
Inconsistent Growth Responses of Alpine Rhododendron Shrubs to Climate Change at Two Sites on the Eastern Tibetan Plateau




Abstract
:1. Introduction
2. Materials and Methods
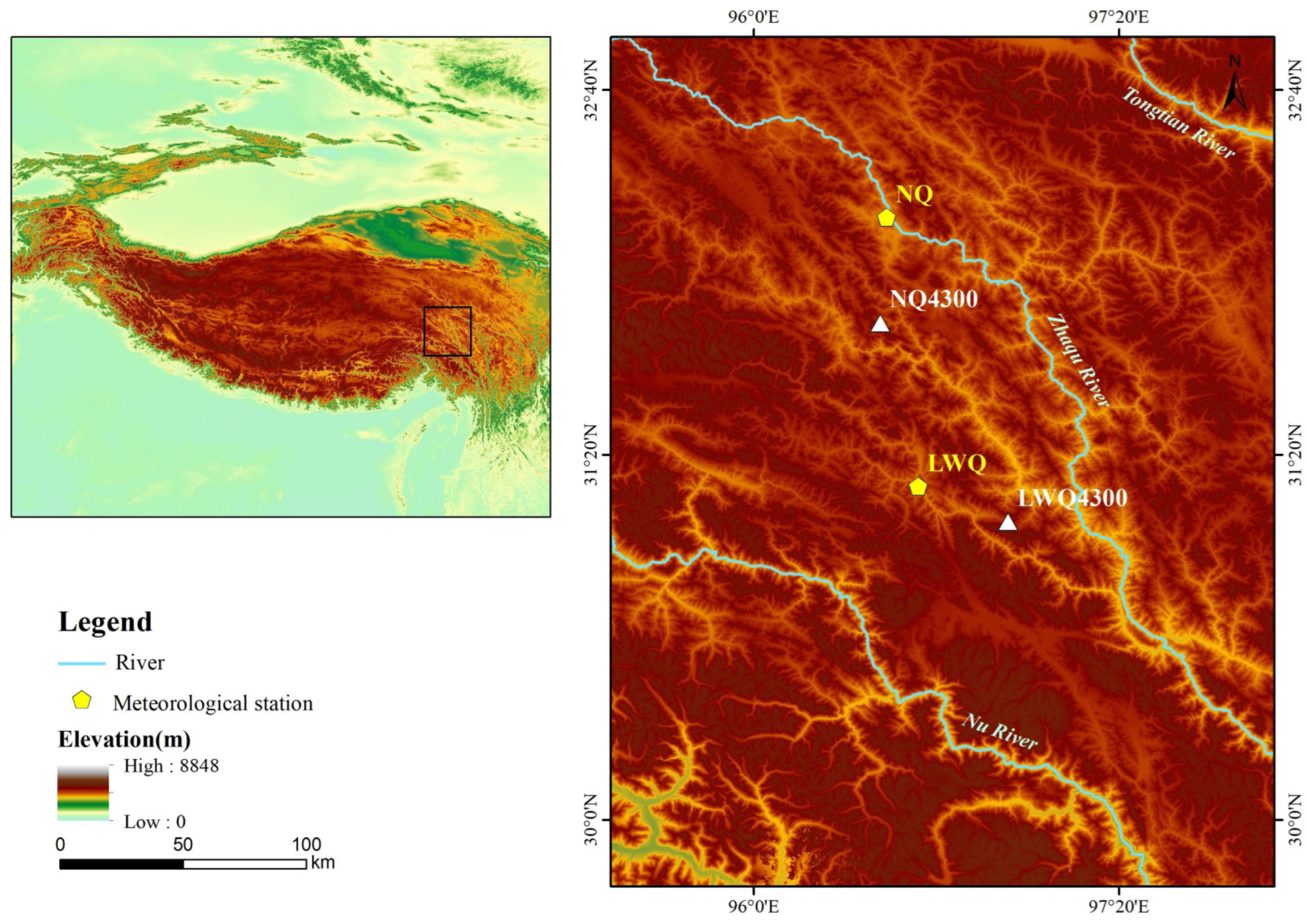
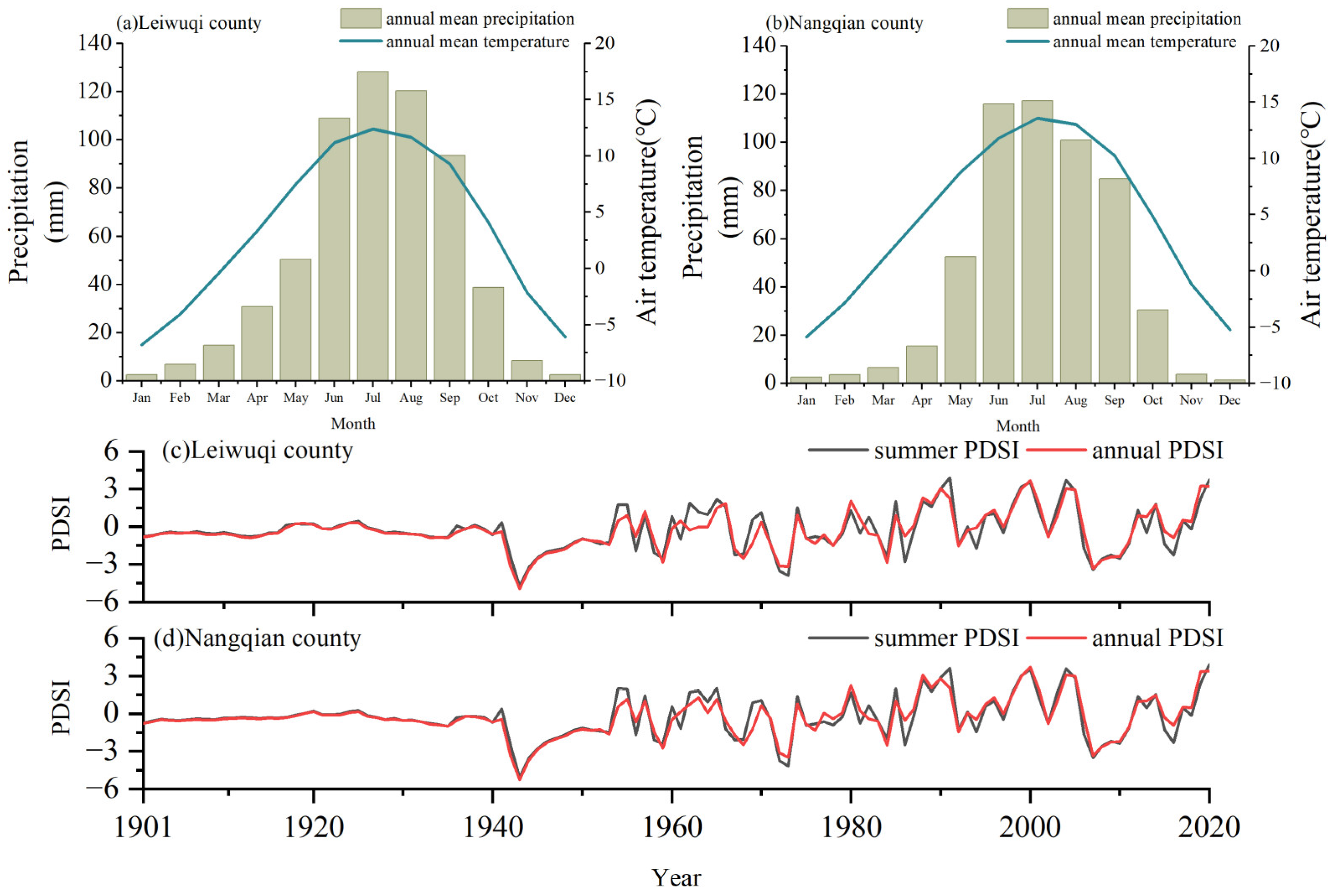
2.1. Study Area and Study Species
2.2. Sample Collection and Chronology Development
2.3. Analysis of Climate–Growth Relationships
3. Results
3.1. Characteristics of Shrub Chronologies
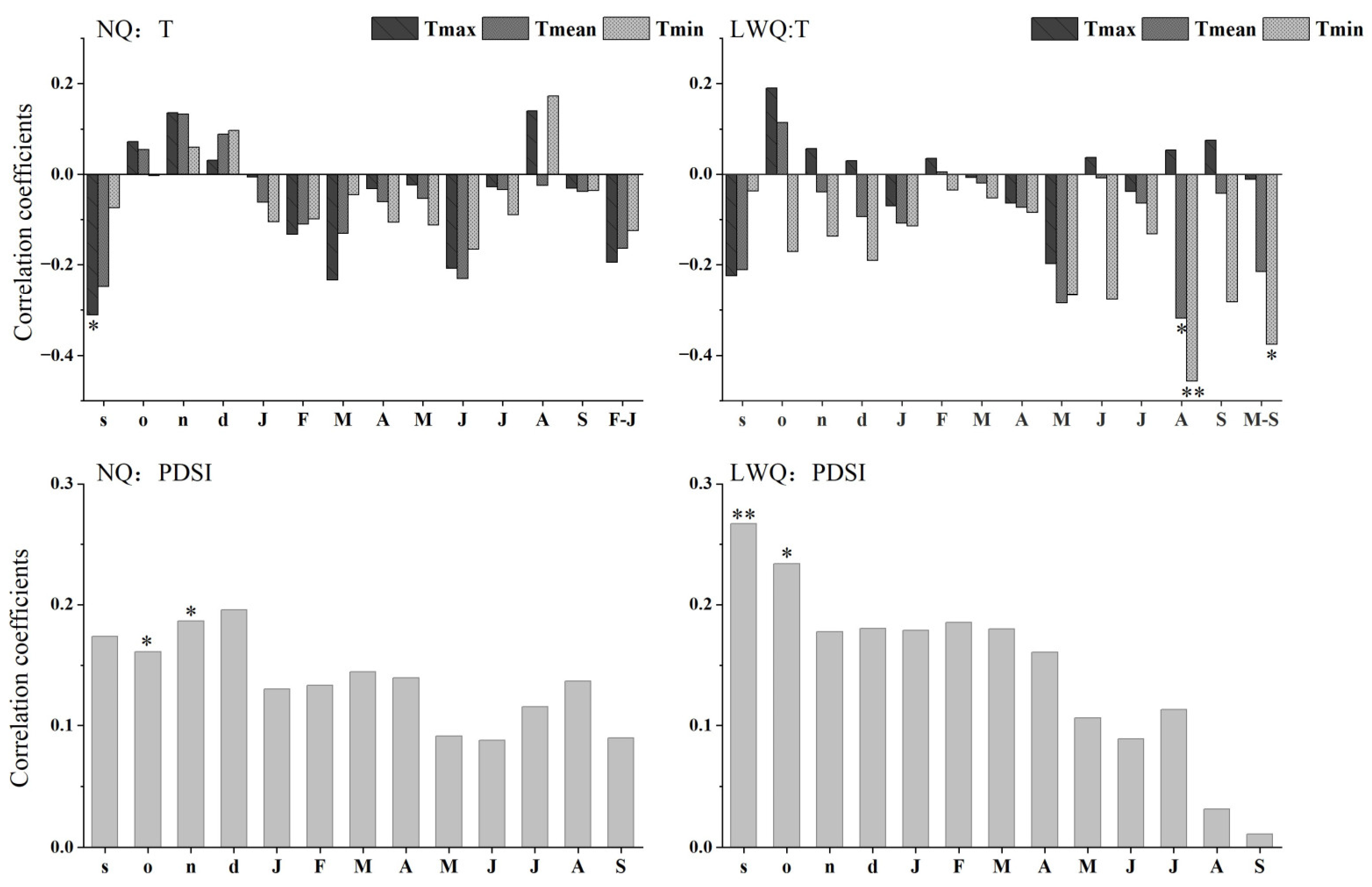
3.2. Climate–Growth Relationships
3.3. Partial Correlation Analysis
3.4. Predictors of Shrub Growth
4. Discussion
4.1. Growth-Limiting Factor for Rhododendron przewalskii Shrubs
4.2. Comparison between the Results of Previous Studies and this Study
4.3. Potential Influence of Climate Warming on Alpine Shrubs
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peng, D.; Wang, Y.; Xian, G.; Huete, A.R.; Huang, W.; Shen, M.; Wang, F.; Yu, L.; Liu, L.; Xie, Q.; et al. Investigation of land surface phenology detections in shrublands using multiple scale satellite data. Remote Sens. Environ. 2021, 252, 112133. [Google Scholar] [CrossRef]
- National Forestry and Grassland Administration. The Ninth National Forest Resources Inventory Report. Available online: http://www.forestry.gov.cn/gjslzyqc.html (accessed on 2 October 2022).
- Cáceres, Y.; Llambí, L.D.; Rada, F. Shrubs as foundation species in a high tropical alpine ecosystem: A multi-scale analysis of plant spatial interactions. Plant Ecol. Divers. 2015, 8, 147–161. [Google Scholar] [CrossRef]
- Wezel, A.; Rajot, J.-L.; Herbrig, C. Influence of shrubs on soil characteristics and their function in Sahelian agro-ecosystems in semi-arid Niger. J. Arid Environ. 2000, 44, 383–398. [Google Scholar] [CrossRef]
- Musa, A.; Deming, J.; Cunyang, N. The applicable density of sand-fixing shrub plantation in Horqin Sand Land of Northeastern China. Ecol. Eng. 2014, 64, 250–254. [Google Scholar] [CrossRef]
- Xiao, S.; Peng, X.; Tian, Q.; Zhu, G. Stem radial growth indicate the options of species, topography and stand management for artificial forests in the western Loess Plateau, China. Sci. Cold Arid Reg. 2019, 11, 226–238. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Hallinger, M.; Blok, D.; Sass-Klaassen, U.; Rayback, S.A.; Weijers, S.; Trant, A.J.; Tape, K.D.; Naito, A.T.; Wipf, S.; et al. Methods for measuring arctic and alpine shrub growth: A review. Earth-Sci. Rev. 2015, 140, 1–13. [Google Scholar] [CrossRef]
- Escaravage, N.; Questiau, S.; Pornon, A.; Doche, B.; Taberlet, P. Clonal diversity in a Rhododendron ferrugineum L. (Ericaceae) population inferred from AFLP markers. Mol. Ecol. 1998, 7, 975–982. [Google Scholar] [CrossRef]
- Schweingruber, F.H.; Poschlod, P. Growth Rings in Herbs and Shrubs: Life Span, Age Determination and Stem Anatomy; Swiss Federal Research Institute WSL Birmensdorf: Birmensdorf, Switzerland, 2005; Volume 79. [Google Scholar]
- Kumar, P. Assessment of impact of climate change on Rhododendrons in Sikkim Himalayas using Maxent modelling: Limitations and challenges. Biodivers. Conserv. 2012, 21, 1251–1266. [Google Scholar] [CrossRef]
- Francon, L.; Corona, C.; Roussel, E.; Saez, J.L.; Stoffel, M. Warm summers and moderate winter precipitation boost Rhododendron ferrugineum L. growth in the Taillefer massif (French Alps). Sci. Total Environ. 2017, 586, 1020–1031. [Google Scholar] [CrossRef]
- Lu, X.; Camarero, J.J.; Wang, Y.; Liang, E.; Eckstein, D. Up to 400-year-old Rhododendron shrubs on the southeastern Tibetan Plateau: Prospects for shrub-based dendrochronology. Boreas 2015, 44, 760–768. [Google Scholar] [CrossRef]
- Liang, E.; Eckstein, D. Dendrochronological potential of the alpine shrub Rhododendron nivale on the south-eastern Tibetan Plateau. Ann. Bot. 2009, 104, 665–670. [Google Scholar] [CrossRef]
- Panthi, S.; Fan, Z.-X.; Bräuning, A. Ring widths of Rhododendron shrubs reveal a persistent winter warming in the central Himalaya. Dendrochronologia 2021, 65, 125799. [Google Scholar] [CrossRef]
- Bi, Y.; Xu, J.; Yang, J.; Li, Z.; Gebrekirstos, A.; Liang, E.; Zhang, S.; Yang, Y.; Yang, Y.; Yang, X. Ring-widths of the above tree-line shrub Rhododendron reveal the change of minimum winter temperature over the past 211 years in Southwestern China. Clim. Dyn. 2017, 48, 3919–3933. [Google Scholar] [CrossRef]
- Francon, L.; Corona, C.; Till-Bottraud, I.; Carlson, B.Z.; Stoffel, M. Some (do not) like it hot: Shrub growth is hampered by heat and drought at the alpine treeline in recent decades. Am. J. Bot. 2020, 107, 607–617. [Google Scholar] [CrossRef]
- Young, A.B.; Watts, D.A.; Taylor, A.H.; Post, E. Species and site differences influence climate-shrub growth responses in West Greenland. Dendrochronologia 2016, 37, 69–78. [Google Scholar] [CrossRef]
- Hallinger, M.; Manthey, M.; Wilmking, M. Establishing a missing link: Warm summers and winter snow cover promote shrub expansion into alpine tundra in Scandinavia. New Phytol. 2010, 186, 890–899. [Google Scholar] [CrossRef]
- Su, K.; Chen, Y.; Yang, H. Advances in the rhododendron and rhododendron shrub communities. J. Trop. Subtrop. Bot. 2020, 28, 527–536. (In Chinese) [Google Scholar]
- Zhuang, P.; Wang, F.; Shao, H. Comparative study on Rhododendron and its distribution in W-Sichuan and SE-Tibet. Guihaia 2013, 33, 791–797. (In Chinese) [Google Scholar]
- Li, Z.; Liu, G.; Fu, B.; Zhang, Q.; Ma, K.; Pederson, N. The growth-ring variations of alpine shrub Rhododendron przewalskii reflect regional climate signals in the alpine environment of Miyaluo Town in Western Sichuan Province, China. Acta Ecol. Sin. 2013, 33, 23–31. [Google Scholar] [CrossRef]
- Liang, E.; Shao, X.; Qin, N. Tree-ring based summer temperature reconstruction for the source region of the Yangtze River on the Tibetan Plateau. Glob. Planet. Chang. 2008, 61, 313–320. [Google Scholar] [CrossRef]
- Wang, Y.; Li, D.; Ren, P.; Ram Sigdel, S.; Camarero, J.J. Heterogeneous Responses of alpine treelines to climate warming across the Tibetan Plateau. Forests 2022, 13, 788. [Google Scholar] [CrossRef]
- Alley, W.M. The Palmer drought severity index: Limitations and assumptions. J. Appl. Meteorol. Climatol. 1984, 23, 1100–1109. [Google Scholar] [CrossRef]
- Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 2020, 7, 109. [Google Scholar] [CrossRef] [PubMed]
- Wen, X. An overview of China climate change over the 20th century using UK UEA/CRU high resolution grid data. Chin. J. Atmos. Sci. 2006, 5, 894–904. (In Chinese) [Google Scholar]
- Geng, Y. The Genus Rhododendron of China; Shanghai Scientific & Technical Publishers: Shanghai, China, 2014. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 51–67. [Google Scholar]
- Cook, E.R.; Krusic, P.J.; Peters, K.; Holmes, R.L. Program ARSTAN (Version 4.4), Autoregressive Tree–Ring Standardization Program; Tree–Ring Laboratory of Lamont–Doherty Earth Observatory: Columbia, IN, USA, 2017. [Google Scholar]
- Meko, D.; Touchan, R.; Anchukaitis, K. Seascorr: A MATLAB program for identifying the seasonal climate signal in an annual tree-ring time series. Comput. Geosci. 2011, 37, 1234–1241. [Google Scholar] [CrossRef]
- Anderson, D.; Burnham, K. Model Selection and Multi-Model Inference; Springer: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Pinheiro, J.; Bates, D.; Deb Roy, S.; Sarkar, D.; Team, R.C. Linear and Nonlinear Mixed Effects Models. Available online: https://www.yumpu.com/ (accessed on 12 October 2022).
- Liang, E.; Lu, X.; Ren, P.; Li, X.; Zhu, L.; Eckstein, D. Annual increments of juniper dwarf shrubs above the tree line on the central Tibetan Plateau: A useful climatic proxy. Ann. Bot. 2011, 109, 721–728. [Google Scholar] [CrossRef]
- Li, X.; Rossi, S.; Liang, E.; Camarero, J.J. Temperature thresholds for the onset of xylogenesis in alpine shrubs on the Tibetan Plateau. Trees 2016, 30, 2091–2099. [Google Scholar] [CrossRef]
- Harlod, C. Tree Rings and Climate; Academic Press: London, UK, 1976. [Google Scholar]
- Guo, B.; Zhang, Y.; Wang, X. Response of Picea purpurea and Abies faxoniana tree rings at different slope aspects to rapid warming in western Sichuan. Chin. J. Appl. Ecol. 2016, 27, 354–364. (In Chinese) [Google Scholar]
- Li, H.; Ma, Y.; Bai, Y. An analysis of the impact of climate change on vegetation succession in qinghai province. J. Glaciol. Geocryol. 2010, 32, 414–421. (In Chinese) [Google Scholar]
- Kagawa, A.; Sugimoto, A.; Maximov, T.C. Seasonal course of translocation, storage and remobilization of 13 C pulse-labeled photoassimilate in naturally growing Larix gmelinii saplings. New Phytol. 2006, 171, 793–804. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Chapin, E.S.F.S.; Schulze, E.; Mooney, H.A. The Ecology and Economics of Storage in Plants. Annu. Rev. Ecol. Syst. 1990, 21, 423–447. [Google Scholar] [CrossRef]
- Han, Y.; Wang, Y.; Liu, B.; Huang, R.; Camarero, J.J. Moisture mediates temperature-growth couplings of high-elevation shrubs in the Tibetan plateau. Trees 2022, 36, 273–281. [Google Scholar] [CrossRef]
- Mu, H.-X.; Han, F.; Zhang, B.-P.; Liang, T.; Wang, Z.-Y.; Wang, Z. Characteristics of timberline and treeline altitudinal distribution in Mt. Namjagbarwa and their geographical interpretation. J. Mt. Sci. 2022, 19, 2846–2860. [Google Scholar] [CrossRef]
- Yang, R.-Q.; Zhao, F.; Fan, Z.-X.; Panthi, S.; Fu, P.-L.; Bräuning, A.; Grießinger, J.; Li, Z.-S. Long-term growth trends of Abies delavayi and its physiological responses to a warming climate in the Cangshan Mountains, southwestern China. For. Ecol. Manag. 2022, 505, 119943. [Google Scholar] [CrossRef]
- Porter, T.J.; Pisaric, M.F.J. Temperature-growth divergence in white spruce forests of Old Crow Flats, Yukon Territory, and adjacent regions of northwestern North America. Glob. Chang. Biol. 2011, 17, 3418–3430. [Google Scholar] [CrossRef]
- Harrison, X.A.; Donaldson, L.; Correa-Cano, M.E.; Evans, J.; Fisher, D.N.; Goodwin, C.E.D.; Robinson, B.S.; Hodgson, D.J.; Inger, R. A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 2018, 6, e4794. [Google Scholar] [CrossRef]
- Arnqvist, G. Mixed Models Offer No Freedom from Degrees of Freedom. Trends Ecol. Evol. 2020, 35, 329–335. [Google Scholar] [CrossRef]
- Gelman, A. Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Gessler, A.; Ferrio, J.P.; Hommel, R.; Treydte, K.; Werner, R.A.; Monson, R.K. Stable isotopes in tree rings: Towards a mechanistic understanding of isotope fractionation and mixing processes from the leaves to the wood. Tree Physiol. 2014, 34, 796–818. [Google Scholar] [CrossRef]
- Dolezal, J.; Jandova, V.; Macek, M.; Liancourt, P. Contrasting biomass allocation responses across ontogeny and stress gradients reveal plant adaptations to drought and cold. Funct. Ecol. 2021, 35, 32–42. [Google Scholar] [CrossRef]
- Wang, Y.; Liang, E.; Lu, X.; Camarero, J.J.; Babst, F.; Shen, M.; Peñuelas, J. Warming-induced shrubline advance stalled by moisture limitation on the Tibetan Plateau. Ecography 2021, 44, 1631–1641. [Google Scholar] [CrossRef]
- Ranjitkar, S.; Luedeling, E.; Shrestha, K.K.; Guan, K.; Xu, J. Flowering phenology of tree rhododendron along an elevation gradient in two sites in the Eastern Himalayas. Int. J. Biometeorol. 2013, 57, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems, 2nd ed.; Springer: New York, NY, USA, 2003. [Google Scholar]
- The Intergovernmental Panel on Climate Change. Climate Change 2022: Mitigation of Climate Change. Available online: https://www.ipcc.ch/report/ar6/wg3/. (accessed on 15 October 2022).
- Liang, E.; Leuschner, C.; Dulamsuren, C.; Wagner, B.; Hauck, M. Global warming-related tree growth decline and mortality on the north-eastern Tibetan plateau. Clim. Chang. 2016, 134, 163–176. [Google Scholar] [CrossRef]
- Mou, Y.-M.; Fang, O.; Cheng, X.; Qiu, H. Recent tree growth decline unprecedented over the last four centuries in a Tibetan juniper forest. J. For. Res. 2019, 30, 1429–1436. [Google Scholar] [CrossRef]
- Wang, L.; Chen, W. A CMIP5 multimodel projection of future temperature, precipitation, and climatological drought in China. Int. J. Clim. 2014, 34, 2059–2078. [Google Scholar] [CrossRef]
- Pandey, J.; Sigdel, S.R.; Lu, X.; Salerno, F.; Dawadi, B.; Liang, E.; Camarero, J.J. Early growing-season precipitation drives radial growth of alpine juniper shrubs in the central Himalayas. Geogr. Ann. Ser. A Phys. Geogr. 2020, 102, 317–330. [Google Scholar] [CrossRef]
- Lu, X.; Liang, E.; Babst, F.; Camarero, J.J.; Büntgen, U. Warming-induced tipping points of Arctic and alpine shrub recruitment. Proc. Natl. Acad. Sci. USA 2022, 119, e2118120119. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Time Span | No. Individuals (No. Radii) | Mean Ring Width in the Total Time Span (mm) | Mean Ring Width since 1970 (mm) | Rbar | MS | EPS > 0.85 since |
|---|---|---|---|---|---|---|---|
| NQ | 1855–2020 | 27 (54) | 0.21 ± 0.07 | 0.23 ± 0.04 | 0.61 | 0.42 | 1910 |
| LWQ | 1913–2020 | 26 (52) | 0.21 ± 0.06 | 0.22 ± 0.05 | 0.28 | 0.33 | 1970 |
| Site | Climatic Variables/Month | Controlled Climatic Variable | Partial Correlation |
|---|---|---|---|
| NQ | Tmax/p9 | PDSI_p9 | −0.27 * |
| PDSI/p11 | Tmax_p11 | 0.18 | |
| PDSI/p12 | Tmax_p12 | 0.20 | |
| LWQ | Tmin/c8 | PDSI_c8 | −0.53 ** |
| Tmean/c8 | PDSI_c8 | −0.32 * | |
| PDSI/p9 | Tmax_p9 | 0.20 | |
| PDSI/p10 | Tmax_p10 | 0.31 |
| Name | Climate Variable/Month | R²m | R²c | p | AIC |
|---|---|---|---|---|---|
| LN | SV | 0.12 | 0.14 | <0.01 | 72.21 |
| LWQ | Tmean/c8 | 0.09 | 0.22 | <0.05 | 12.03 |
| Tmin/c8 | 0.18 | 0.30 | <0.01 | 8.19 | |
| PDSI/p9 | 0.12 | 0.24 | <0.05 | 12.56 | |
| PDSI/p10 | 0.09 | 0.22 | <0.05 | 14.35 | |
| NQ | Tmax/p9 | 0.10 | 0.10 | <0.05 | 36.40 |
| PDSI/p11 | 0.03 | 0.03 | >0.05 | 41.90 | |
| PDSI/p12 | 0.03 | 0.03 | >0.05 | 41.68 |
| Forest Region | Species | Length (years) | RW (mm) | GLF |
|---|---|---|---|---|
| Sygera Mts, southeastern TP, China [12] | A | 342 | 0.20 | Low July mean temperature |
| 272 | 0.20 | |||
| 282 | 0.21 | |||
| 117 | 0.18 | |||
| Hengduan Mts, southeastern TP [15] | B | 196 | / | High December–February mean minimum temperature |
| C | 212 | / | ||
| D | 67 | / | ||
| E | 77 | / | ||
| C | 142 | / | ||
| Zhegu Mts, eastern margin of the TP [21] | D | 61 | 0.5 | High January–February mean temperature |
| Central Himalaya, southern margin of the TP [14] | F | 227 | / | Low December–February minimum temperature |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-L.; Wang, Y.-F.; Camarero, J.J. Inconsistent Growth Responses of Alpine Rhododendron Shrubs to Climate Change at Two Sites on the Eastern Tibetan Plateau. Forests 2023, 14, 331. https://doi.org/10.3390/f14020331
Wang Y-L, Wang Y-F, Camarero JJ. Inconsistent Growth Responses of Alpine Rhododendron Shrubs to Climate Change at Two Sites on the Eastern Tibetan Plateau. Forests. 2023; 14(2):331. https://doi.org/10.3390/f14020331
Chicago/Turabian StyleWang, Ya-Lei, Ya-Feng Wang, and Jesús Julio Camarero. 2023. "Inconsistent Growth Responses of Alpine Rhododendron Shrubs to Climate Change at Two Sites on the Eastern Tibetan Plateau" Forests 14, no. 2: 331. https://doi.org/10.3390/f14020331
APA StyleWang, Y.-L., Wang, Y.-F., & Camarero, J. J. (2023). Inconsistent Growth Responses of Alpine Rhododendron Shrubs to Climate Change at Two Sites on the Eastern Tibetan Plateau. Forests, 14(2), 331. https://doi.org/10.3390/f14020331