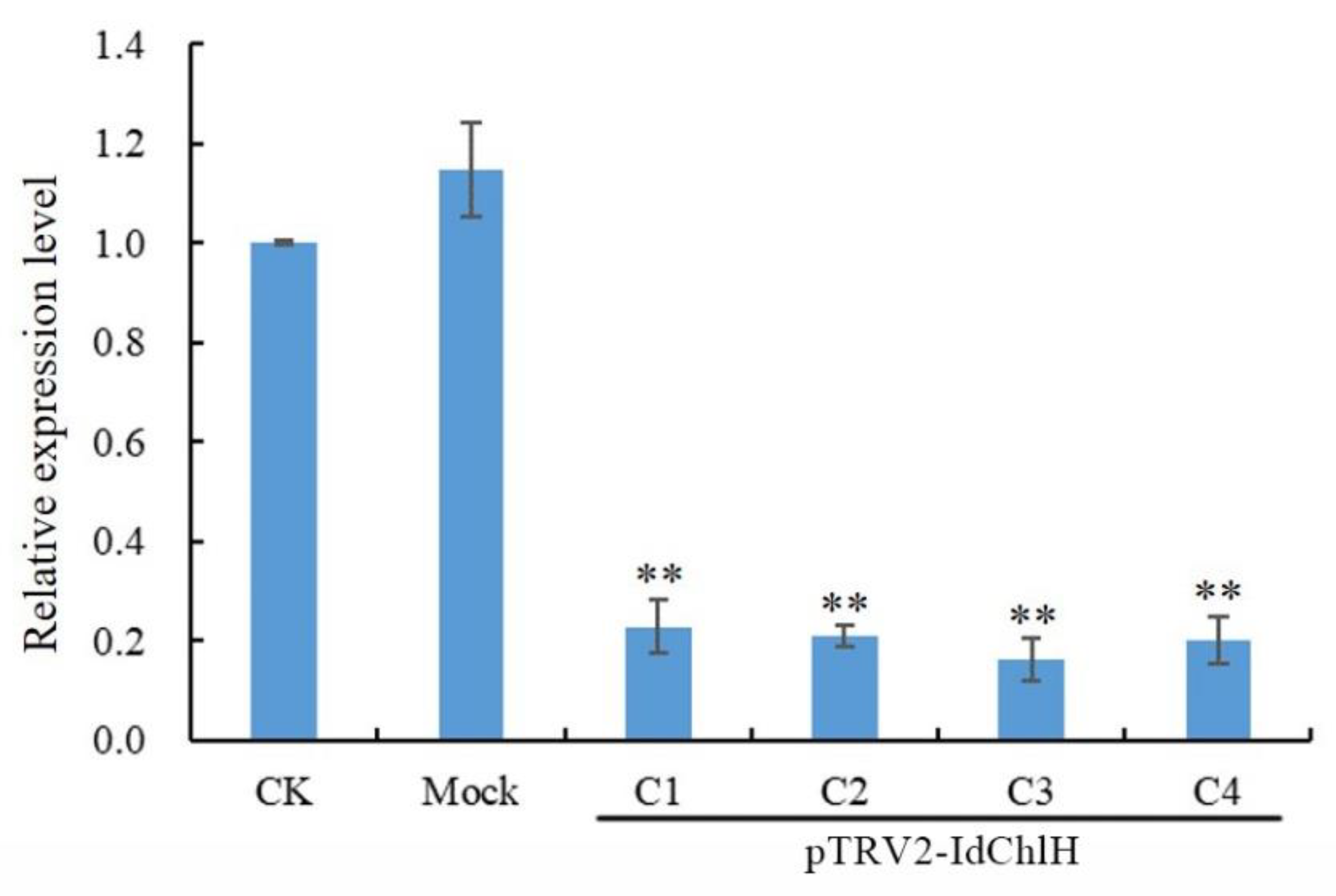
3.4. qRT-PCR Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Su, T.; Zhang, M.; Shan, Z.; Li, X.; Han, M. Comparative survey of morphological variations and plastid genome sequencing reveals phylogenetic divergence between four endemic Ilex species. Forests 2020, 11, 964. [Google Scholar] [CrossRef]
- Chong, X.; Li, Y.; Yan, M.; Wang, Y.; Li, M.; Zhou, Y.; Chen, H.; Lu, X.; Zhang, F. Comparative chloroplast genome analysis of 10 Ilex species and the development of species-specific identification markers. Ind. Crops Prod. 2022, 187, 115408. [Google Scholar] [CrossRef]
- Yao, X.; Zhang, F.; Corlett, R.T. Utilization of the Hollies (Ilex L. spp.): A Review. Forests 2022, 13, 94. [Google Scholar] [CrossRef]
- Manen, J.; Barriera, G.; Loizeau, P.; Naciri, Y. The history of extant Ilex species (Aquifoliaceae): Evidence of hybridization within a Miocene radiation. Mol. Phylogenet. Evol. 2010, 57, 961–977. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Song, Y.; Yang, J.B.; Tan, Y.H.; Corlett, R.T. Phylogeny and biogeography of the hollies (Ilex L., Aquifoliaceae). J. Syst. Evol. 2020, 59, 73–82. [Google Scholar] [CrossRef]
- Shi, L.; Li, N.; Wang, S.; Zhou, Y.; Huang, W.; Yang, Y.; Ma, Y.; Zhou, R. Molecular evidence for the hybrid origin of Ilex dabieshanensis (Aquifoliaceae). PloS ONE 2016, 11, e0147825. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Ning, K.; Mo, Z.; Zhang, F.; Zhou, Y.; Chong, X.; Zhang, D.; El-Kassaby, Y.; Bian, J.; Chen, H. Complete chloroplast genome of Ilex dabieshanensis: Genome structure, comparative analyses with three traditional Ilex tea species, and its phylogenetic relationships within the family Aquifoliaceae. PloS ONE 2022, 17, e0268679. [Google Scholar] [CrossRef]
- Burch-Smith, T.; Anderson, J.; Martin, G.; Dinesh-Kumar, S. Applications and advantages of virus-induced gene silencing for gene function studies in plants. Plant J. 2004, 39, 734–746. [Google Scholar] [CrossRef]
- Baulcombe, D. Fast forward genetics based on virus-induced gene silencing. Curr. Opin. Plant Biol. 1999, 2, 109–113. [Google Scholar] [CrossRef]
- Zhou, P.; Peng, J.; Zeng, M.; Wu, L.; Fan, Y.; Zeng, L. Virus-induced gene silencing (VIGS) in Chinese narcissus and its use in functional analysis of NtMYB3. Hortic. Plant J. 2021, 7, 565–572. [Google Scholar] [CrossRef]
- Dommes, A.; Gross, T.; Herbert, D.; Kivivirta, K.; Becker, A. Virus-induced gene silencing: Empowering genetics in non-model organisms. J. Exp. Bot. 2019, 70, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Senthil-Kumar, M.; Mysore, K. New dimensions for VIGS in plant functional genomics. Trends Plant Sci. 2011, 16, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Tuo, D.; Yan, P.; Zhao, G.; Cui, H.; Zhu, G.; Liu, Y.; Yang, X.; Wang, H.; Li, X.; Shen, W.; et al. An efficient papaya leaf distortion mosaic potyvirus vector for virus-induced gene silencing in papaya. Hortic. Res. 2021, 8, 144. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, R.; Shafiq, M.; Mansoor, S.; Briddon, R.; Scheffler, B.; Scheffler, J.; Amin, I. Virus-induced gene silencing in cultivated cotton (Gossypium spp.) using tobacco rattle virus. Mol. Biotechnol. 2016, 58, 65–72. [Google Scholar] [CrossRef]
- Ratcliff, F.; Martin-Hernandez, A.; Baulcombe, D. Tobacco rattle virus as a vector for analysis of gene function by silencing. Plant J. 2001, 25, 237–245. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Dinesh-Kumar, S. Virus-induced gene silencing in tomato. Plant J. 2002, 31, 777–786. [Google Scholar] [CrossRef]
- Andres, R.; Coneva, V.; Frank, M.; Tuttle, J.; Samayoa, L.; Han, S.; Kaur, B.; Zhu, L.; Fang, H.; Bowman, D.; et al. Modifications to a LATE MERISTEM IDENTITY1 gene are responsible for the major leaf shapes of Upland cotton (Gossypium hirsutum L.). Proc. Natl. Acad. Sci. USA 2017, 114, E57–E66. [Google Scholar] [CrossRef]
- Burch-Smith, T.; Schiff, M.; Liu, Y.; Dinesh-Kumar, S. Efficient virus-induced gene silencing in Arabidopsis. Plant Physiol. 2006, 142, 21–27. [Google Scholar] [CrossRef]
- Yan, H.; Shi, S.; Ma, N.; Cao, X.; Zhang, H.; Qiu, X.; Wang, Q.; Jian, H.; Zhou, N.; Zhang, Z.; et al. Graft-accelerated virus-induced gene silencing facilitates functional genomics in rose flowers. J. Integr. Plant Biol. 2018, 60, 34–44. [Google Scholar] [CrossRef]
- Chen, J.; Jiang, C.; Gookin, T.; Hunter, D.; Clark, D.; Reid, M. Chalcone synthase as a reporter in virus-induced gene silencing studies of flower senescence. Plant Mol. Biol. 2004, 55, 521–530. [Google Scholar] [CrossRef]
- Yan, H.; Fu, D.; Zhu, B.; Liu, H.; Shen, X.; Luo, Y. Sprout vacuum-infiltration: A simple and efficient agroinoculation method for virus-induced gene silencing in diverse solanaceous species. Plant Cell Rep. 2012, 31, 1713–1722. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Höfgen, R.; Willmitzer, L. Storage of competent cells for Agrobacterium transformation. Nucleic Acids Res. 1988, 16, 9877. [Google Scholar] [CrossRef]
- Xu, H.; Xu, L.; Yang, P.; Cao, Y.; Tang, Y.; He, G.; Yuan, S.; Ming, J. Tobacco rattle virus-induced PHYTOENE DESATURASE (PDS) and Mg-chelatase H subunit (ChlH) gene silencing in Solanum pseudocapsicum L. PeerJ 2018, 6, e4424. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Scofield, S.; Huang, L.; Brandt, A.; Gill, B. Development of a virus-induced gene-silencing system for hexaploid wheat and its use in functional analysis of the Lr21-mediated leaf rust resistance pathway. Plant Physiol. 2005, 138, 2165–2173. [Google Scholar] [CrossRef] [PubMed]
- Kurth, E.; Peremyslov, V.; Prokhnevsky, A.; Kasschau, K.; Miller, M.; Carrington, J.; Dolja, V. Virus-derived gene expression and RNA interference vector for grapevine. J. Virol. 2012, 86, 6002–6009. [Google Scholar] [CrossRef]
- Jiang, Y.; Ye, S.; Wang, L.; Duan, Y.; Lu, W.; Liu, H.; Fan, D.; Zhang, F.; Luo, K. Heterologous gene silencing induced by tobacco rattle virus (TRV) is efficient for pursuing functional genomics studies in woody plants. Plant Cell Tiss. Organ Cult. 2014, 116, 163–174. [Google Scholar] [CrossRef]
- Sasaki, S.; Yamagishi, N.; Yoshikawa, N. Efficient virus-induced gene silencing in apple, pear and Japanese pear using Apple latent spherical virus vectors. Plant Methods 2011, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D. VIGS vectors for gene silencing: Many targets, many tools. Annu. Rev. Plant Biol. 2004, 55, 495–519. [Google Scholar] [CrossRef]
- Naylor, M.; Reeves, J.; Cooper, J.; Edwards, M.; Wang, H. Construction and properties of a gene-silencing vector based on Poplar mosaic virus (genus Carlavirus). J. Virol. Methods 2005, 124, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Lansac, M.; Eyquard, J.; Salvador, B.; Garcia, J.; Le Gall, O.; Decroocq, V.; Schurdi-Levraud Escalettes, V. Application of GFP-tagged Plum pox virus to study Prunus-PPV interactions at the whole plant and cellular levels. J. Virol. Methods 2005, 129, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Sun, J.; Yao, J.; Wang, S.; Ding, M.; Zhang, H.; Qian, Z.; Zhao, N.; Sa, G.; Zhao, R.; et al. High rates of virus-induced gene silencing by tobacco rattle virus in Populus. Tree Physiol. 2015, 35, 1016–1029. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fu, D.; Zhu, B.; Yan, H.; Shen, X.; Zuo, J.; Zhu, Y.; Luo, Y. Virus-induced gene silencing in eggplant (Solanum melongena). J. Integr. Plant Biol. 2012, 54, 422–429. [Google Scholar] [CrossRef]
- Grimsley, N.; Hohn, B.; Hohn, T.; Walden, R. “Agroinfection,” an alternative route for viral infection of plants by using the Ti plasmid. Proc. Natl. Acad. Sci. USA 1986, 83, 3282–3286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Shi, J.-A.; Chen, J.-B.; Li, D.-D.; Li, J.-J.; Guo, H.-L.; Zong, J.-Q.; Wang, Y.; Guo, A.-G.; Liu, J.-X. Efficient virus-induced gene silencing in Cynodon dactylon and Zoysia japonica using rice tungro bacilliform virus vectors. Sci. Hortic. 2016, 207, 97–103. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Prime Name | Primer Sequence (5′ to 3′) |
|---|---|
| IdChlH-F | GAAGGCCTCCATGGGGATCCGCGCTGGATCCACAATCTATTC |
| IdChlH-R | CGAGACGCGTGAGCTCCTCTAACCTCAACGCCAAGCGC |
| IdChlH-qd-F | TTCTTCGTGGAATGATGAGA |
| IdChlH-qd-R | CCGTGAGTGAGATTTCTGA |
| Actin-F | CCACCTACAACTCCATCAT |
| Actin-R | TTCCTTGCTCATACGATCA |
| Table | Number of Treated Plants | Silencing Efficiency a |
|---|---|---|
| pTRV2-IdChlH | 46 | 39/46 (84.8%) |
| Mock | 9 | 0/9 (0%) |
| Control | 9 | 0/9 (0%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chong, X.; Wang, Y.; Xu, X.; Zhang, F.; Wang, C.; Zhou, Y.; Zhou, T.; Li, Y.; Lu, X.; Chen, H. Efficient Virus-Induced Gene Silencing in Ilex dabieshanensis Using Tobacco Rattle Virus. Forests 2023, 14, 488. https://doi.org/10.3390/f14030488
Chong X, Wang Y, Xu X, Zhang F, Wang C, Zhou Y, Zhou T, Li Y, Lu X, Chen H. Efficient Virus-Induced Gene Silencing in Ilex dabieshanensis Using Tobacco Rattle Virus. Forests. 2023; 14(3):488. https://doi.org/10.3390/f14030488
Chicago/Turabian StyleChong, Xinran, Yue Wang, Xiaoyang Xu, Fan Zhang, Chuanyong Wang, Yanwei Zhou, Ting Zhou, Yunlong Li, Xiaoqing Lu, and Hong Chen. 2023. "Efficient Virus-Induced Gene Silencing in Ilex dabieshanensis Using Tobacco Rattle Virus" Forests 14, no. 3: 488. https://doi.org/10.3390/f14030488
APA StyleChong, X., Wang, Y., Xu, X., Zhang, F., Wang, C., Zhou, Y., Zhou, T., Li, Y., Lu, X., & Chen, H. (2023). Efficient Virus-Induced Gene Silencing in Ilex dabieshanensis Using Tobacco Rattle Virus. Forests, 14(3), 488. https://doi.org/10.3390/f14030488