
Functional Diversity Accelerates the Decomposition of Litter Recalcitrant Carbon but Reduces the Decomposition of Labile Carbon in Subtropical Forests

 and
and
Abstract
:1. Introduction
2. Method
2.1. Experiment Site
2.2. Experimental Design
2.3. Litter Traits Determination
2.4. Data Analysis
3. Results
3.1. The Decomposition Patterns of Litter Mass and Carbon Fractions
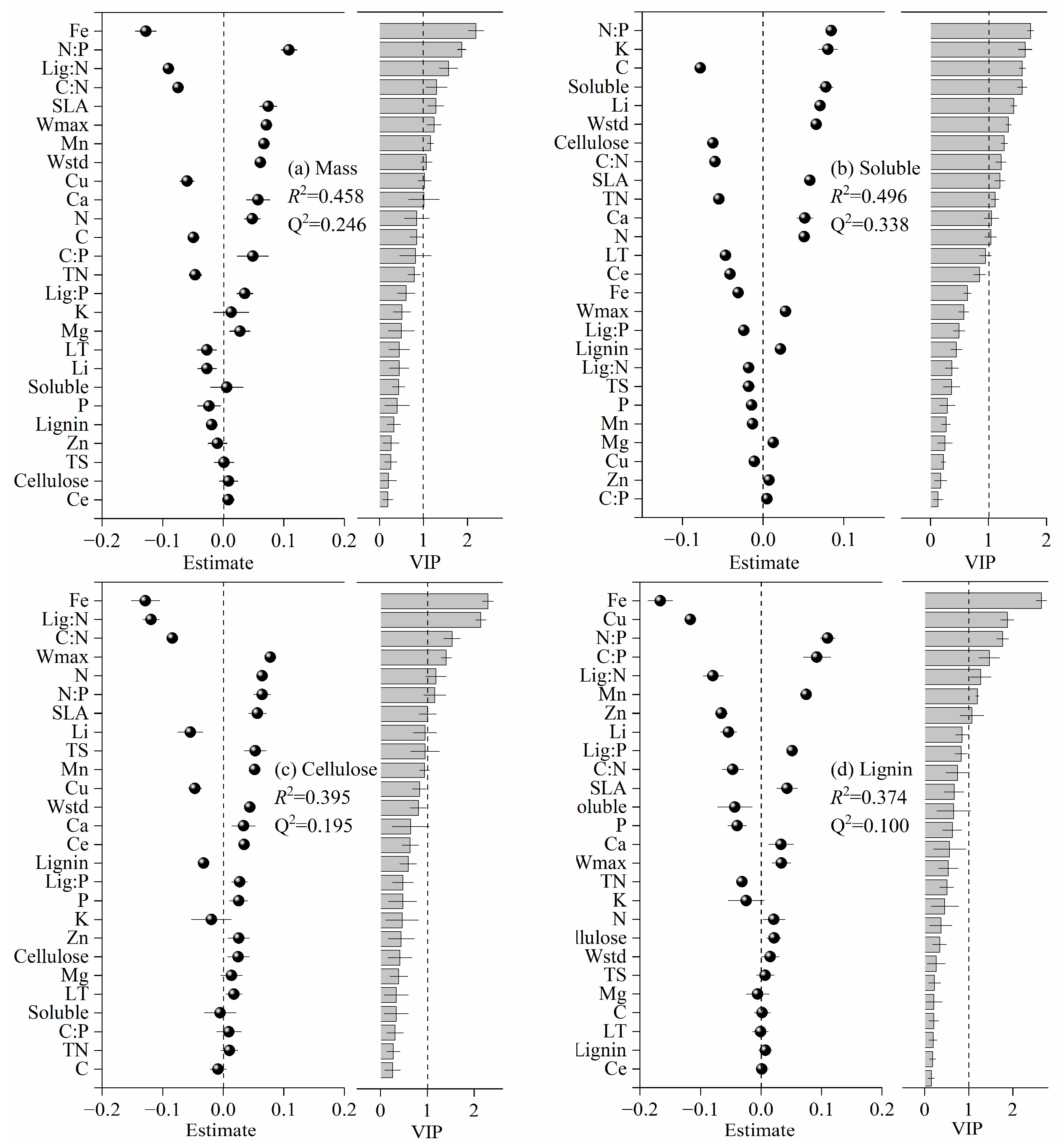
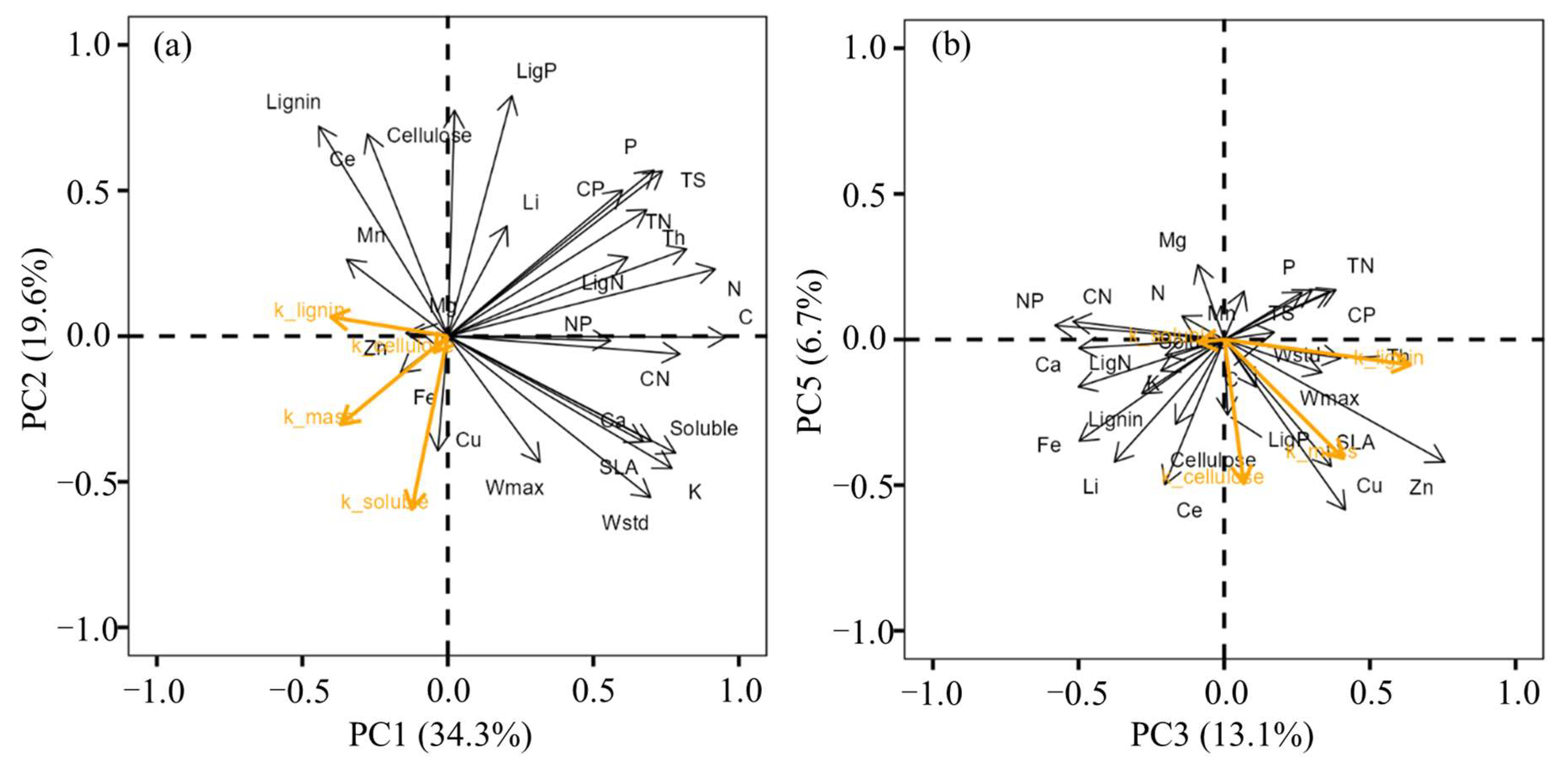
3.2. Relationships between Litter Traits and Decomposition
3.3. The Relative Mixing Effects of Litter Mass and Carbon Fractions
3.4. Relationships between Functional Dissimilarity and RMEs
4. Discussion
4.1. The Decomposition Patterns of Different Carbon Fractions
4.2. Initial Litter Traits Influence the Decomposition Rates of Litter Mass and Carbon Fractions
4.3. The Relative Mixing Effects of Litter Mass and Carbon Fractions
4.4. Functional Dissimilarity may Drive the Mixing Effects of Different Carbon Fractions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vellend, M.; Baeten, L.; Becker-Scarpitta, A.; Boucher-Lalonde, V.; McCune, J.L.; Messier, J.; Myers-Smith, I.H.; Sax, D.F. Plant biodiversity change across scales during the anthropocene. Annu. Rev. Plant Biol. 2017, 68, 563–586. [Google Scholar] [CrossRef] [PubMed]
- Hättenschwiler, S.; Tiunov, A.V.; Scheu, S. Biodiversity and litter decomposition in terrestrial ecosystems. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 191–218. [Google Scholar] [CrossRef]
- Gessner, M.O.; Swan, C.M.; Dang, C.K.; McKie, B.G.; Bardgett, R.D.; Wall, D.H.; Hättenschwiler, S. Diversity meets decomposition. Trends Ecol. Evol. 2010, 25, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Gartner, T.B.; Cardon, Z.G. Decomposition dynamics in mixed-species leaf litter. Oikos 2004, 104, 230–246. [Google Scholar] [CrossRef]
- Kou, L.; Jiang, L.; Hättenschwiler, S.; Zhang, M.; Niu, S.; Fu, X.; Dai, X.; Yan, H.; Li, S.; Wang, H. Diversity-decomposition relationships in forests worldwide. eLife 2020, 9, e55813. [Google Scholar] [CrossRef]
- Mori, A.S.; Cornelissen, J.H.C.; Fujii, S.; Okada, K.-i.; Isbell, F. A meta-analysis on decomposition quantifies afterlife effects of plant diversity as a global change driver. Nat. Commun. 2020, 11, 4547. [Google Scholar] [CrossRef]
- Srivastava, D.S.; Cardinale, B.J.; Downing, A.L.; Duffy, J.E.; Jouseau, C.; Sankaran, M.; Wright, J.P. Diversity has stronger top-down than bottom-up effects on decomposition. Ecology 2009, 90, 1073–1083. [Google Scholar] [CrossRef]
- Grossman, J.J.; Cavender-Bares, J.; Hobbie, S.E. Functional diversity of leaf litter mixtures slows decomposition of labile but not recalcitrant carbon over two years. Ecol. Monogr. 2020, 90, e01407. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Finerty, G.E.; de Bello, F.; Bílá, K.; Berg, M.P.; Dias, A.T.C.; Pezzatti, G.B.; Moretti, M. Exotic or not, leaf trait dissimilarity modulates the effect of dominant species on mixed litter decomposition. J. Ecol. 2016, 104, 1400–1409. [Google Scholar] [CrossRef]
- Purschke, O.; Schmid, B.C.; Sykes, M.T.; Poschlod, P.; Michalski, S.G.; Durka, W.; Kühn, I.; Winter, M.; Prentice, H.C. Contrasting changes in taxonomic, phylogenetic and functional diversity during a long-term succession: Insights into assembly processes. J. Ecol. 2013, 101, 857–866. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Matulich, K.L.; Hooper, D.U.; Byrnes, J.E.; Duffy, E.; Gamfeldt, L.; Balvanera, P.; O’Connor, M.I.; Gonzalez, A. The functional role of producer diversity in ecosystems. Am. J. Bot. 2011, 98, 572–592. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Zeng, D.-H. Functional identity rather than functional diversity or species richness controls litter mixture decomposition in a subtropical forest. Plant Soil 2018, 428, 179–193. [Google Scholar] [CrossRef]
- Tardif, A.; Shipley, B. The relationship between functional dispersion of mixed-species leaf litter mixtures and species’ interactions during decomposition. Oikos 2015, 124, 1050–1057. [Google Scholar] [CrossRef]
- Lecerf, A.; Marie, G.; Kominoski, J.S.; LeRoy, C.J.; Bernadet, C.; Swan, C.M. Incubation time, functional litter diversity, and habitat characteristics predict litter-mixing effects on decomposition. Ecology 2011, 92, 160–169. [Google Scholar] [CrossRef]
- Barantal, S.; Schimann, H.; Fromin, N.; Hättenschwiler, S. C, N and P fertilization in an Amazonian rainforest supports stoichiometric dissimilarity as a driver of litter diversity effects on decomposition. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141682. [Google Scholar] [CrossRef]
- Handa, I.T.; Aerts, R.; Berendse, F.; Berg, M.P.; Bruder, A.; Butenschoen, O.; Chauvet, E.; Gessner, M.O.; Jabiol, J.; Makkonen, M.; et al. Consequences of biodiversity loss for litter decomposition across biomes. Nature 2014, 509, 218–221. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Jørgensen, H.B. Carbon quality rather than stoichiometry controls litter decomposition in a tropical rain forest. J. Ecol. 2010, 98, 754–763. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, Y.; Chen, Y.; Xu, Z.; Zhang, J.; Liu, Y.; Joly, F.-X. Litter diversity accelerates labile carbon but slows recalcitrant carbon decomposition. Soil Biol. Biochem. 2022, 168, 108632. [Google Scholar] [CrossRef]
- Schimel, J.P.; Hättenschwiler, S. Nitrogen transfer between decomposing leaves of different N status. Soil Biol. Biochem. 2007, 39, 1428–1436. [Google Scholar] [CrossRef]
- Makkonen, M.; Berg, M.P.; van Logtestijn, R.S.P.; van Hal, J.R.; Aerts, R. Do physical plant litter traits explain non-additivity in litter mixtures? A test of the improved microenvironmental conditions theory. Oikos 2013, 122, 987–997. [Google Scholar] [CrossRef]
- Kominoski, J.S.; Hoellein, T.J.; Kelly, J.J.; Pringle, C.M. Does mixing litter of different qualities alter stream microbial diversity and functioning on individual litter species? Oikos 2009, 118, 457–463. [Google Scholar] [CrossRef]
- Talbot, J.M.; Treseder, K.K. Interactions among lignin, cellulose, and nitrogen drive litter chemistry–decay relationships. Ecology 2012, 93, 345–354. [Google Scholar] [CrossRef]
- Moorhead, D.L.; Sinsabaugh, R.L. A theoretical model of litter decay and microbial interaction. Ecol. Monogr. 2006, 76, 151–174. [Google Scholar] [CrossRef]
- Shipley, B.; Tardif, A. Causal hypotheses accounting for correlations between decomposition rates of different mass fractions of leaf litter. Ecology 2021, 102, e03196. [Google Scholar] [CrossRef]
- Berg, B.; Matzner, E. Effect of N deposition on decomposition of plant litter and soil organic matter in forest systems. Environ. Rev. Doss. Environ. 1997, 5, 1–25. [Google Scholar] [CrossRef]
- Berg, B. Decomposition patterns for foliar litter—A theory for influencing factors. Soil Biol. Biochem. 2014, 78, 222–232. [Google Scholar] [CrossRef]
- Adair, E.C.; Parton, W.J.; Del Grosso, S.J.; Silver, W.L.; Harmon, M.E.; Hall, S.A.; Burke, I.C.; Hart, S.C. Simple three-pool model accurately describes patterns of long-term litter decomposition in diverse climates. Glob. Chang. Biol. 2008, 14, 2636–2660. [Google Scholar] [CrossRef]
- Butenschoen, O.; Krashevska, V.; Maraun, M.; Marian, F.; Sandmann, D.; Scheu, S. Litter mixture effects on decomposition in tropical montane rainforests vary strongly with time and turn negative at later stages of decay. Soil Biol. Biochem. 2014, 77, 121–128. [Google Scholar] [CrossRef]
- Preston, C.M.; Nault, J.R.; Trofymow, J.A. Chemical changes during 6 years of decomposition of 11 litters in some canadian forest sites. Part 2. 13C abundance, solid-state 13C NMR spectroscopy and the meaning of “Lignin”. Ecosystems 2009, 12, 1078–1102. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Y.; Zhou, Y.; Zheng, H.; Xu, Z.; Tan, B.; You, C.; Zhang, L.; Li, H.; Guo, L.; et al. Litter chemical traits strongly drove the carbon fractions loss during decomposition across an alpine treeline ecotone. Sci. Total Environ. 2021, 753, 142287. [Google Scholar] [CrossRef] [PubMed]
- Makkonen, M.; Berg, M.P.; Handa, I.T.; Hättenschwiler, S.; van Ruijven, J.; van Bodegom, P.M.; Aerts, R. Highly consistent effects of plant litter identity and functional traits on decomposition across a latitudinal gradient. Ecol. Lett. 2012, 15, 1033–1041. [Google Scholar] [CrossRef]
- Terashima, N.; Kitano, K.; Kojima, M.; Yoshida, M.; Yamamoto, H.; Westermark, U. Nanostructural assembly of cellulose, hemicellulose, and lignin in the middle layer of secondary wall of ginkgo tracheid. J. Wood Sci. 2009, 55, 409–416. [Google Scholar] [CrossRef]
- Ververis, C.; Georghiou, K.; Christodoulakis, N.; Santas, P.; Santas, R. Fiber dimensions, lignin and cellulose content of various plant materials and their suitability for paper production. Ind. Crop. Prod. 2004, 19, 245–254. [Google Scholar] [CrossRef]
- Rao, C.R. Diversity and dissimilarity coefficients: A unified approach. Theor. Popul. Biol. 1982, 21, 24–43. [Google Scholar] [CrossRef]
- Wider, R.K.; Lang, G.E. A Critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology 1982, 63, 1636–1642. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bonner, K.I.; Nicholson, K.S. Biodiversity and plant litter: Experimental evidence which does not support the view that enhanced species richness improves ecosystem function. Oikos 1997, 79, 247–258. [Google Scholar] [CrossRef]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Trap, J.; Akpa-Vinceslas, M.; Margerie, P.; Boudsocq, S.; Richard, F.; Decaëns, T.; Aubert, M. Slow decomposition of leaf litter from mature Fagus sylvatica trees promotes offspring nitrogen acquisition by interacting with ectomycorrhizal fungi. J. Ecol. 2017, 105, 528–539. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Aber, J.D.; Melillo, J.M.; McClaugherty, C.A. Predicting long-term patterns of mass-loss, nitrogen dynamics, and soil organic-matter formation from initial fine litter chemistry in temperate forest ecosystems. Can. J. Bot.-Rev. Can. Bot. 1990, 68, 2201–2208. [Google Scholar] [CrossRef]
- Berg, B.; Hannus, K.; Popoff, T.; Theander, O. Changes in organic-chemical components of needle litter during decomposition—Long-term decomposition in a scots pine forest.1. Can. J. Bot.-Rev. Can. Bot. 1982, 60, 1310–1319. [Google Scholar] [CrossRef]
- Berg, B.; Agren, G.I. Decomposition of needle litter and its organic-chemical components—Theory and field experiments—Long-term decomposition in a scots pine forest.3. Can. J. Bot.-Rev. Can. Bot. 1984, 62, 2880–2888. [Google Scholar] [CrossRef]
- Djukic, I.; Kepfer-Rojas, S.; Schmidt, I.K.; Larsen, K.S.; Beier, C.; Berg, B.; Verheyen, K.; Caliman, A.; Paquette, A.; Gutiérrez-Girón, A.; et al. Early stage litter decomposition across biomes. Sci. Total Environ. 2018, 628–629, 1369–1394. [Google Scholar] [CrossRef]
- Parton, W.; Silver, W.L. Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 2007, 315, 361–364. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, J.H.C.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N.; et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.R.; Parkinson, D.; Parsons, W.F.J. Nitrogen and lignin content as predictors of litter decay rates: A microcosm test. Ecology 1989, 70, 97–104. [Google Scholar] [CrossRef]
- Agethen, S.; Knorr, K.-H. Juncus effusus mono-stands in restored cutover peat bogs—Analysis of litter quality, controls of anaerobic decomposition, and the risk of secondary carbon loss. Soil Biol. Biochem. 2018, 117, 139–152. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, L.; Chen, Y.; Zhang, J.; Liu, Y. Litter stoichiometric traits have stronger impact on humification than environment conditions in an alpine treeline ecotone. Plant Soil 2020, 453, 545–560. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J.; Modrzynski, J.; Mrozinski, P.; Hobbie, S.E.; Eissenstat, D.M.; Chorover, J.; Chadwick, O.A.; Hale, C.M.; Tjoelker, M.G. Linking litter calcium, earthworms and soil properties: A common garden test with 14 tree species. Ecol. Lett. 2005, 8, 811–818. [Google Scholar] [CrossRef]
- Norton, R.A.; Behanpelletier, V.M. Calcium-carbonate and calcium-oxalate as cuticular hardening agents in oribatid mites (acari, oribatida). Can. J. Zool.-Rev. Can. Zool. 1991, 69, 1504–1511. [Google Scholar] [CrossRef]
- Berg, B.; Ekbohm, G.; Johansson, M.B.; McClaugherty, C.; Rutigliano, F.; DeSanto, A.V. Maximum decomposition limits of forest litter types: A synthesis. Can. J. Bot.-Rev. Can. Bot. 1996, 74, 659–672. [Google Scholar] [CrossRef]
- Berg, B.; Davey, M.P.; De Marco, A.; Emmett, B.; Faituri, M.; Hobbie, S.E.; Johansson, M.B.; Liu, C.; McClaugherty, C.; Norell, L.; et al. Factors influencing limit values for pine needle litter decomposition: A synthesis for boreal and temperate pine forest systems. Biogeochemistry 2010, 100, 57–73. [Google Scholar] [CrossRef]
- Sun, T.; Cui, Y.; Berg, B.; Zhang, Q.; Dong, L.; Wu, Z.; Zhang, L. A test of manganese effects on decomposition in forest and cropland sites. Soil Biol. Biochem. 2019, 129, 178–183. [Google Scholar] [CrossRef]
- Berg, B.; Erhagen, B.; Johansson, M.-B.; Nilsson, M.; Stendahl, J.; Trum, F.; Vesterdal, L. Manganese in the litter fall-forest floor continuum of boreal and temperate pine and spruce forest ecosystems—A review. For. Ecol. Manag. 2015, 358, 248–260. [Google Scholar] [CrossRef]
- Perez, J.; Jeffries, T.W. Roles of manganese and organic-acid chelators in regulating lignin degradation and biosynthesis of peroxidases by phanerochaete-chrysosporium. Appl. Environ. Microbiol. 1992, 58, 2402–2409. [Google Scholar] [CrossRef]
- Hofrichter, M. Review: Lignin conversion by manganese peroxidase (MnP). Enzym. Microb. Technol. 2002, 30, 454–466. [Google Scholar] [CrossRef]
- Freedman, B.; Hutchinson, T.C. Effects of smelter pollutants on forest leaf litter decomposition near a nickel-copper smelter at sudbury, ontario. Can. J. Bot.-Rev. Can. Bot. 1980, 58, 1722–1736. [Google Scholar] [CrossRef]
- Hall, S.J.; Silver, W.L.; Timokhin, V.I.; Hammel, K.E. Iron addition to soil specifically stabilized lignin. Soil Biol. Biochem. 2016, 98, 95–98. [Google Scholar] [CrossRef]
- Hall, S.J.; Silver, W.L.; Timokhin, V.I.; Hammel, K.E. Lignin decomposition is sustained under fluctuating redox conditions in humid tropical forest soils. Glob. Chang. Biol. 2015, 21, 2818–2828. [Google Scholar] [CrossRef]
- Eusterhues, K.; Neidhardt, J.; Haedrich, A.; Kuesel, K.; Totsche, K.U. Biodegradation of ferrihydrite-associated organic matter. Biogeochemistry 2014, 119, 45–50. [Google Scholar] [CrossRef]
- Liao, C.; Huang, W.; Wells, J.; Zhao, R.; Allen, K.; Hou, E.; Huang, X.; Qiu, H.; Tao, F.; Jiang, L.; et al. Microbe-iron interactions control lignin decomposition in soil. Soil Biol. Biochem. 2022, 173, 108803. [Google Scholar] [CrossRef]
- Riedel, T.; Zak, D.; Biester, H.; Dittmar, T. Iron traps terrestrially derived dissolved organic matter at redox interfaces. Proc. Natl. Acad. Sci. USA 2013, 110, 10101–10105. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, H.; He, J.-S.; Feng, X. Iron-mediated soil carbon response to water-table decline in an alpine wetland. Nat. Commun. 2017, 8, 15972. [Google Scholar] [CrossRef]
- Liu, G.; Wang, L.; Jiang, L.; Pan, X.; Huang, Z.; Dong, M.; Cornelissen, J.H.C. Specific leaf area predicts dryland litter decomposition via two mechanisms. J. Ecol. 2018, 106, 218–229. [Google Scholar] [CrossRef]
- Vaieretti, M.V.; Harguindeguy, N.P.; Gurvich, D.E.; Cingolani, A.M.; Cabido, M. Decomposition dynamics and physico-chemical leaf quality of abundant species in a montane woodland in central argentina. Plant Soil 2005, 278, 223–234. [Google Scholar] [CrossRef]
- Berg, B.; Mcclaugherty, C. Plant Litter. Decomposition, Humus Formation, Carbon Sequestration; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Kuzyakov, Y.; Friedel, J.K.; Stahr, K. Review of mechanisms and quantification of priming effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Tunlid, A. Ectomycorrhizal fungi—Potential organic matter decomposers, yet not saprotrophs. New Phytol. 2015, 205, 1443–1447. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Litter Mixtures | SR | FDis | Mass Loss | REMs | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | Soluble | Cellulose | Lignin | Total | Soluble | Cellulose | Lignin | ||||
| M1 | Qg + Cs | 2 | 2.70 | 42.89 | 54.83 | 51.198 | 22.78 | 15.81 | −20.20 | 5.90 | 119.84 |
| M2 | Qg + Pb | 2 | 2.93 | 36.54 | 44.09 | 40.003 | 29.08 | 20.14 | −33.71 | 8.89 | 159.76 |
| M3 | Ss + Cs | 2 | 2.74 | 43.42 | 47.12 | 49.750 | 30.71 | 4.16 | −30.62 | −10.84 | 150.94 |
| M4 | Qg + Qr | 2 | 3.49 | 44.96 | 53.00 | 48.621 | 36.98 | 38.26 | −14.94 | 8.62 | 255.00 |
| M5 | Lf + Pb | 2 | 3.07 | 34.38 | 64.57 | 37.657 | 10.75 | −7.97 | −10.17 | −13.59 | −34.52 |
| M6 | Lf + Qg | 2 | 3.80 | 43.25 | 61.79 | 39.687 | 34.18 | −3.10 | −18.82 | −26.74 | 119.43 |
| M7 | Qg + Cs + Pb | 3 | 3.37 | 35.08 | 47.42 | 36.213 | 26.67 | 8.28 | −28.29 | −13.54 | 141.81 |
| M8 | Ss + Qg + Cs | 3 | 3.20 | 48.96 | 62.75 | 53.223 | 34.60 | 21.32 | −9.43 | 0.48 | 201.41 |
| M9 | Ss + Qg + Pb | 3 | 3.63 | 38.42 | 52.19 | 40.323 | 29.17 | 6.88 | −23.09 | −12.06 | 144.94 |
| M10 | Pm + Qg + Qr | 3 | 4.75 | 43.42 | 50.84 | 48.812 | 33.87 | 16.19 | −23.14 | −1.13 | 185.47 |
| M11 | Pm + Lf + Qg | 3 | 4.90 | 43.04 | 59.48 | 52.162 | 22.96 | −5.29 | −20.43 | −7.23 | 33.21 |
| M12 | Lf + Qg + Qr | 3 | 4.06 | 48.04 | 57.82 | 56.921 | 32.27 | 23.61 | −17.32 | 13.59 | 173.30 |
| M13 | Ss + Qg + Cs + Pb | 4 | 3.67 | 36.08 | 46.62 | 44.384 | 19.18 | 0.09 | −30.66 | −5.65 | 64.60 |
| M14 | Pm + Qg + Cs + Pb | 4 | 4.31 | 39.63 | 52.22 | 48.575 | 21.55 | 9.87 | −22.97 | 6.14 | 58.46 |
| M15 | Pm + Ss + Qg + Cs | 4 | 4.29 | 42.96 | 32.68 | 51.577 | 37.99 | 2.20 | −53.33 | −5.41 | 171.35 |
| M16 | Pm + Lf + Qr + Pb | 4 | 4.60 | 37.58 | 46.34 | 44.346 | 25.46 | 0.83 | −31.90 | −6.40 | 75.92 |
| M17 | Lf + Qg + Qr + Pb | 4 | 3.97 | 39.78 | 53.03 | 47.836 | 23.45 | 13.87 | −15.95 | 8.19 | 97.36 |
| M18 | Pm + Lf + Qg + Qr | 4 | 4.93 | 42.21 | 55.12 | 51.496 | 24.67 | 3.16 | −21.74 | −1.92 | 75.61 |
| M19 | Pm + Ss + Qg + Cs + Pb | 5 | 4.38 | 39.50 | 44.69 | 49.373 | 24.41 | 3.26 | −34.57 | 0.19 | 79.82 |
| M20 | Qr + Ss + Qg + Cs + Pb | 5 | 3.94 | 41.46 | 48.16 | 50.236 | 26.54 | 20.85 | −25.18 | 8.57 | 161.45 |
| M21 | Lf + Ss + Qg + Cs + Pb | 5 | 4.09 | 42.92 | 47.24 | 50.506 | 30.61 | 9.61 | −32.96 | 1.63 | 125.16 |
| M22 | Pm + Lf + Qg + Qr + Cs | 5 | 4.86 | 39.00 | 45.34 | 49.143 | 22.74 | −2.52 | −34.79 | −5.18 | 68.05 |
| M23 | Pm + Lf + Ss + Qg + Qr | 5 | 4.79 | 46.50 | 48.74 | 50.939 | 40.30 | 10.36 | −30.79 | −7.07 | 189.12 |
| M24 | Pm + Lf + Qg + Qr + Pb | 5 | 4.66 | 39.13 | 51.37 | 46.964 | 23.87 | 4.72 | −25.30 | −0.45 | 75.03 |
| M25 | Pm + Lf + Ss + Qg + Qr + Cs | 6 | 4.72 | 44.50 | 47.70 | 48.782 | 37.30 | 8.08 | −31.52 | −9.27 | 175.82 |
| M26 | Pm + Lf + Ss + Qg + Qr + Pb | 6 | 4.68 | 38.33 | 42.19 | 46.290 | 27.48 | −1.63 | −38.88 | −7.54 | 101.89 |
| M27 | Pm + Lf + Ss + Qg + Cs + Pb | 6 | 4.61 | 42.38 | 44.94 | 50.385 | 31.21 | 4.70 | −36.46 | −1.69 | 110.29 |
| M28 | Pm + Lf + Ss + Qr + Cs + Pb | 6 | 4.69 | 42.44 | 49.69 | 46.902 | 32.70 | 9.55 | −26.93 | −7.36 | 136.37 |
| M29 | Pm + Lf + Qg + Qr + Cs + Pb | 6 | 4.67 | 43.83 | 50.19 | 51.265 | 34.34 | 17.85 | −26.46 | 7.37 | 158.85 |
| M30 | Pm + Ss + Qg + Qr + Cs + Pb | 6 | 4.50 | 34.17 | 26.99 | 41.274 | 29.36 | −6.22 | −59.04 | −14.43 | 144.44 |
| M31 | Lf + Ss + Qg + Qr + Cs + Pb | 6 | 4.19 | 38.67 | 52.40 | 46.048 | 21.35 | 3.99 | −22.84 | −5.19 | 78.12 |
| M32 | Pm + Lf + Ss + Qg + Qr + Cs + Pb | 7 | 4.64 | 40.04 | 53.02 | 45.733 | 25.60 | 3.75 | −22.65 | −8.58 | 92.50 |
| Tree Species | Mass Loss (%) | |||
|---|---|---|---|---|
| Total Mass | Soluble | Cellulose | Lignin | |
| Pinus massoniana | 47.1(0.43) a | 73.1(0.19) a | 62.8(0.52) a | 22.9(0.56) a |
| Liquidambar formosana | 51.6(6.15) a | 79.1(3.01) ab | 66.2(4.40) a | 28.9(6.06) a |
| Schima superba | 47.0(1.51) a | 70.5(1.58) bc | 64.6(0.74) a | 15.9(2.44) a |
| Quercus glauca | 37.7(1.86) b | 76.6(1.13) cd | 47.5(1.93) b | 17.4(1.99) a |
| Quercus rubra | 27.3(1.47) b | 54.9(3.00) de | 47.1(0.55) b | 18.2(2.91) a |
| Castanopsis sclerophylla | 36.4(0.48) c | 66.8(0.55) e | 50.5(1.80) b | 17.9(2.06) a |
| Phoebe bournei | 23.1(0.81) c | 62.4(1.21) f | 27.3(1.58) c | 14.9(9.73) a |
| Decomposition Type | Additive | Synergistic | Antagonistic |
|---|---|---|---|
| Total mass | 6 | 23 | 3 |
| Soluble | 0 | 0 | 32 |
| Cellulose | 9 | 9 | 14 |
| Lignin | 0 | 31 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, G.; Wan, J.; Gu, Z.; Ding, W.; Hu, S.; Du, Q.; Meng, S.; Yang, C. Functional Diversity Accelerates the Decomposition of Litter Recalcitrant Carbon but Reduces the Decomposition of Labile Carbon in Subtropical Forests. Forests 2023, 14, 2258. https://doi.org/10.3390/f14112258
Zhou G, Wan J, Gu Z, Ding W, Hu S, Du Q, Meng S, Yang C. Functional Diversity Accelerates the Decomposition of Litter Recalcitrant Carbon but Reduces the Decomposition of Labile Carbon in Subtropical Forests. Forests. 2023; 14(11):2258. https://doi.org/10.3390/f14112258
Chicago/Turabian StyleZhou, Guang, Jing Wan, Zhenjun Gu, Wei Ding, Shan Hu, Qiang Du, Shengwang Meng, and Chunxia Yang. 2023. "Functional Diversity Accelerates the Decomposition of Litter Recalcitrant Carbon but Reduces the Decomposition of Labile Carbon in Subtropical Forests" Forests 14, no. 11: 2258. https://doi.org/10.3390/f14112258
APA StyleZhou, G., Wan, J., Gu, Z., Ding, W., Hu, S., Du, Q., Meng, S., & Yang, C. (2023). Functional Diversity Accelerates the Decomposition of Litter Recalcitrant Carbon but Reduces the Decomposition of Labile Carbon in Subtropical Forests. Forests, 14(11), 2258. https://doi.org/10.3390/f14112258