1. Introduction
Originating from single-celled prokaryotes in the ocean approximately 2.5 billion years ago, photosynthesis is a natural process in plants consisting of a series of well-coordinated biochemical reactions that use sunlight energy to reduce atmospheric CO
2 into energy-rich carbohydrates in chloroplasts. In the stroma of a chloroplast, CO
2 is fixed into carbohydrates via the Calvin-Benson cycle utilizing the ATP and NADPH produced by photosynthesis in the thylakoid membrane. The first step of the Calvin-Benson cycle is catalyzed by the enzyme ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO) which, as the name suggests, can accept either CO
2 or O
2 as substrates to carry out either a carboxylation or oxygenation reaction, respectively. The carboxylation reaction leads to carbon fixation while the oxygenation reaction leads to photorespiration. The fixed CO
2 molecule moves on to anabolic pathways for sucrose and starch biosynthesis. The oxygenation product phosphoglycolate must be converted in a series of reactions that consume energy and CO
2, eventually generating one molecule of 3-phosphoglycerate [
1]. This energy consumption and the loss of previously fixed CO
2 have made the photorespiration pathway a target for improving photosynthetic efficiency in C3 plants.
Plants grown at elevated CO
2 concentrations, that is, with low RuBisCO oxygenation activity, have been shown to have increased productivity [
2]. Such results have boosted the belief that engineering the RuBisCO enzyme for increasing substrate specificity toward CO
2 could substantially improve photosynthetic efficiency leading to higher productivity [
3,
4,
5]. However, enzyme engineering approaches to improve RuBisCO catalysis by random or site-directed mutagenesis or directed evolution have largely failed to yield substantial kinetic enhancements [
4,
6,
7,
8]. The lack of considerable improvements from these studies is potentially due to the existence of a trade-off between CO
2 specificity and carboxylation velocity—such that carboxylation activity decreases when CO
2 specificity is increased [
9,
10]. However, no statistically significant relationship between carboxylase efficiencies and CO
2 specificities of RuBisCO were found between 11 diatom species [
11]. A computational modeling analysis of RuBisCO kinetics also challenged the trade-off idea [
12]. It remains a possibility that RuBisCO can be improved via enzyme engineering with a better designed screening system. For instance, the successful expression of a functional simplified α-carboxysome from cyanobacteria in tobacco chloroplasts led to an increase in biomass production [
13]. Other attempts have aimed to improve enzymes in the Calvin-Benson cycle to enhance the rate of ribulose 1,5-bisphosphate (RuBP) regeneration which limits carbon fixation under certain conditions. Computational models have suggested that the natural distribution of enzymes within the Calvin-Benson cycle is suboptimal, especially under elevated CO
2 concentrations [
14]. Overexpression of sedoheptulose 1,7-bisphosphatase or fructose-1,6-bisphosphate aldolase in
N. tabacum resulted in a higher carbon fixation rate and increased biomass at elevated CO
2 levels [
15,
16]. Even at ambient CO
2 concentration, overexpression of limiting enzymes in the Calvin cycle was shown to boost carbon fixation in tobacco [
17,
18,
19].
The next obvious approach centered around engineering photorespiration bypass pathways to metabolize the byproduct of RuBisCO oxygenation, phosphoglycolate, preventing it from entering the natural photorespiratory pathway thus minimizing the loss of CO
2 and energy. The first such bypass pathway involved engineering an
E. coli glycolate catabolic pathway consisting of three enzymes: glycolate dehydrogenase, glyoxylate carboligase, and tartronic semialdehyde reductase into Arabidopsis [
20]. The design aimed to mimic the natural photorespiration process of returning 75% glycolate to the Calvin-Benson cycle without ammonia production. Increases in photosynthesis rate and biomass were observed in these transgenic Arabidopsis. A second bypass aimed to convert glycolate into hydroxypyruvate in peroxisomes by two enzymes from
E. coli: glyoxylate carboligase and hydroxypyruvate isomerase [
21]. This bypass releases CO
2 in the peroxisome and does not produce extra ammonia. However, transgenic tobacco showed stunted growth in ambient air [
21].
Another bypass was designed to completely convert glycolate into CO
2 in the chloroplast [
22]. The pathway consisted of three new enzymes: Arabidopsis glycolate oxidase (directing the transgenic enzyme into the chloroplast instead of the peroxisome as for the endogenous one), pumpkin malate synthase, and
E. coli catalase. Together with two endogenous enzymes (NADP-malic enzyme and pyruvate dehydrogenase), this pathway mimics the
E. coli glycolate oxidative cycle in Arabidopsis chloroplasts. Increased photosynthesis rate and dry weight biomass were observed [
22]. A polyprotein comprising all three subunits of
E. coli glycolate dehydrogenase was expressed in plastids of transgenic potato, resulting in significantly higher carbohydrate production and a substantial increase in shoot and leaf biomass, as well as tuber yield [
23]. Recently, South et al. reported yet another bypass design that not only has a shunt pathway to metabolize glycolate utilizing the glycolate dehydrogenase from
Chlamydomonas reinhardtii and malate synthase from
Cucurbita maxima, but also includes an RNAi design to inhibit the plastidial glycolate transporter, plastidial glycolate glycerate translocator 1 (PLGG1), in an attempt to limit the introduction of glycolate into the photorespiration pathway [
24]. With a successful reduction in the expression of tobacco PLGG1, transgenic plants showed biomass increases as high as 40% in field conditions [
24].
The effectiveness of photorespiration bypass pathway designs to enhance photosynthesis efficiency has recently been tested in one crop species, rice. The first design, called the GOC bypass, consists of three rice enzymes: glycolate oxidase, oxalate oxidase, and catalase [
25] and, similar to Maier et al., aimed to completely metabolize glycolate into CO
2 in the chloroplast [
22]. Increases in photosynthetic rate and biomass were observed in transgenic rice, especially under high light conditions, but seed setting was reduced [
25]. A GCGT bypass design with four genes: a rice glycolate oxidase and three
E. coli enzymes, including catalase, glyoxylate carboligase, and tartronic semialdehyde reductase, was expected to lead to a greater increase in photosynthetic rate and grain yield by mimicking the natural 75% return of glycolate to the Calvin cycle as opposed to 100% conversion into CO
2 [
26]. Indeed, increases in photosynthesis rate, biomass, and grain yield were observed. However, like the GOC bypass, seed setting was reduced in transgenic rice. Interestingly, despite compromise on grain chalkiness, both GOC and GCGT grain showed higher nutritional value, including protein content and better cooking quality [
27].
In trees, however, there have rarely been any attempts to increase woody biomass via manipulations of enzymes or metabolic pathways for photosynthesis enhancement. In an attempt to create the next generation of trees using synthetic biology approaches to help combat the looming consequences of climate change, we selected a bypass design based on the work of South et al. to test the impact on photosynthetic efficiency in the poplar [
24]. This design includes an RNAi to reduce PLGG1 expression to limit glycolate transport out of the chloroplast, a transgenic chloroplast-targeted glycolate dehydrogenase to convert the retained glycolate into glyoxylate, and a transgenic chloroplast-targeted malate synthase to convert glyoxylate to malate [
24,
28]. Malate is then sequentially converted to pyruvate and acetyl-CoA. These reactions result in the release of CO
2 in the chloroplast, rather than the peroxisome as in the endogenous photorespiration pathway, allowing the released CO
2 to be fixed by RuBisCO carboxylation. While the growth performance of poplar plants harboring this design is under a multi-year field trial, this paper summarized the construct impact on transgenic plants grown in a controlled environment. If proven effective in the field and implemented on a large scale, the engineered trees could have the potential for substantial drawdown of CO
2, helping to counter the effects of climate change.
2. Materials and Methods
2.1. Poplar Plant Growth and Propagation
Hybrid Populus tremula × Populus alba, clone INRA 717-1B4, a widely-used laboratory clone, was a gift received from Dr. Steve Strauss of Oregon State University. Plants were kept in a growth room using horticultural LED lighting at a constant photosynthetic photon flux density (PPFD) of 120 μmol m−2s−1, with a 25 °C/22 °C day/night temperature, and humidity levels of 60% relative humidity. Parental plants were shaped to allow each plant to produce a minimum of 10 axillary shoots approximately 10–13 cm in length from which ramets would be collected. A number of cuttings were taken from each parental plant to create sister plants or ramets. Ramets were given a 0.35% IBA (indole-3-butyric acid) treatment then placed in rockwool plugs in trays containing water. Trays were kept in a controlled environment at a constant 24 °C temperature, 60% relative humidity, and a PPFD of 100 μmol m−2s−1 for root initiation. Ramets were given 14 days for root formation.
Once the ramets had sufficient root growth they were transplanted into 2-gallon pots and randomly distributed within the growth room. Similar growth conditions to the parental plants were applied to the ramets through the duration of the experiment. The growth room was maintained at a constant photosynthetic photon flux density (PPFD) of 120 or 280 μmol m−2s−1 measured at a 90 cm height, with a 25 °C/22 °C day/night temperature, and 60% relative humidity. Ramets were supplemented with 100 ppm nitrogen solution throughout the experiment. Plants were transplanted again after 10 weeks in the 2-gallon pots into 3-gallon pots and moved to a high ceiling growth room for biomass accumulation experiments.
2.2. Growth and Biomass Analysis
From one week post potting, phenotype evaluation was carried out on a weekly basis. This evaluation included height measurement, root collar diameter (RCD) measurement, leaf number counting, SPAD reading (soil plant analysis development: an instrument to measure chlorophyll content, used to determine the relative health of the plant), and morphology recording. Plant height and RCD were measured weekly. From these measurements we calculated plant volume as a proxy to above-ground biomass, using the formula:
For biomass analysis, the plants were harvested at 15 or 21 weeks. Plant material was separated into stems, leaves, and roots. Roots were removed from pots and washed with a gentle massage to remove residual soil, after which they were tapped dry on paper towels. The fresh weight of each material was recorded, then the material was dried in an oven at 60 °C for 7 days, after which dry weight was recorded.
2.3. Agrobacterium-Mediated Transformation of Poplar Hybrid Clone 717-1B4
Agrobacterium tumefaciens strain MP90 transformed with the binary vector pLC0102 was cultured in LB media with 200 µM acetosyringone for 24 h at 28 °C and shaken at 220 rpm. Stem explants isolated from in vitro propagated plants of hybrid Populus tremula × Populus alba, clone INRA 717-1B4, were used for Agrobacterium-mediated transformation. The explants were swirled in the Agrobacterium suspensions for 20 min, and then co-cultured on callus induction medium (CIM) [Murashige-Skoog (MS) medium supplemented with 10 µM NAA, 5 µM 2iP, 0.7% micropropagation Type I agar (Caisson Labs, Smithfield, UT, USA)] and incubated at 27 °C for 48 h in the dark. Explants were washed four times with deionized water with 250 mg/L cefotaxime, transferred to CIM with 100 mg/L Kanamycin, 250 mg/L cefotaxime and incubated for 21 days at 27 °C in the dark. Stem explants were then incubated under light (16 h light/8 h dark). Developing green calli were transferred to a shoot induction medium (SIM) [MS medium supplemented with 0.2 µM thidiazuron (TDZ), 100 mg/L Kanamycin, 250 mg/L cefotaxime, and 0.7% agar] for selection of transformed shoots and to control Agrobacterium overgrowth. Regenerated shoots were excised when they were approximately 1 cm long and transferred to rooting medium supplemented with ½ MS salts with vitamins, 0.5 µM indole-3-butyric acid (IBA), 50 mg/L kanamycin, and 250 mg/L cefotaxime. After 5–8 weeks, rooted and elongated shoots were propagated on the same medium in Magenta boxes. Rooted LC0102 transgenic plants were transferred into soil pots, acclimated, and grown under an inverted diurnal cycle. Eight weeks post potting, T0 plants went through a vegetative propagation cycle using a stem-cutting method frequently used in horticulture practice. Briefly, the apical meristem was removed to release the apical dominance to stimulate the formation of axillary buds for ramet propagation. To track the genetic lineage of transgenic lines, cuttings from the T0 plant are called C1 plants. Cuttings from C1 plants are called C2 plants.
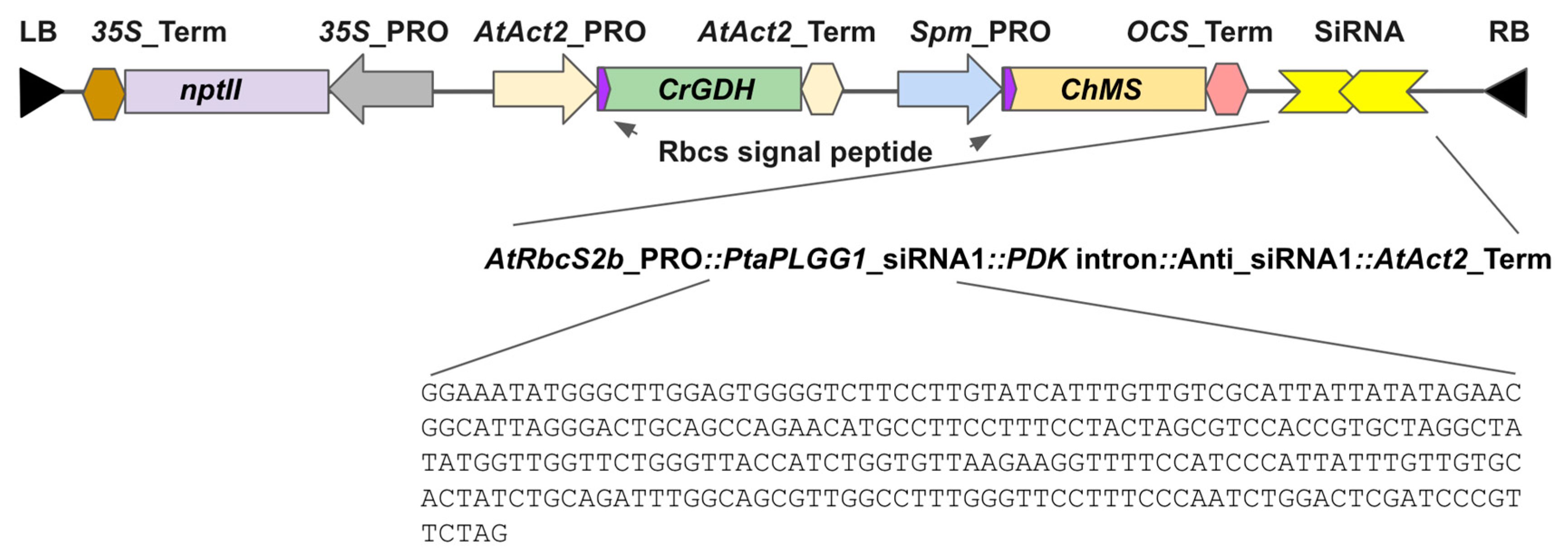
2.4. RNAi Design, Vector Construction, and Sequence Information
Poplar PLGG1 and various reference genes were identified through search of various databases including JGI’s Phytozome (
https://phytozome-next.jgi.doe.gov/, accessed on 7 December 2019), NCBI’s Populus nr database (
https://www.ncbi.nlm.nih.gov/, accessed on 6 July 2020), Popgenie (
https://popgenie.org, accessed on 6 July 2020), and AspenDB (
http://aspendb.uga.edu/, accessed on 1 May 2021). Using the amino acid sequence of plastidal glycolate/glycerate translocator in Arabidopsis (AtPLGG1) as the query led to the identification of PLGG1 homologs in
P. trichocarpa (Potri.003G099600.1) and
P. alba ×
P. tremula (XM_035032358 and Potra000595g04516.1, details described in results). To minimize the likelihood of off-target down-regulation, a region of PotriPLGG1 with minimal paralogy was selected. The RNAi region is 256 bp in length, 886–1142 of the transcript and 750–1006 measured from the start codon. RNAi element was designed as an intronic hairpin RNA interference system (ihRNAi) as in the pKANNIBAL plasmid series, which utilizes the PDK intron between a sense-antisense sequence of the previously defined target region [
29].
Construct LC0102 was designed computationally to resemble the bypass design of South et al. for expression in the poplar using Geneious Prime software (Boston, MA, USA), synthesized at GenScript (Piscataway, NJ, USA), and cloned into a binary vector pCambia3200 that was then introduced into Agrobacterium MP90 for poplar transformation.
GenBank, AspenDB, and Popgenie accession number of gene sequences in this work: AtAct2, U41998.1; AtPLGG1, AT1G32080; AtRbcS, NM_105379.4; AtRbcS2b, X14564.1; ChMS, X56948.1; CrGDH, XP_001695381.1; FtPDK, X79095.1; OCS_Ter, CP033030.1; PaPLGG1, XM_035032358; PotriPLGG1, Potri.003G099600.1; PtaAct, Potri.001G309500, sPta717, v1.1; PtaEF1B-1, Potri.001G224700, sPta717, v1.1; PtaPP2A-2, Potri.015G068300, sPta717, v1.1; PtaRP, Potri.001G342500, sPta717, v1.1; PtrPLGG1, Potra000595g04516.1; Spm_PRO, M25427.1.
2.5. Nucleic Acid Extraction and Quantification
Approximately 100 mg of leaf tissue from the second to fourth fully emerged leaf from the apical bud was used for DNA extraction using the NucleoSpin Plant II kit (Macherey-Nagel, Allentown, PA, USA). Leaf tissue was disrupted with 400 µL PL1 from the kit using BeadBeater (BioSpec, Bartlesville, OK, USA) at 2400 rpm, 1 min with four 2.3 mm steel beads 6 times until the tissue was fully homogenized. 10 µL of RNase was added to the homogenized tissues then incubated at 65 °C for 10 min. Samples then proceeded with filtration, binding, washing, and elution steps according to the manufacturer’s protocol.
For RNA extraction, 30 mg of leaf tissue was collected from plants that were at the end of 8 weeks after being transplanted to the 2-gallon pot. Tissues were soaked with 110 µL lysis buffer containing 100 µL PFL and 10 µL PFR from the NucleoSpin RNA Plant and Fungi kit (Macherey-Nagel, Allentown, PA, USA) and disrupted using BeadBeater (BioSpec, Bartlesville, OK, USA) at 2400 rpm, 30 s with four 2.3 mm steel beads 2–3 times until fully homogenized. An amount of 440 µL of lysis buffer containing 400 µL PFL and 40 µL PFR were added into the homogenized sample and incubated at 56 °C for 5 min. Samples were then treated with 95 µL DNase for 15 min at RT after PFW1.
DNA and RNA concentrations were measured using Cytation 1 plate reader with Take3Trio plates (Agilent, Santa Clara, CA, USA) in 2 µL volume.
2.6. Primer Design and Evaluation
Primers and probes were designed using PrimerQuest Tool (Integrated DNA Technologies, Coralville, IA, USA) with default settings. Gene sequences were obtained from NCBI or AspenDB. Each primer set was mapped to the
P. tremula and
P. alba sequences obtained from AspenDB using Geneious Prime and manually adjusted to ensure primer specificity and universal amplification from both alleles. The specificity of the primers was further validated using PCR and DNA gel electrophoresis on 1% (
w/
v) agarose gels containing 1× SYBR Green I Nucleic Acid Gel Stain (Thermo Fisher Scientific, Waltham, MA, USA) in 1× TAE buffer. Primer efficiencies were evaluated with qPCR or qRT-PCR in singleplex condition using DNA or RNA purified from transgenic plants as standard with 5× dilution series starting from 100 ng and 1 µg respectively. Standard curves were generated using CFX Maestro software (Bio-Rad, Hercules, CA, USA) to calculate the primer efficiency. Primer sequence, concentration, and amplicon length are listed in
Table S1.
2.7. PCR, qPCR, and qRT-PCR
PCR analysis was performed to provide quick validation of transgenic status or primer specificity (
Table S1). The 20 µL reaction contains 20 ng genomic DNA, 1× GoTaq Master Mix (Promega, Madison, WI, USA), and 500 nM each of forward and reverse primers. PCR program: 2 min 95 °C initial denaturation; 35 cycles (95 °C, 30 s; 54.1 °C, 30 s; 72 °C, 2 min); 5 min 72 °C final extension.
Multiplex qPCR analysis with technical quadruplicates in each event was used for copy number estimation using a CFX Opus 96 machine (Bio-Rad). Multiplex condition was validated by comparing singleplex and multiplex results to see if the Cq value changes according to the standard mentioned in the primer design section. The 20 µL reaction contains 20 ng genomic DNA, 1× TaqPath ProAmp Multiplex Master Mix (Thermo Fisher Scientific, Waltham, MA, USA), primers, and probes (
Table S1). qPCR was performed with the following conditions: 95 °C, 10 min; 40 cycles (95 °C, 15 s; 60 °C, 30 s). RP was selected as the reference gene with a copy number of 2. The estimated copy number of the target was calculated using CopyCaller (Thermo Fisher Scientific, Waltham, USA). The copy number was calculated based on 2
(−ΔΔCq)×copy number of the target gene in the calibrator, where ΔCq = Cq
target gene − Cq
reference gene, and ΔΔCq = ΔCq
sample − ΔCq
calibrator.
Multiplex qRT-PCR analysis with technical duplicates for each event using CFX Opus 96 machine (Bio-Rad, Hercules, CA, USA) was used for gene expression analysis, with multiplex conditions validated as described previously. The 20 µL reaction contains 200 ng total RNA, 1× custom-made One-Step RT-qPCR Master Mix with a lower amount of DTT (Launchworks, Bothell, WA, USA), primers, and probes (
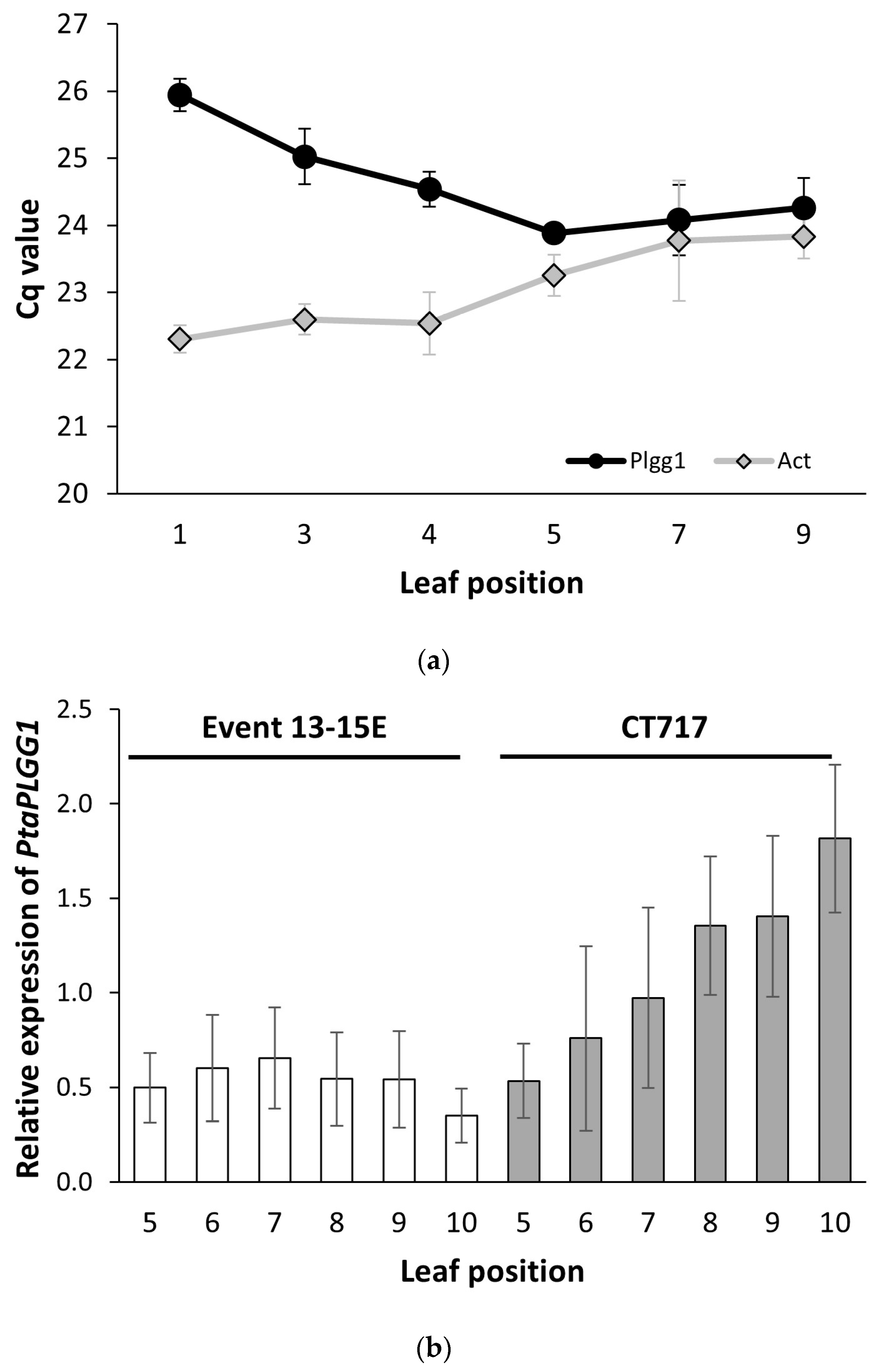
Table S1). qRT-PCR was performed with the following conditions: 53 °C, 10 min, reverse transcription; 95 °C, 2 min, initial denaturation; 40 cycles amplification (15 s 95 °C denaturation; 30 s 60 °C annealing/ extension). Reference genes were selected based on expression evaluation in leaves of various developmental stages.
PtaAct,
PtaEF1B-1,
PtaRP, and
PtaPP2A-2 were selected for testing based on previous reports [
30,
31,
32,
33,
34,
35]. The average Cq of the selected reference genes were used for ΔCq calculation. The 2
(−ΔΔCq) method was used to analyze the expression data.
2.8. Gas-Exchange Measurements
Photosynthesis activity was measured using a Li-Cor 6800 infrared gas analyzer (Li-Cor Biosciences, Lincoln, NE, USA) on poplar plants grown in a controlled environment as described in the plant growth and propagation section. Measurements were performed on various developmental stages of poplar plants depending on the experimental plan. Generally Leaf 8 or 9, counting from the first leaf with a petiole separated from the apical bud, was selected for these measurements and all measurements were taken between 8 am and 2 pm. The leaves were maintained at 25 °C, 1500 PAR, with VPD fixed at 1.3 kPa. For photosynthetic CO
2 response curves or
A-Ci curves, before starting the measurements leaves were acclimated at 410 ppm CO
2 for a minimum of 8 min until a steady state was reached. Once steady state had been achieved, two data points were logged with a 1.5 min interval to record ambient
Anet (net CO
2 assimilation under ambient 410 ppm CO
2). Following these ambient measurements, the leaf was subjected to a series of CO
2 concentrations ranging from 0–2000 ppm CO
2 (400, 300, 200, 100, 50, 0, 400, 600, 800, 1000, 1200, 1500, 2000), and measurements were taken when assimilation reached a steady-state rate at each CO
2 setting.
Jmax and
Vccmax were derived from an
A-Ci curve fitted using the R package “plantecophys” [
36].
Jmax is the maximum rate of photosynthetic electron transport,
Vcmax is the maximum rate of carboxylation), and
Amax is the CO
2 assimilation rate at 2000 ppm CO
2.
2.9. Statistical Analyses
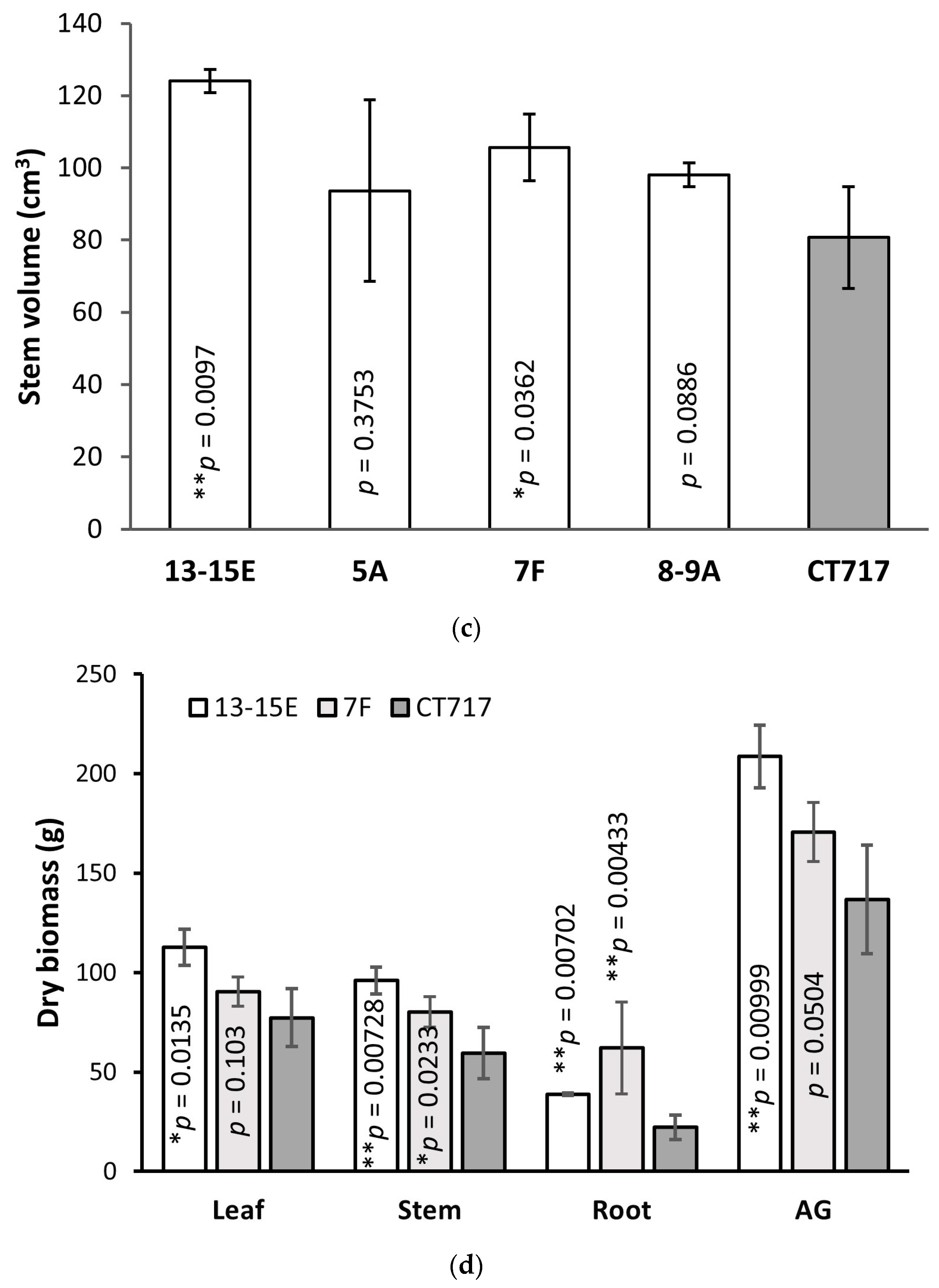
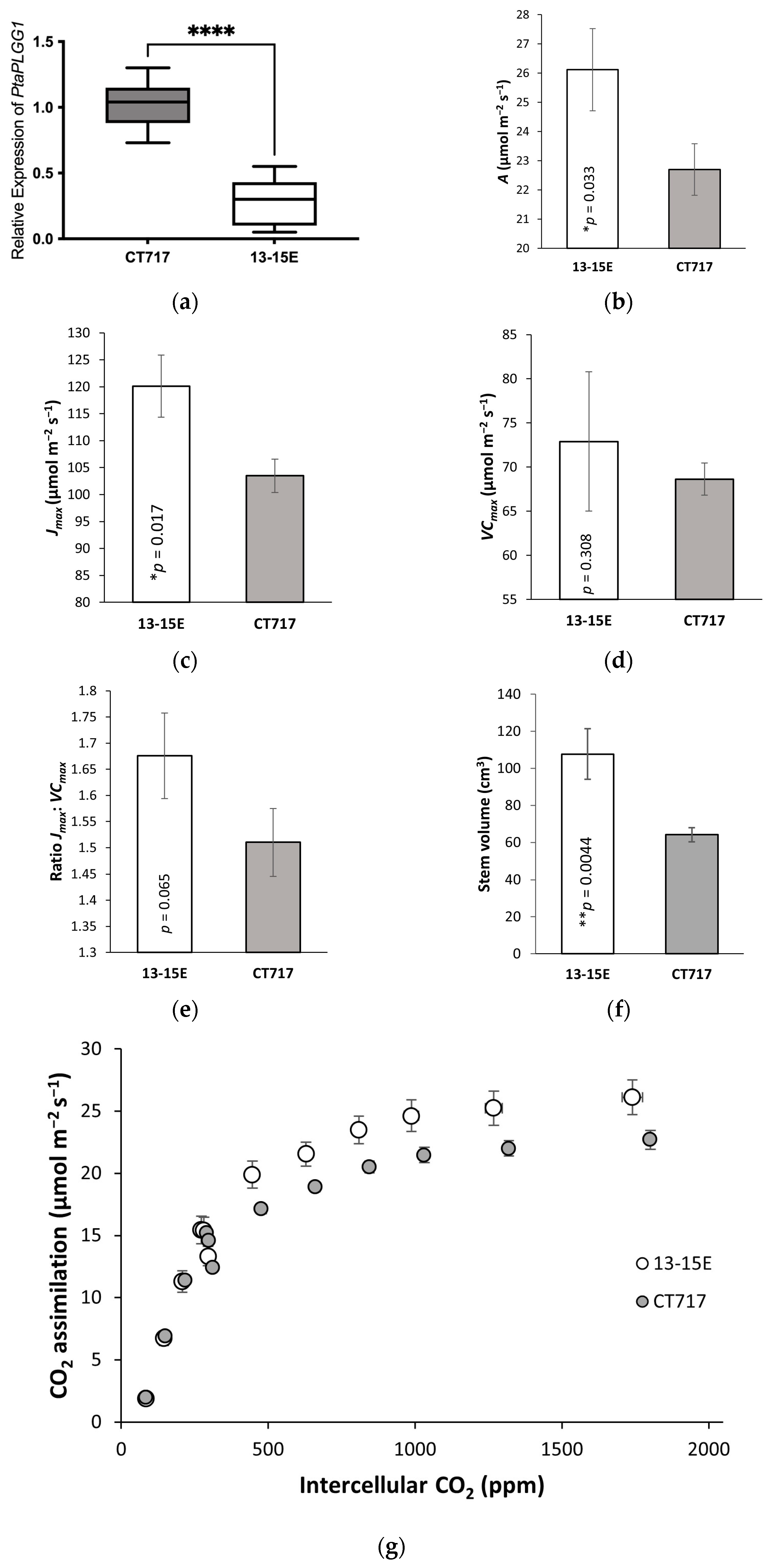
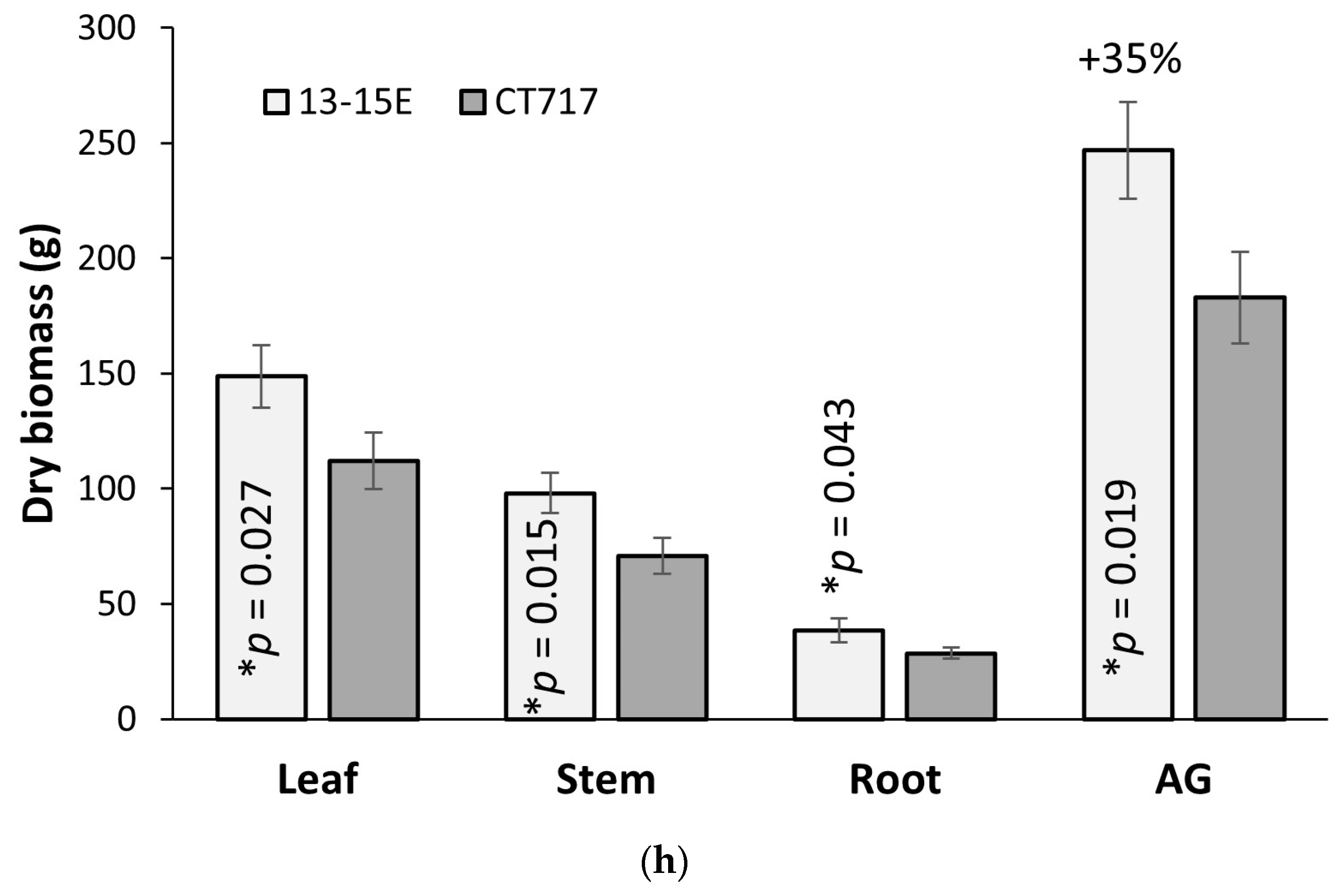
Statistical analysis of gene expression data was performed using Prism (GraphPad, Boston, MA, USA). Statistical significance was reported based on ANOVA followed by Tukey’s multiple comparison tests. *, **, and **** represent a significant difference at p < 0.05, p < 0.01 and p < 0.0001 respectively. Statistical significance of gas exchange, growth rate, and biomass data was reported based on one-tailed t-tests, comparing each transgenic event to the non-transgenic controls. Significance is represented by * (p < 0.05) and ** (p < 0.01).
4. Discussion
A number of photorespiration bypass pathways have been conceived and tested in C3 annuals thanks to the advancement of knowledge in synthetic biology [
20,
21,
22,
23,
24,
25,
26]. Significant increases in photosynthetic efficiency and crop grain yield have been observed [
25,
26]. After evaluation of design principles, we elected to test a pathway in conjunction with glycolate transporter inhibition in the poplar hybrid INRA 717-1B4 [
24]. The results of molecular analysis and photobiology measurements support our observation that this bypass increased biomass production in the poplars by 35%–53% compared to the control plants that do not have the functional bypass pathway (
Figure 4 and
Figure 5). To our knowledge, this is the first time that photosynthetic efficiency has been enhanced in a tree species by such a magnitude.
We reasoned that for this photorespiration bypass to work in poplars, a reduction in the expression level of the glycolate transporter is critical. As stated before, INRA 717-1B4 is a hybrid clone of
P. alba ×
P. tremula. Comparison of the coding sequences of
PLGG1 between
P. alba and
P. tremula revealed 10 allelic differences with a 6 bp gap and 9 SNPs. Our RNAi design avoided sequences containing the 6 bp gap. The various levels of
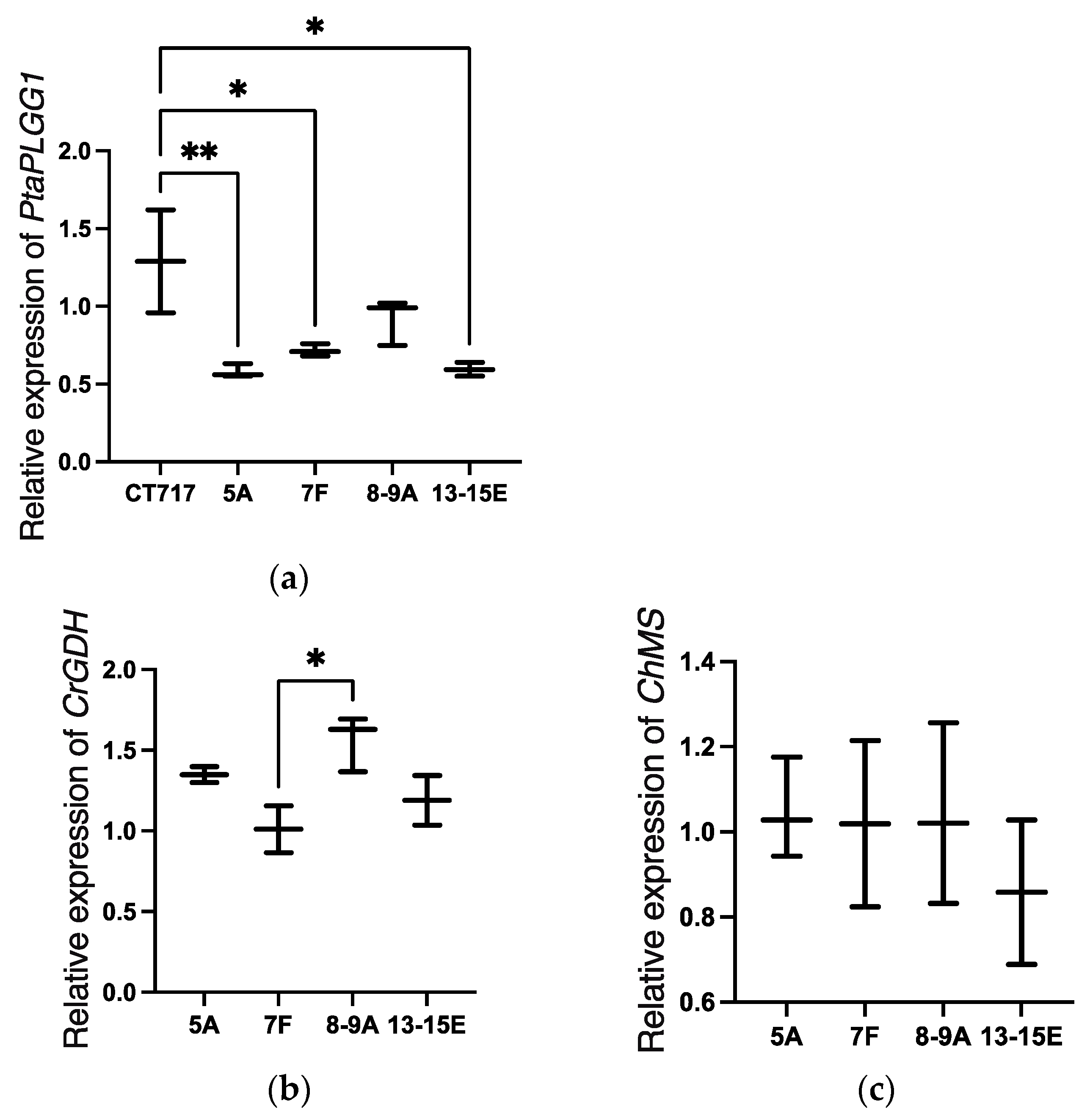
PtaPLGG1 reduction in our transgenic events (
Figure 3a) validated the RNAi design principle. The wide range in reduction of the
PtaPLGG1 expression in transgenic events would also allow for the selection of events with the appropriate amount of glycolate transportation inhibition. However, plant gene expression is well known to vary with growth and developmental stages as well as physiological changes due to (a)biotic environmental changes. Our qRT-PCR analysis was established and optimized through consideration of various factors including the choice of reference genes, the age of plants when samples were collected, and growth conditions, especially light intensity, etc. This analysis protocol enabled minimization of plant-to-plant variation.
In addition to the range of
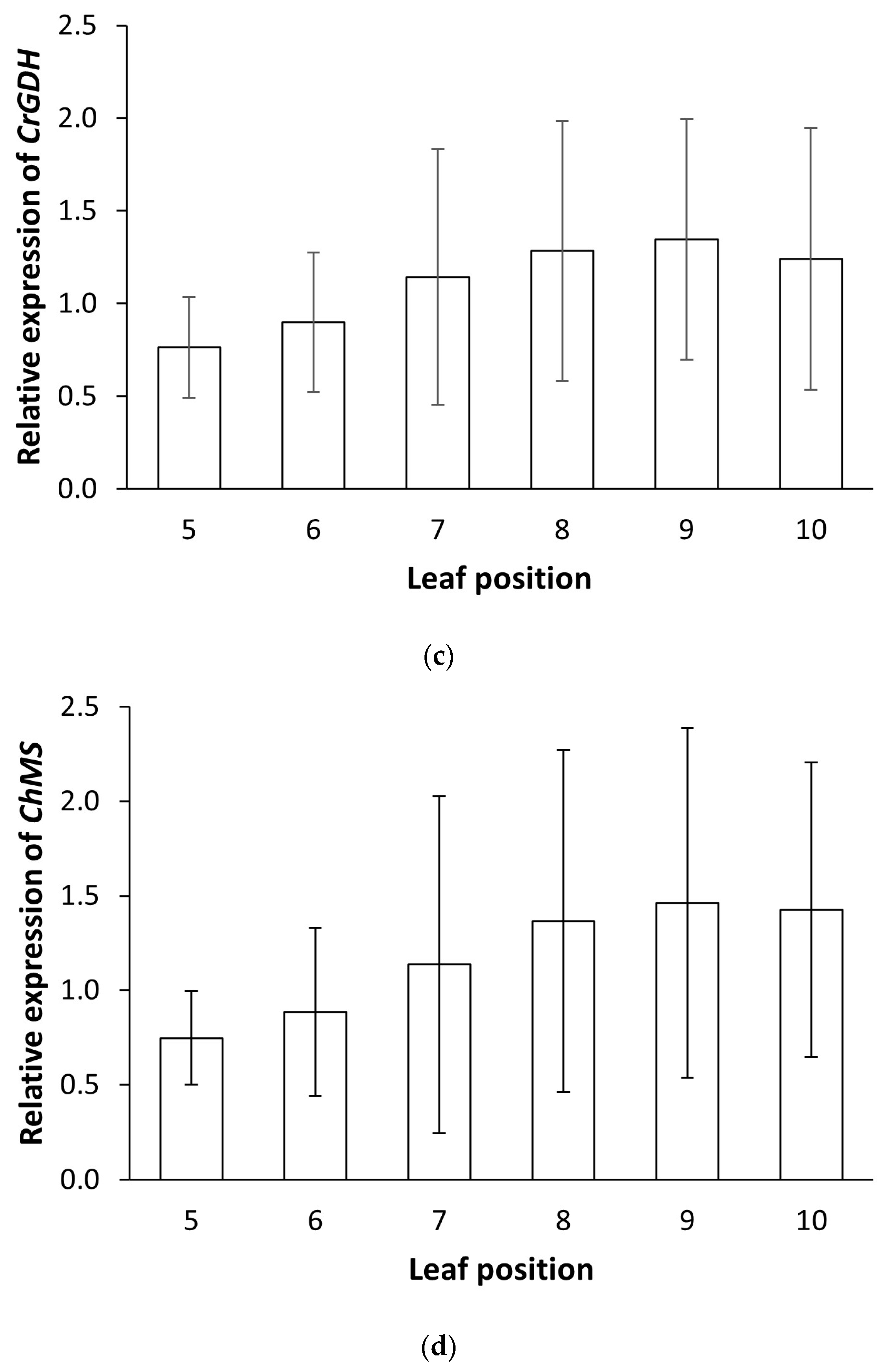
PtaPLGG1 expression reduction, it is desirable to have a range of expression levels for the two transgenic metabolic enzymes. This allows selection of a balanced perturbation of the metabolic flow that would enable an optimized output from the shunt pathway for increased photosynthetic activity. We observed a range of
CrGDH and
ChMS expression levels in the 41 T0 events (
Table S3). Fine-tuned metabolic perturbation was also achieved at the level of post-transcriptional control, such as feedback inhibition and modulation. It would be interesting to quantitatively analyze the levels of related proteins in transgenic plants. Furthermore, enzymatic activities, governed by enzyme kinetic characteristics, provide another degree of fine-tuning regulation on metabolic output. Enzyme engineering through computational design using software such as Rosetta to discern the relationship of protein structure and function has been widely applied in the field of synthetic biology [
37,
38]. With the ever-increasing data on structure-function relationship and the knowledge derived from machine learning on enzyme activities, the future of protein design for biotech applications can only improve. Enzyme engineering through directed evolution (DE) to accumulate beneficial amino acid sequence changes, thereby improving enzymatic activity, has proven to be practical and fruitful in synthetic biology applications [
6,
39,
40,
41]. Examples of using DE to improve photosynthetic efficiency have been reported, albeit not as successful [
6]. Challenges are not with the technology itself, but rather with how to properly design and screen the enzyme variants that would exert the activity change in a living plant. Therefore, enzyme engineering provides the potential for improving the efficiency of the shunt pathway in the future. Additionally, changes in the perturbation of endogenous metabolic flow can be monitored via metabolite profiling experiments or metabolomics analysis. It remains unclear what changes in the metabolite profile of Events 13-15E and7F contribute to the increased biomass production. We will incorporate metabolite analysis in future trials of these transgenic events to further investigate the mechanism behind the increase in biomass.
When vegetatively propagated plants experience changes in environmental and growth conditions, ranging from mechanical damage, to lack of nutrients during rooting, to transplanting shock, they need a period of time post potting for acclimation and to stabilize, a term used in the field to describe when genetics start to have the dominant effect on the growth characteristics of plants. The growth performance of C1 plants showed a wide range of variation during the first few weeks of growth after transplanting in the growth room. Our results, based on
Vcmax measurement of photosynthesis activity in poplar plants after transplanting, helped to evaluate stabilization at the physiological level (
Figure S1). Additional research is needed to better understand the process of stabilization and to narrow down the time frame that the poplar needs to fully acclimate to soil growth.
The impact of engineered photorespiration bypass on the activity of photosynthesis can be measured with conventional gas-exchange methodologies. Changes in
Jmax and the ratio of
Jmax:
Vcmax reflect changes in photosynthetic activity. For a given plant species,
Vcmax fluctuates with the physiological state of the plant and may also change under the influences of growth and environmental conditions, such as (a)biotic stresses, including transplanting acclimation (
Figure S1). However, for plants of the same species grown under the same growth conditions,
Vcmax fluctuations should not significantly differ from each other. We reason that we should not expect changes in the values of
Vcmax between transgenic plants and non-transgenic controls because the photorespiration bypass design has not altered RuBisCO carboxylation activity.
The increase in biomass production in these engineered poplar trees may not fully reflect the efficiency enhancement in photosynthesis. As the amount of photosynthetic carbohydrates increases in the source tissue, transportation and deposition of these energy-rich carbon molecules to the sink tissue become limiting factors. As such, it would be valuable to assess carbon partitioning and source-sink dynamics in these transgenic trees in the future. Alternative photorespiration bypass pathways have been tested in other C3 plant species. In some cases, the pathway did not work as intended [
42]. In other cases, the pathway seemed to work properly and resulted in enhanced photosynthetic rate, increased biomass and grain yield, however, the seed setting was reduced [
25,
26]. Through an integrated analysis of transcriptomics, physiology, and biochemistry, Wang et al. concluded that photosynthetic carbohydrates in these plants were not transported to grains in an efficient manner [
26]. In woody plants such as poplar, this source-sink carbon partitioning issue may become limiting when photosynthesis is enhanced. Other issues such as C-N balance may add to the complexity as well. Nonetheless, woody plants may offer a simpler system than cereal crops due to complexities in regulating the transition from vegetative growth to reproductive growth for grain production.
While the current study was conducted in controlled growth room settings and produced encouraging results, it is also important to test the photosynthetically enhanced transgenic events in a field setting where trees will be subjected to more (a)biotic stress and higher light intensity. As such, a multi-year field trial of many of our transgenic events is currently underway. It will be interesting to see how the field trial results align with that from growth room studies.



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







