
Response of a Sylvan Moss Species (Didymodon validus Limpr.) with a Narrow Distribution Range to Climate Change



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Traits
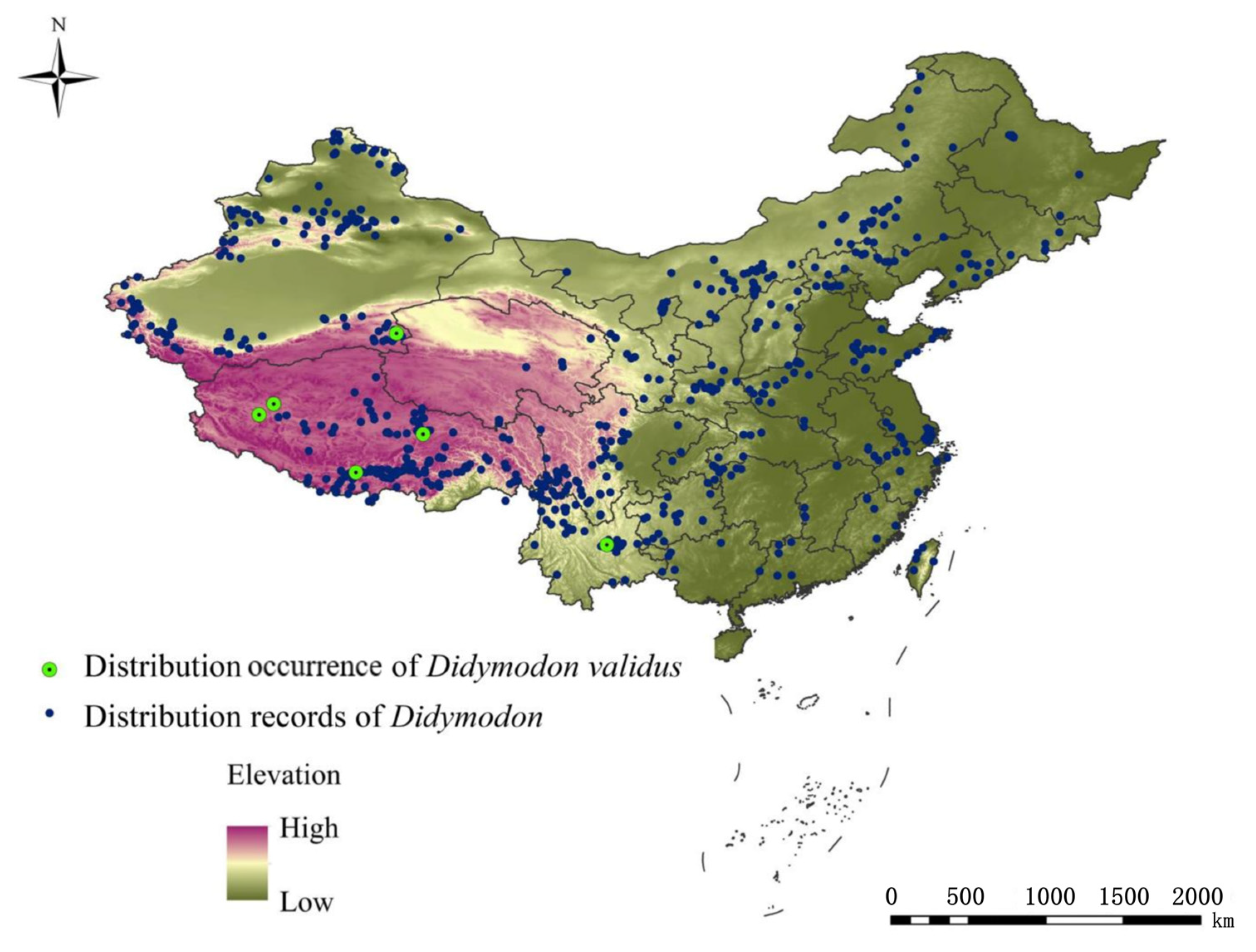
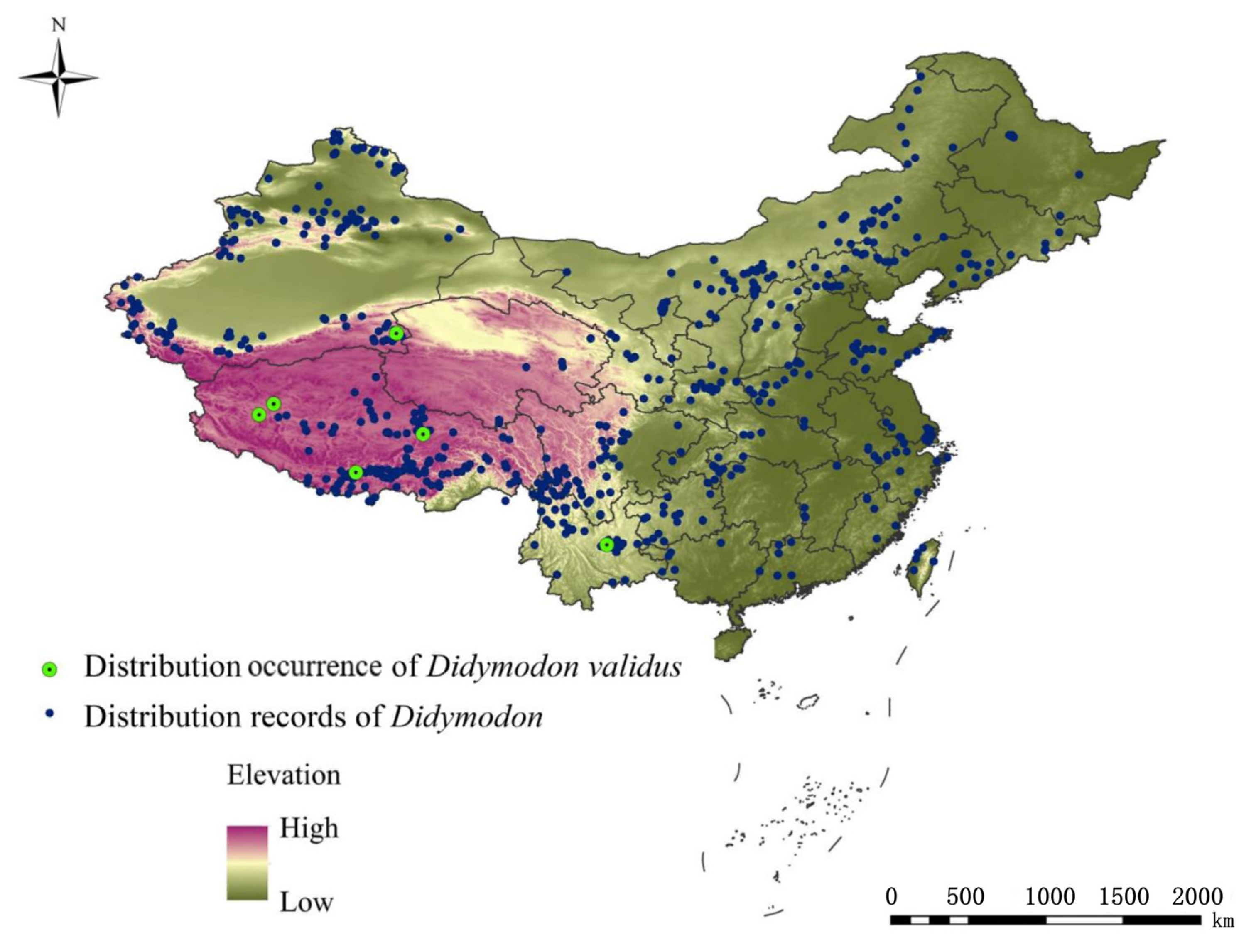
2.2. Study Area and Species Occurrence
2.3. Environmental Variables and Climate Change Scenarios
2.4. Distribution Modeling
2.5. Model Evaluation
3. Results
3.1. Morphological Characteristics
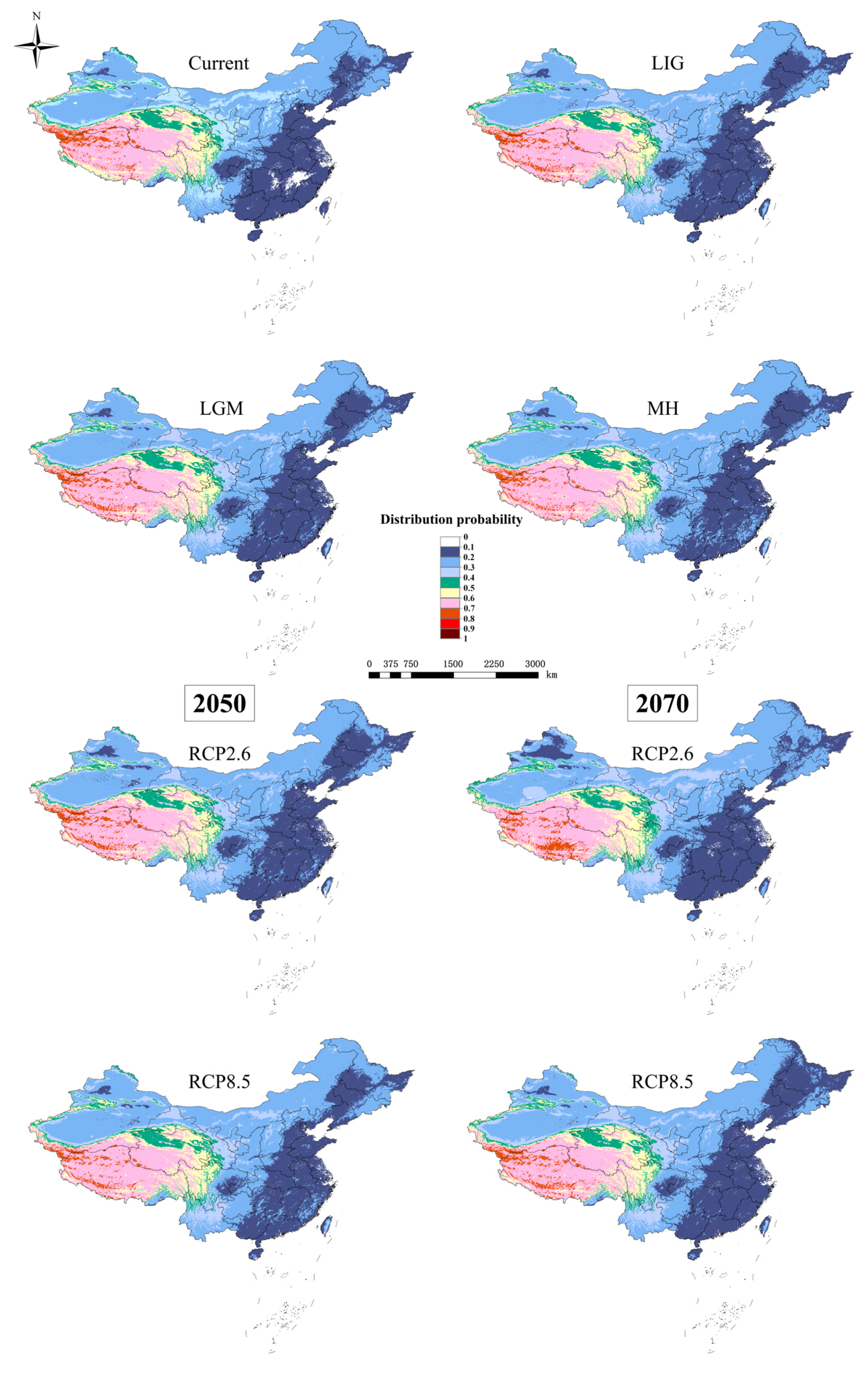
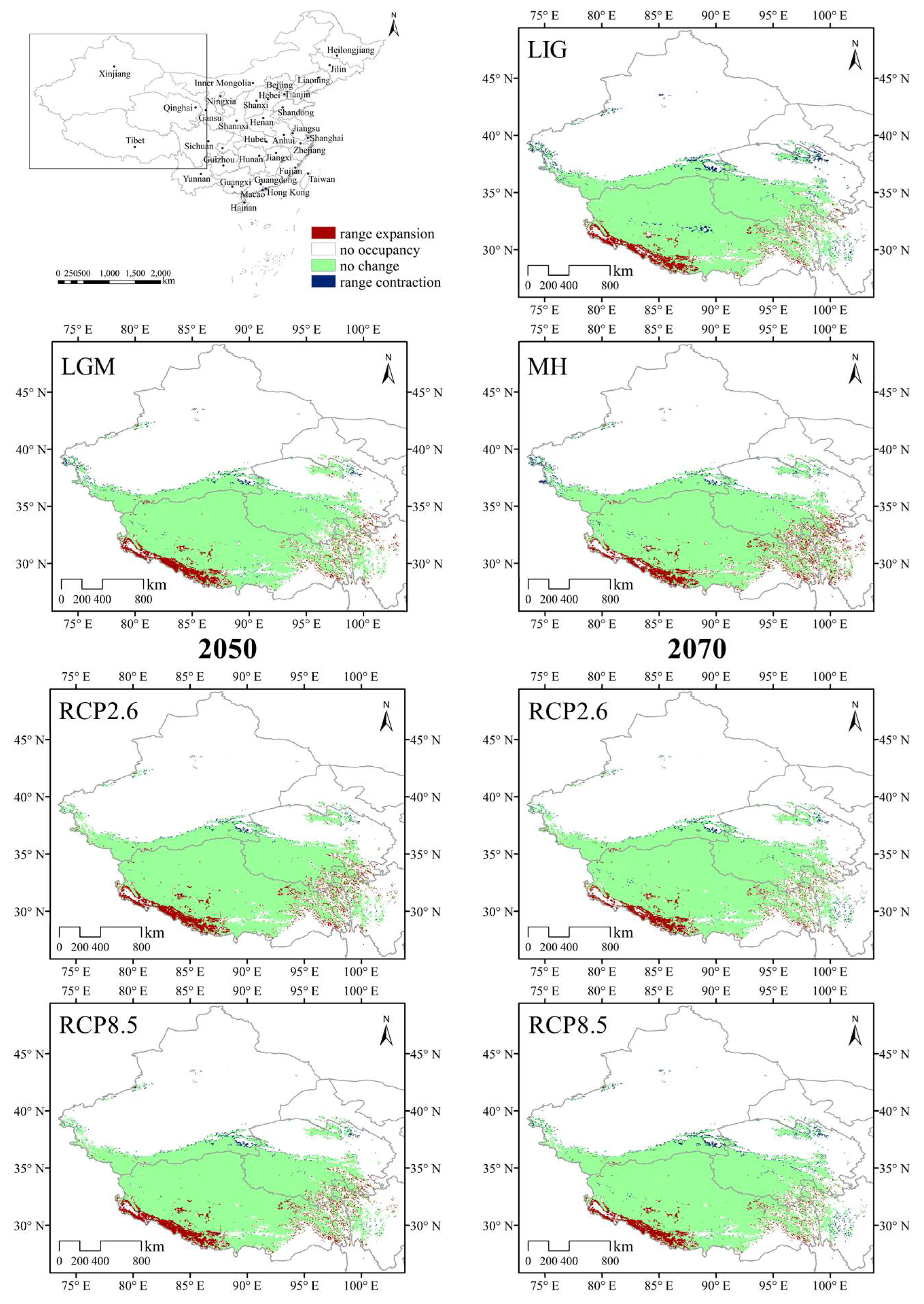
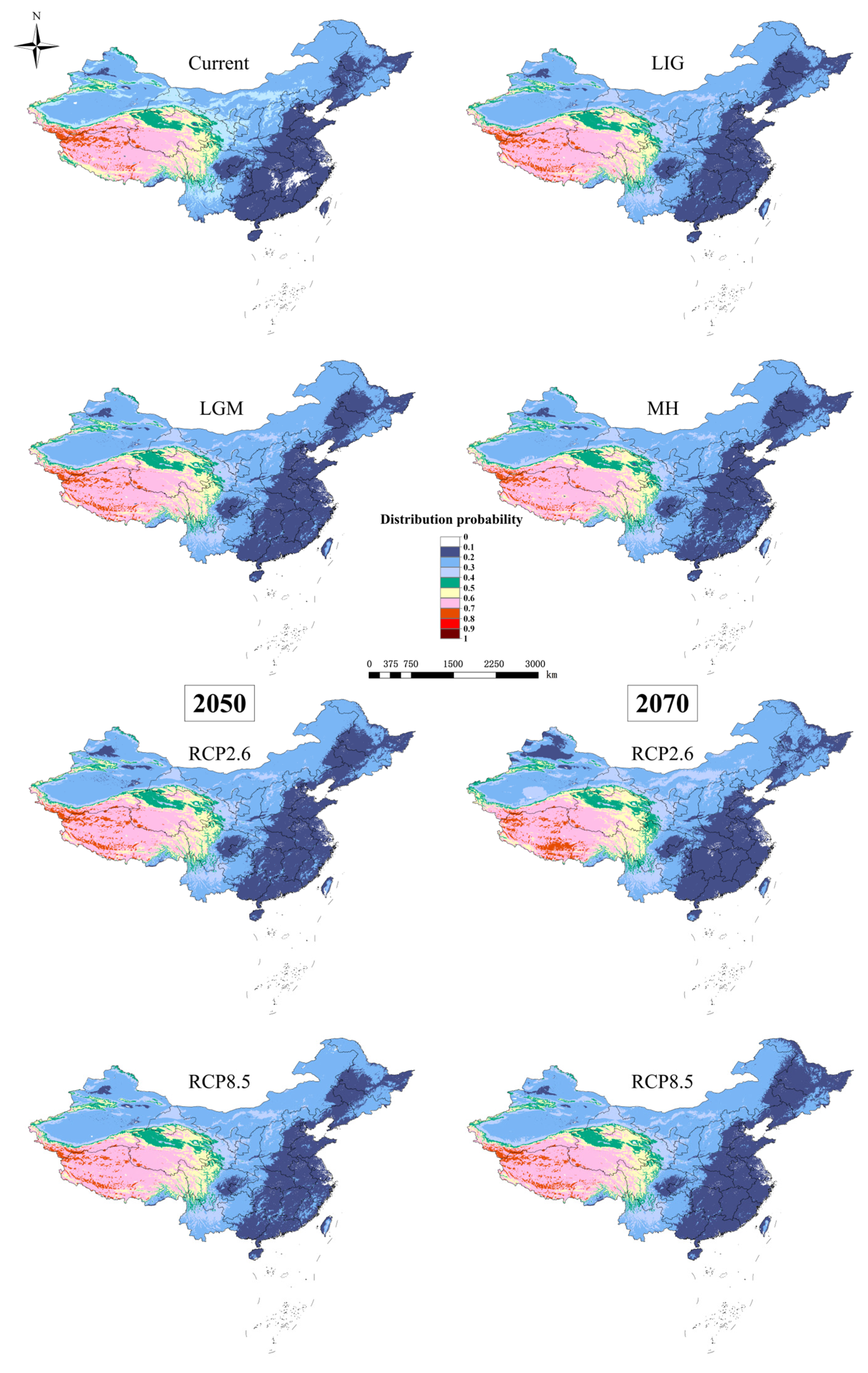
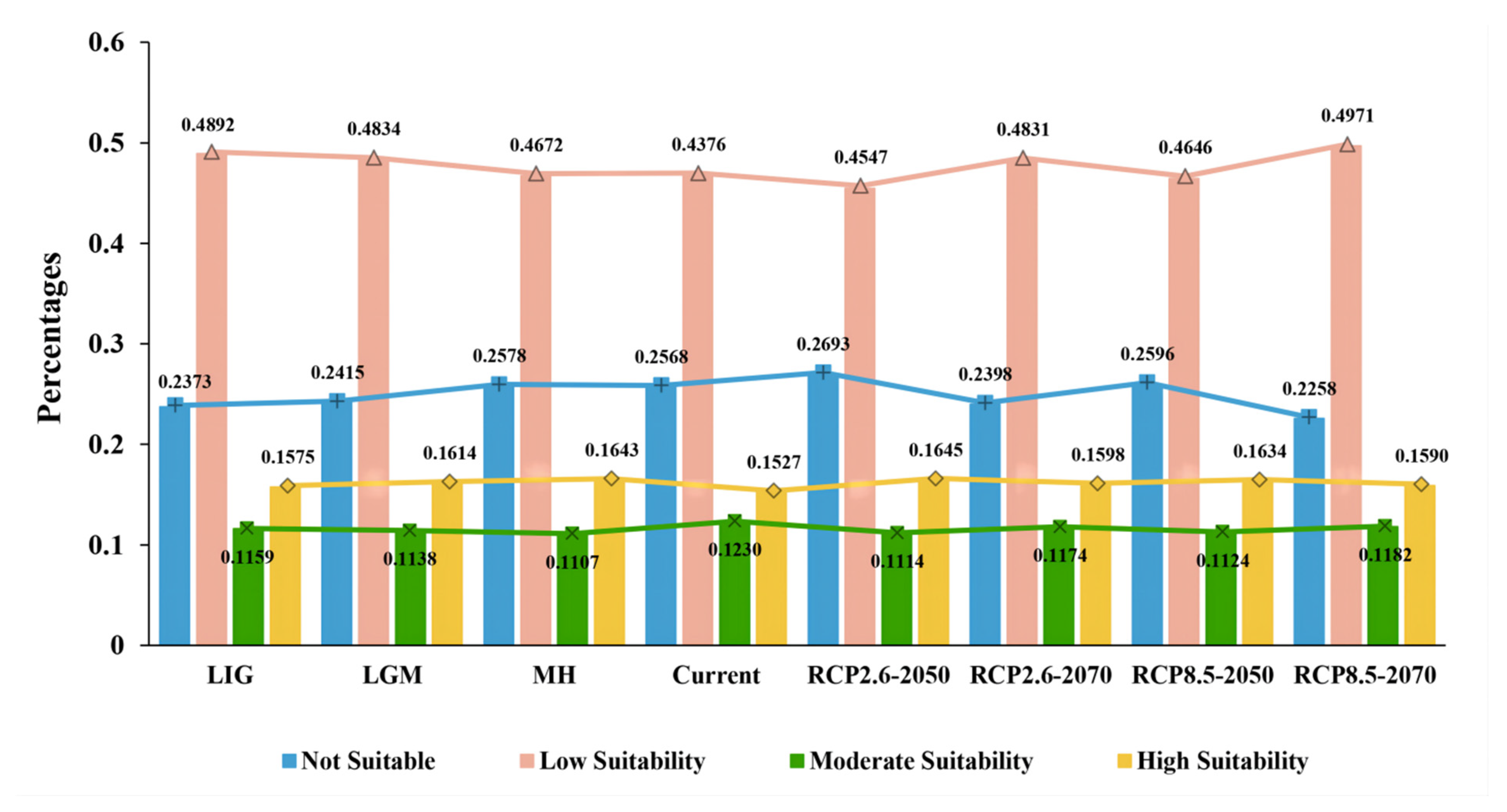
3.2. Potential Distribution Pattern of D. validus
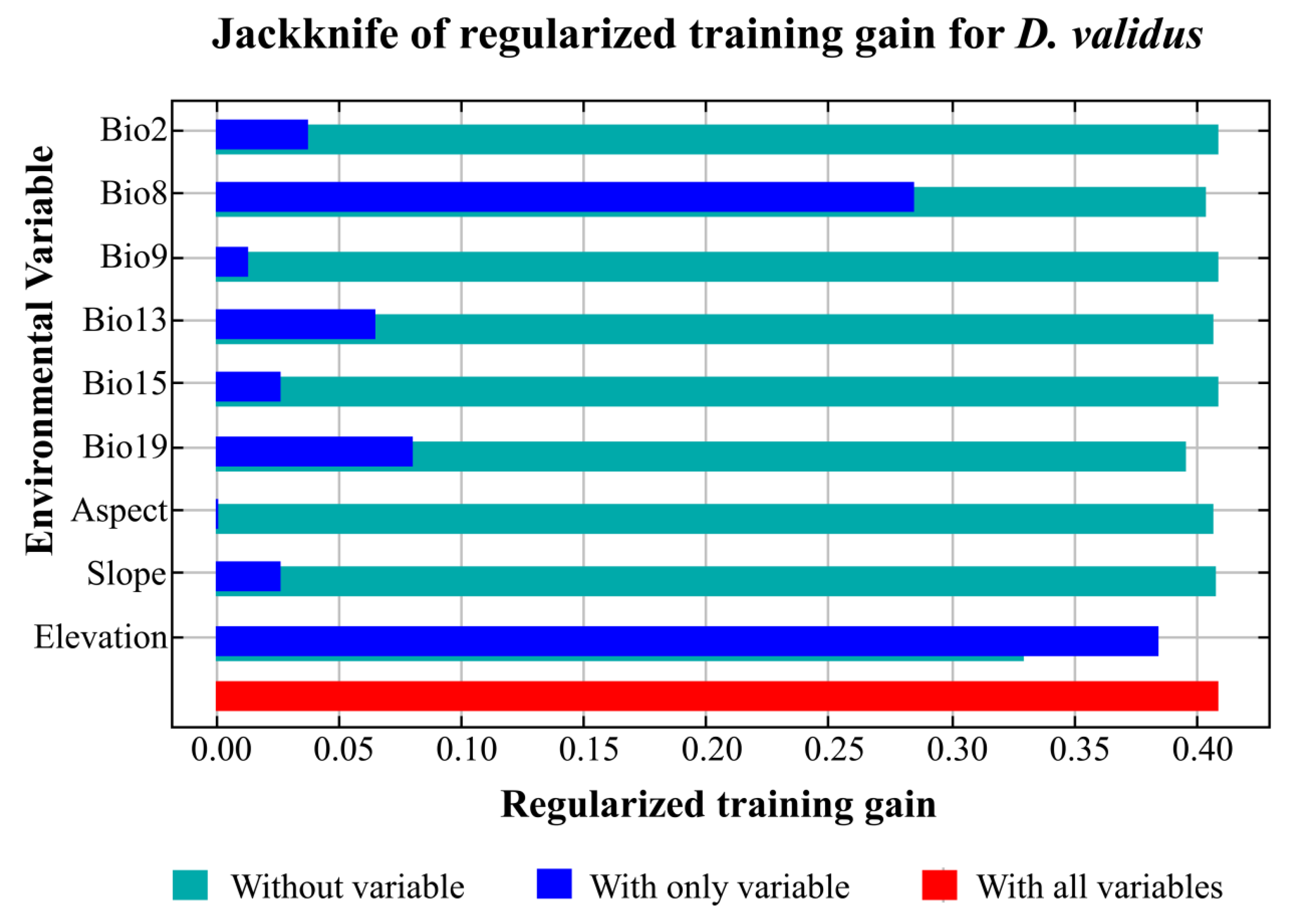
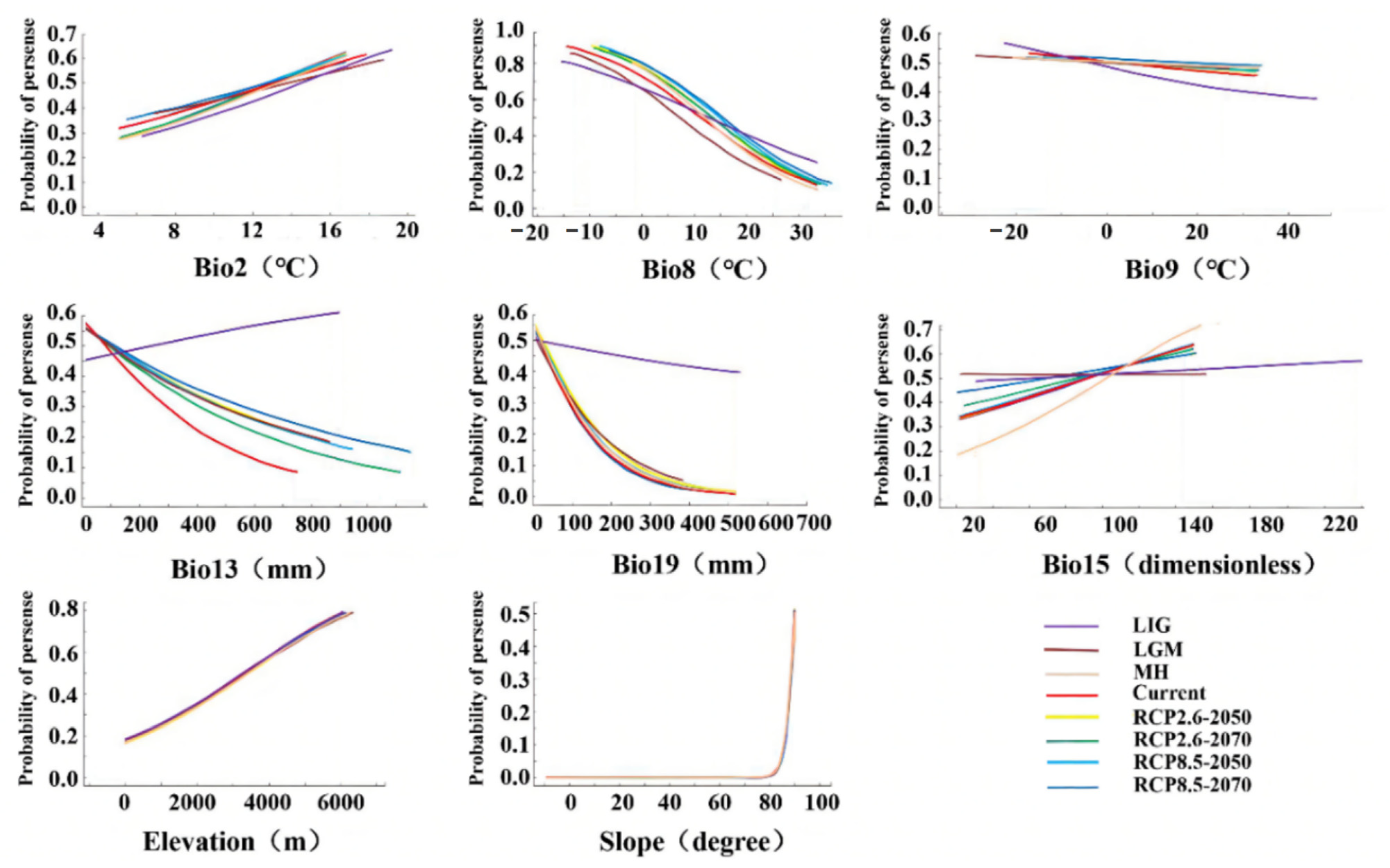
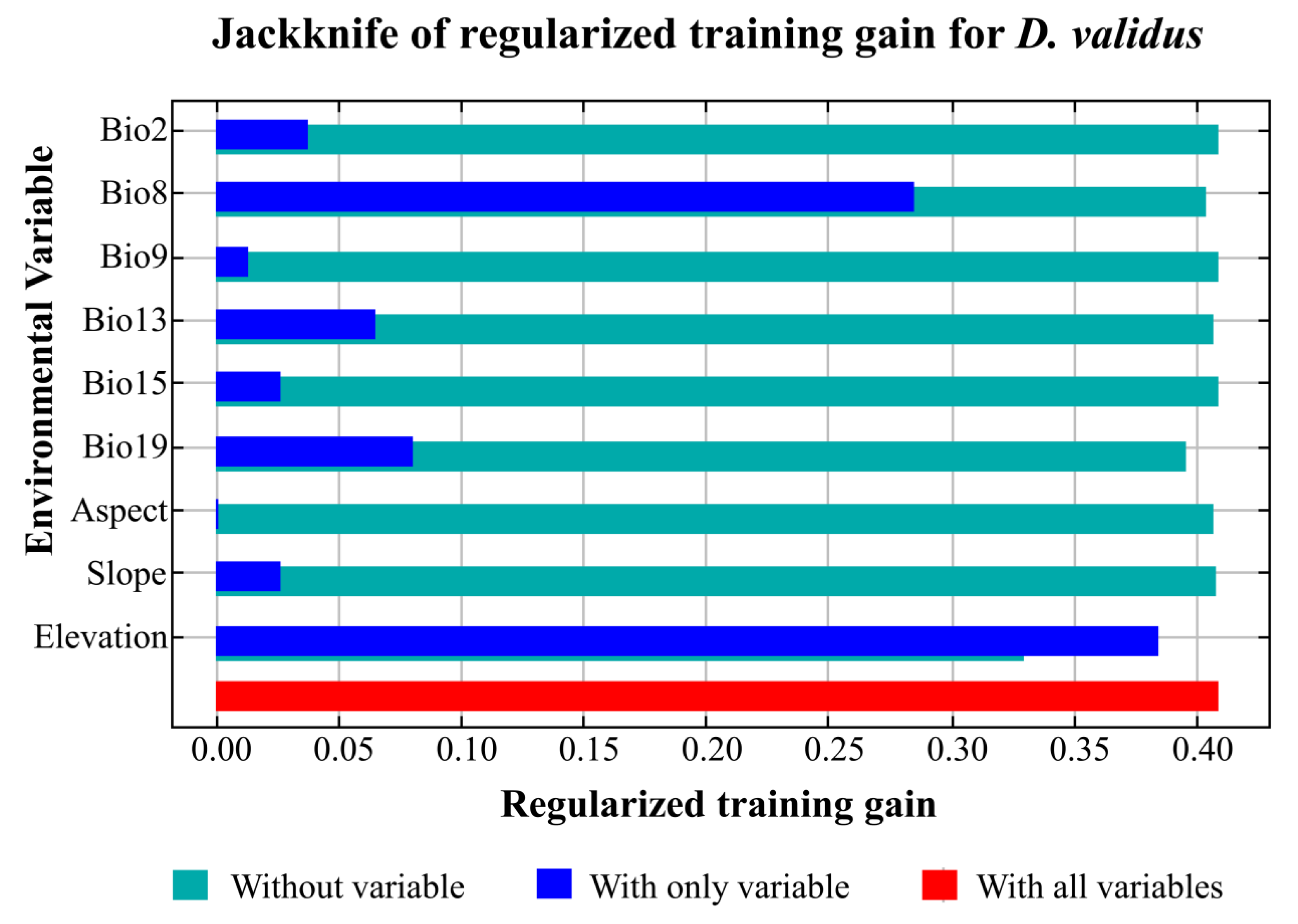
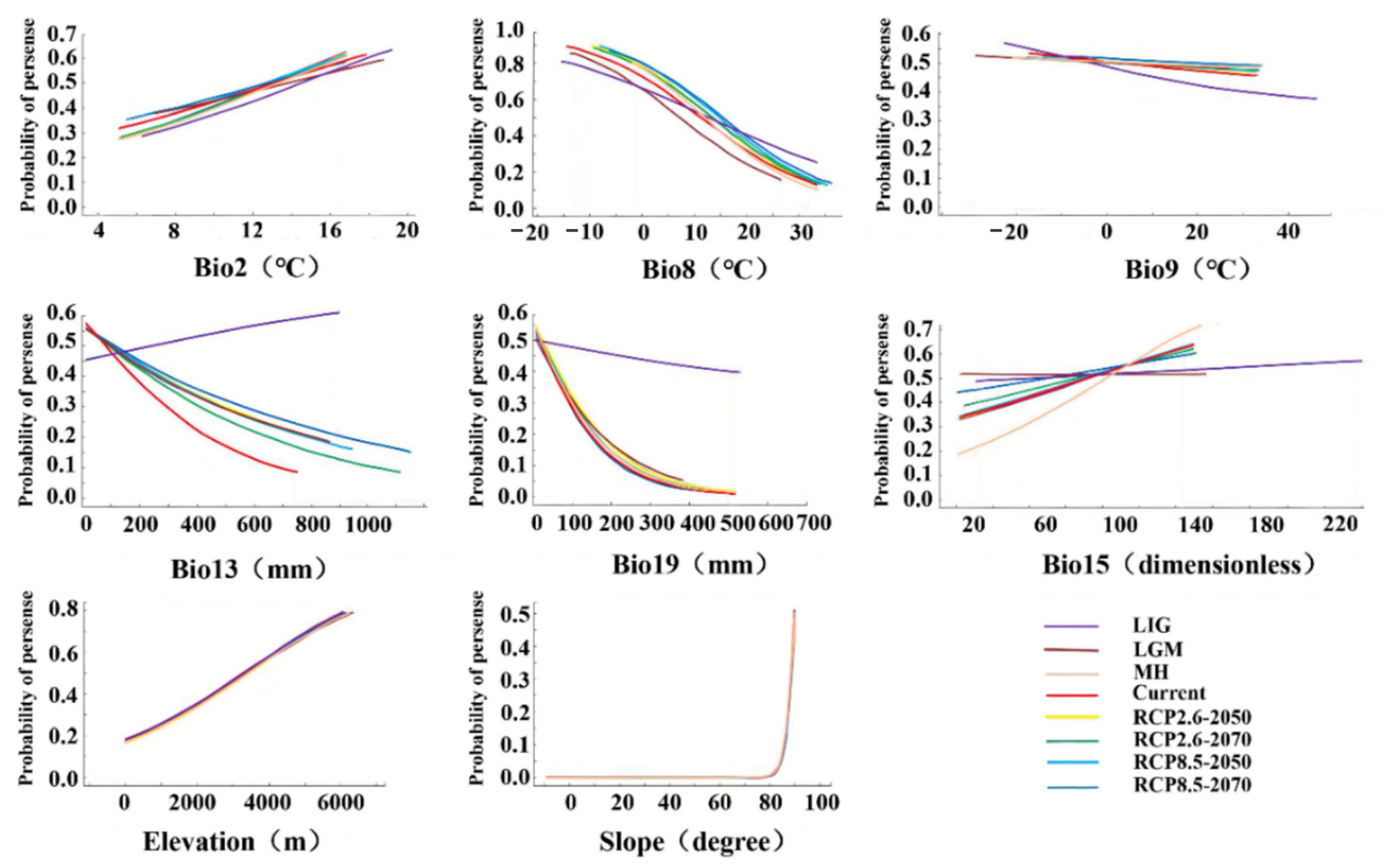
3.3. Relationship between Environmental Factors and the Distribution Pattern of D. validus
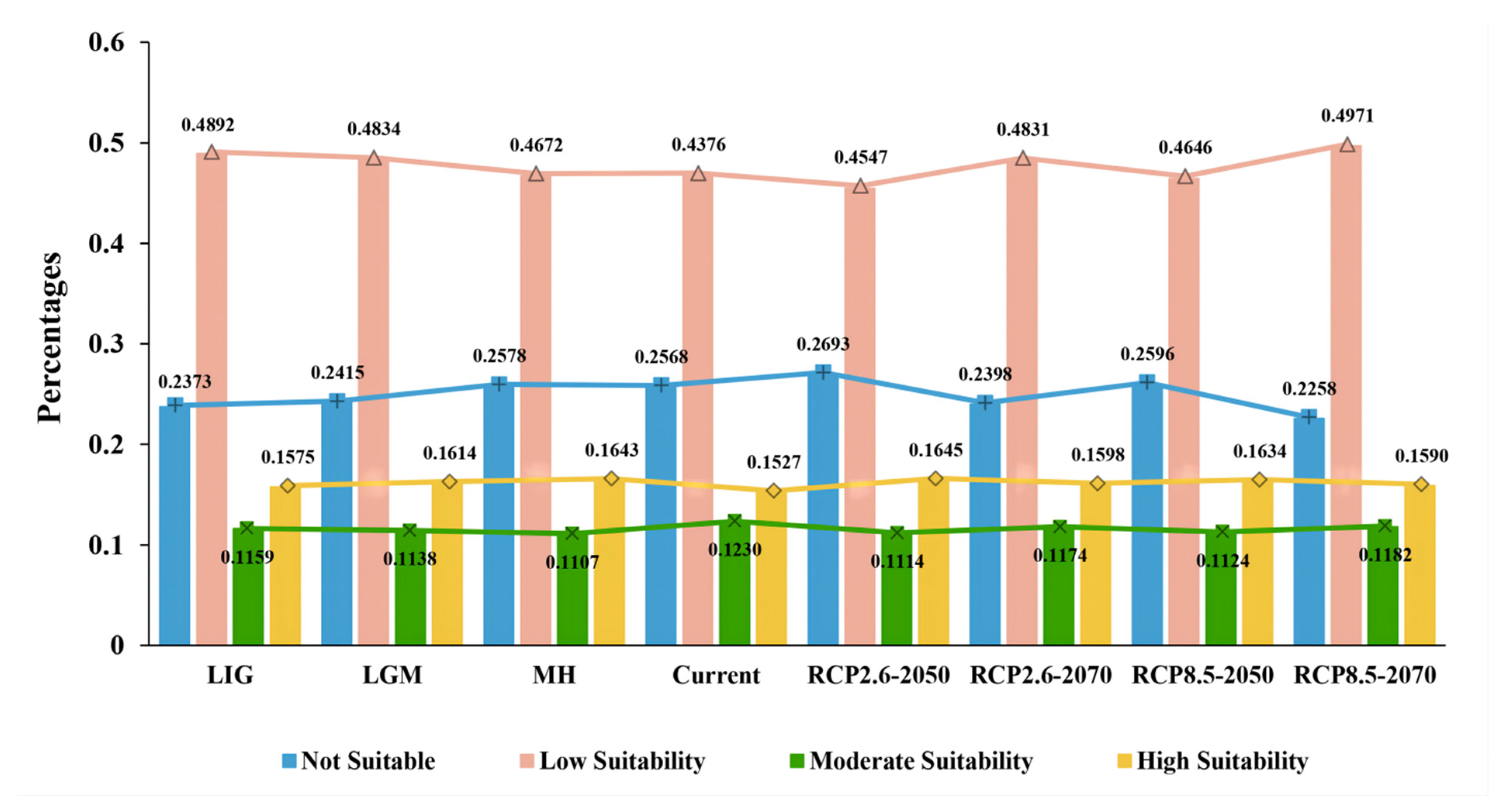
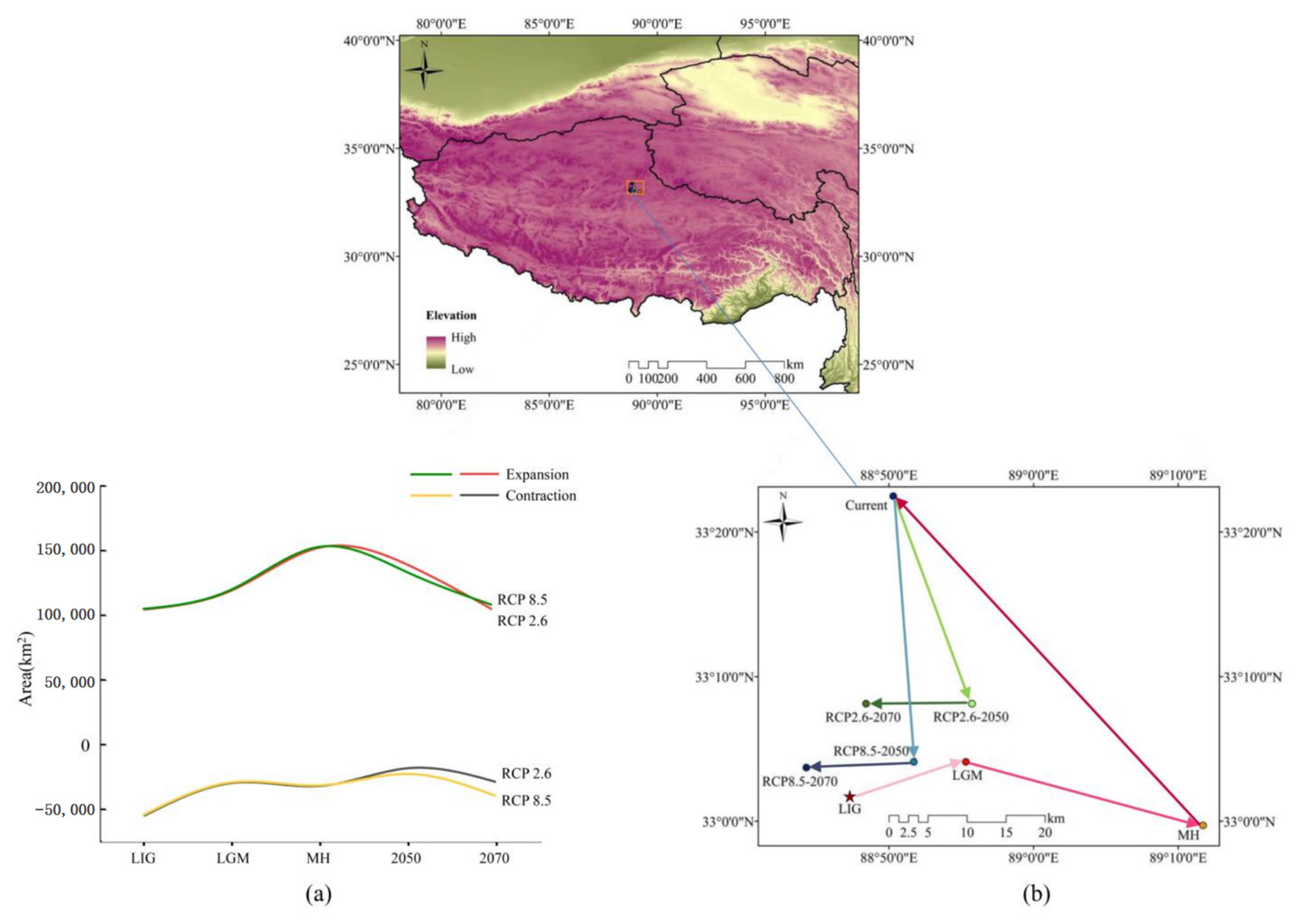
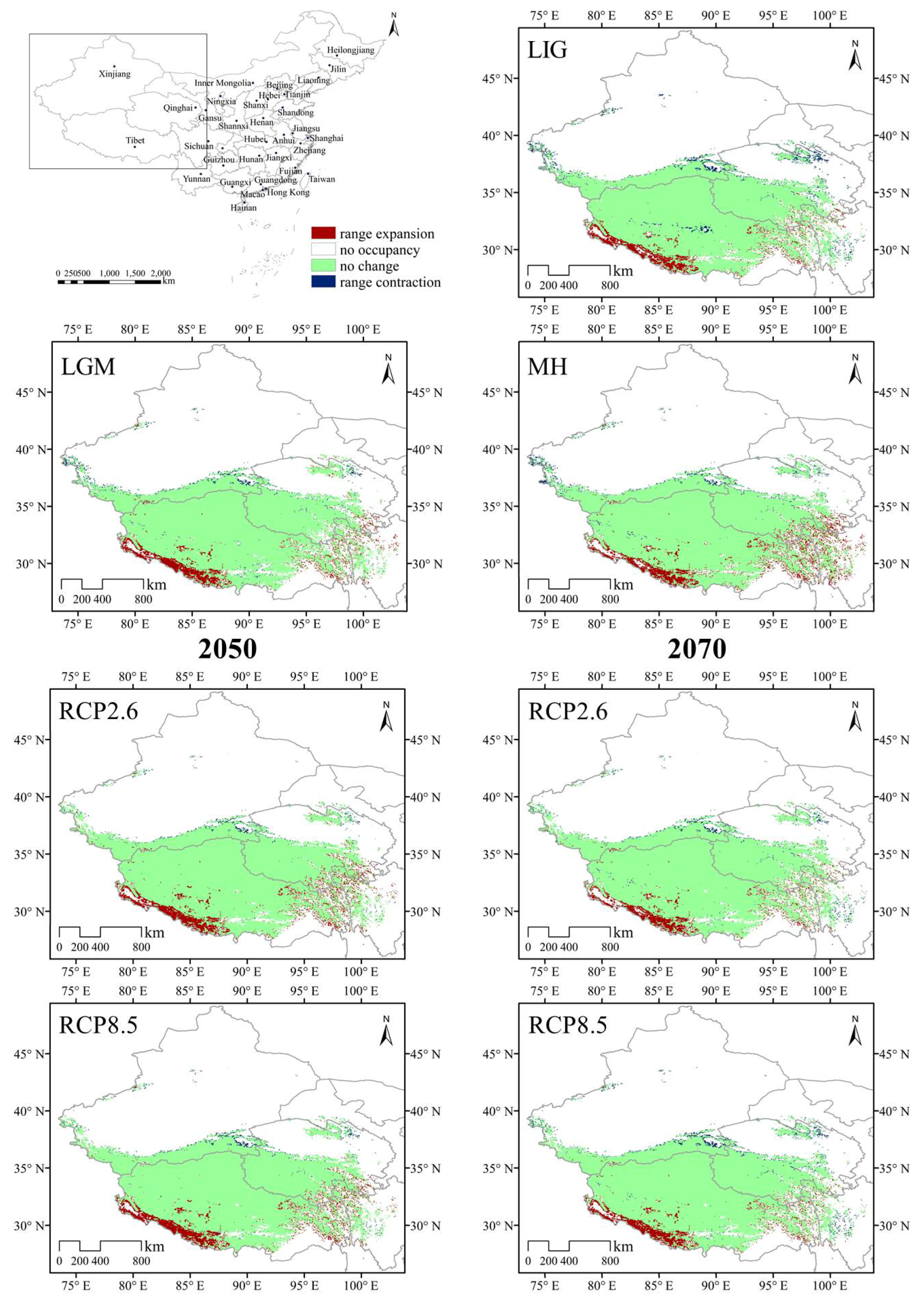
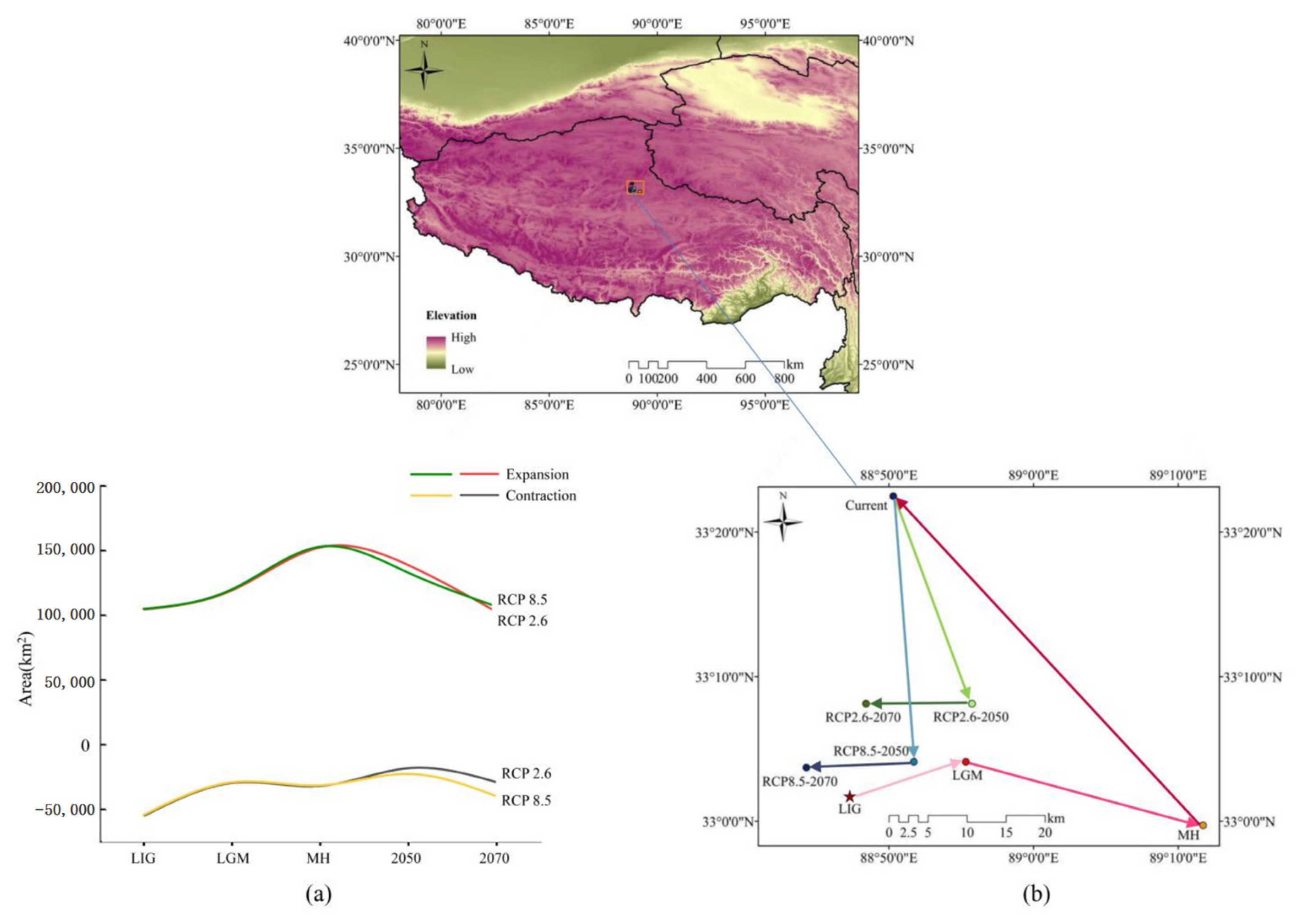
3.4. Species Distribution Shifts under Different Climatic Scenarios
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- He, X.; He, K.S.; Hyvönen, J. Will bryophytes survive in a warming world? Perspect. Plant Ecol. Evol. Syst. 2016, 19, 49–60. [Google Scholar] [CrossRef]
- Boch, S.; Muller, J.; Prati, D.; Fischer, M. Low-intensity management promotes bryophyte diversity in grasslands. Tuexenia 2018, 38, 311–328. [Google Scholar] [CrossRef]
- Bansal, S.; Nilsson, M.C.; Wardle, D.A. Response of photosynthetic carbon gain to ecosystem retrogression of vascular plants and mosses in the boreal forest. Oecologia 2012, 169, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Turetsky, M.R.; Bond-Lamberty, B.; Euskirchen, E.; Talbot, J.; Frolking, S.; McGuire, A.D.; Tuittila, E.S. The resilience and functional role of moss in boreal and arctic ecosystems. New Phytol. 2012, 196, 49–67. [Google Scholar] [CrossRef]
- Wang, S.Q.; Zhang, Z.H.; Wang, Z.H. Bryophyte communities as biomonitors of environmental factors in the Goujiang karst bauxite, southwestern China. Sci. Total Environ. 2015, 538, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, J.L.; Stark, L.R.; Chiquoine, L.P. Effects of Rate of Drying, Life History Phase, and Ecotype on the Ability of the Moss Bryum Argenteum to Survive Desiccation Events and the Influence on Conservation and Selection of Material for Restoration. Front. Ecol. Evol. 2019, 7, 388. [Google Scholar] [CrossRef]
- Ren, J.; Liu, F.; Luo, Y.; Zhu, J.; Luo, X.Q.; Liu, R. The Pioneering Role of Bryophytes in Ecological Restoration of Manganese Waste Residue Areas, Southwestern China. J. Chem. 2021, 2021, 9969253. [Google Scholar] [CrossRef]
- Di Nuzzo, L.; Vallese, C.; Benesperi, R.; Giordani, P.; Chiarucci, A.; Di Cecco, V.; Di Martino, L.; Di Musciano, M.; Gheza, G.; Lelli, C.; et al. Contrasting multitaxon responses to climate change in Mediterranean mountains. Sci. Rep. 2021, 11, 4438. [Google Scholar] [CrossRef]
- Frahm, J.P.; Newton, A.E. A new contribution to the moss flora of Dominican amber. Bryologist 2005, 108, 526–536. [Google Scholar] [CrossRef]
- Berdugo, M.B.; Quant, J.M.; Wason, J.W.; Dovciak, M. Latitudinal patterns and environmental drivers of moss layer cover in extratropical forests. Glob. Ecol. Biogeogr. 2018, 27, 1213–1224. [Google Scholar] [CrossRef]
- Kropik, M.; Zechmeister, H.G.; Moser, D.; Bernhardt, K.G.; Dullinger, S. Deadwood volumes matter in epixylic bryophyte conservation, but precipitation limits the establishment of substrate-specific communities. For. Ecol. Manag. 2021, 493, 119285. [Google Scholar] [CrossRef]
- Zander, R. Genera of the Pottiaceae: Mosses of Harsh Environments; Buffalo Society of Natural Sciences Press: New York, NY, USA, 1993; Volume 32. [Google Scholar]
- Song, S.S.; Liu, X.H.; Bai, X.L.; Jiang, Y.B.; Zhang, X.Z.; Yu, C.Q.; Shao, X.M. Impacts of Environmental Heterogeneity on Moss Diversity and Distribution of Didymodon (Pottiaceae) in Tibet, China. PLoS ONE 2015, 10, e0132346. [Google Scholar] [CrossRef] [PubMed]
- Kou, J.; Feng, C.; Jiang, Y.-B.; Shao, X.-M. Didymodon alpinus (Pottiaceae), a new species from Tibet, China. J. Bryol. 2017, 39, 308–312. [Google Scholar] [CrossRef]
- Kou, J.; Feng, C.; Shao, X.-M. Didymodon obtusus (Bryophyta, Pottiaceae), a new species from Tibet, China. Phytotaxa 2018, 372, 97–103. [Google Scholar] [CrossRef]
- Kou, J.; Wang, T.; Yu, F.; Sun, Y.; Feng, C.; Shao, X. The moss genus Didymodon as an indicator of climate change on the Tibetan Plateau. Ecol. Indic. 2020, 113, 106204. [Google Scholar] [CrossRef]
- Jia, S.-H.; Zhang, Z.-H. Diversity of the Bryophytes and Their Roles in Soil and Water Conservation in Karst City Rocky Desertification. Res. Soil Water Conserv. 2014, 21, 100–105. [Google Scholar] [CrossRef]
- Tian, G.Q.; Zhao, D.P. Species composition of bryophyte crusts and microhabitat formation under artificial vegetations in Huangfuchuan watershed, Inner Mongolia, China. Chin. J. Ecol. 2015, 34, 2448–2456. [Google Scholar]
- Kürschner, H.; Neef, R. A collection of bryophytes from the Tigray province, N Ethiopia and first evidence for a Syntrichion laevipilae Ochsner 1928 community in xerotropical Africa. Nova Hedwig. 2012, 95, 403–422. [Google Scholar] [CrossRef]
- Kürschner, H.; Shumilovskikh, L.; Djamali, M.; de Beaulieu, J.L. A late Holocene subfossil record of Sphagnum squarrosum Crome (Sphagnopsida, Bryophyta) from NW Iran. Nova Hedwig. 2015, 100, 373–381. [Google Scholar] [CrossRef]
- You, J.L.; Qin, X.P.; Ranjitkar, S.; Lougheed, S.C.; Wang, M.C.; Zhou, W.; Ouyang, D.X.; Zhou, Y.; Xu, J.C.; Zhang, W.J.; et al. Response to climate change of montane herbaceous plants in the genus Rhodiola predicted by ecological niche modelling. Sci. Rep. 2018, 8, 5879. [Google Scholar] [CrossRef]
- Yu, H.; Cooper, A.R.; Infante, D.M. Improving species distribution model predictive accuracy using species abundance: Application with boosted regression trees. Ecol. Model. 2020, 432, 109202. [Google Scholar] [CrossRef]
- Yi, Y.-j.; Cheng, X.; Yang, Z.-F.; Zhang, S.-H. Maxent modeling for predicting the potential distribution of endangered medicinal plant (H. riparia Lour) in Yunnan, China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Tarnian, F.; Kumar, S.; Azarnivand, H.; Chahouki, M.A.Z.; Mossivand, A.M. Assessing the effects of climate change on the distribution of Daphne mucronata in Iran. Environ. Monit. Assess. 2021, 193, 562. [Google Scholar] [CrossRef] [PubMed]
- Sérgio, C.; Figueira, R.; Draper, D.; Menezes, R.; Sousa, A.J. Modelling bryophyte distribution based on ecological information for extent of occurrence assessment. Biol. Conserv. 2007, 135, 341–351. [Google Scholar] [CrossRef]
- Kong, F.; Tang, L.; He, H.; Yang, F.X.; Tao, J.; Wang, W.C. Assessing the impact of climate change on the distribution of Osmanthus fragrans using Maxent. Environ. Sci. Pollut. Res. 2021, 28, 34655–34663. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liang, X.-Z.; Gao, W.; Stohlgren, T.J. Regional climate model downscaling may improve the prediction of alien plant species distributions. Front. Earth Sci. 2014, 8, 457–471. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crep. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Gonzalez, I.; Hebel, I.; Jaña, R. Distribución potencial de Sanionia spp. en dos momentos del Holoceno en península Fildes isla Rey Jorge Antártica. Anales del Instituto de la Patagonia 2019, 47, 7–17. [Google Scholar] [CrossRef]
- Nzei, J.M.; Ngarega, B.K.; Mwanzia, V.M.; Musili, P.M.; Wang, Q.-F.; Chen, J.-M. The past, current, and future distribution modeling of four water lilies (Nymphaea) in Africa indicates varying suitable habitats and distribution in climate change. Aquat. Bot. 2021, 173, 103416. [Google Scholar] [CrossRef]
- Yusup, S.; Sulayman, M.; Ilghar, W.; Zhang, Z.X. Prediction of potential distribution of Didymodon ( Bryophyta, Pottiaceae) in Xinjiang based on the MaxEnt model. Plant Sci. J. 2018, 36, 541–553. [Google Scholar]
- Ochyra, R.; Bednarek-Ochyra, H. The Correct Name for Didymodon Validus (Bryophyta, Pottiaceae) at Variety Rank. Pol. Bot. J. 2017, 62, 183–186. [Google Scholar] [CrossRef]
- Kou, J.; Feng, C.; Bai, X.L.; Chen, H. Morphology and taxonomy of leaf papillae and mammillae in Pottiaceae of China. J. Syst. Evol. 2014, 52, 521–532. [Google Scholar] [CrossRef]
- Zhang, G.-L.; Feng, C.; Kou, J.; Han, Y.; Zhang, Y.; Xiao, H.-X. Phylogeny and divergence time estimation of the genus Didymodon (Pottiaceae) based on nuclear and chloroplast markers. J. Syst. Evol. 2023, 61, 115–126. [Google Scholar] [CrossRef]
- Ren, D.M. Studies on Taxonomy and Flora of Pottiaceae in China. Doctoral Thesis, Inner Mongol University, Hohhot, China, 2012. [Google Scholar]
- Jia, Y.; He, S. Species Catalogue of China. Volume 1, Plants, Bryophytes; Science Press: Beijing, China, 2013. [Google Scholar]
- Yusup, S.; Sulayman, M.; Zhao, D.P. Didymodon validus Limpr., New to the moss flora of China. Acta Bot. Boreali Occident. Sin. 2017, 37, 1038–1041. [Google Scholar]
- Zhang, Z.Y.; Zhang, L.; Li, Y.H.; He, X.L. Study on the diversity of soil macrofauna in different habitats in Shilin karsts of Yunnan Province, China. For. Res. 2005, 18, 701–705. [Google Scholar]
- Yehya, M.; Aziz, R.; Sulayman, M. Taxonomy and distribution of Aloina Kindb. in Xinjiang, China. Plant Sci. 2020, 38, 448–457. [Google Scholar]
- Hijmans, R.; Cameron, S.; Parra, J.; Jones, P.; Jarvis, A. Very high resolution interpolated climate surfaces of global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- IPCC. Summer for Policymakers, Climate Change 2014: Impacts, Adaptation, and Vulnerability; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Hoveka, L.N.; Bezeng, B.S.; Yessoufou, K.; Boatwright, J.S.; Van der Bank, M. Effects of climate change on the future distributions of the top five freshwater invasive plants in South Africa. S. Afr. J. Bot. 2016, 102, 33–38. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Tao, J. Predicting the Potential Distribution of Paeonia veitchii (Paeoniaceae) in China by Incorporating Climate Change into a Maxent Model. Forests 2019, 10, 190. [Google Scholar] [CrossRef]
- Thuiller, W. Patterns and uncertainties of species’ range shifts under climate change. Glob. Change Biol. 2004, 10, 2020–2027. [Google Scholar] [CrossRef]
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Pepe, G.; Giusti, B.; Yacoub, M. An expanding role of biomarkers in acute aortic syndromes. Clin. Cardiol. 2006, 29, 432–433. [Google Scholar] [CrossRef]
- Pearce, J.; Ferrier, S. Evaluating the predictive performance of habitat models developed using logistic regression. Ecol. Model. 2000, 133, 225–245. [Google Scholar] [CrossRef]
- del Hoyo, L.V.; Martin Isabel, M.P.; Martinez Vega, F.J. Logistic regression models for human-caused wildfire risk estimation: Analysing the effect of the spatial accuracy in fire occurrence data. Eur. J. For. Res. 2011, 130, 983–996. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Jiménez, J.A. Taxonomic revision of the genus Didymodon Hedw. (Pottiaceae, Bryophyta) in Europe, North Africa and Southwest and Central Asia. J. Hattori Bot. Lab. 2006, 100, 211–292. [Google Scholar]
- Ma, B.; Sun, J. Predicting the distribution of Stipa purpurea across the Tibetan Plateau via the MaxEnt model. BMC Ecol. 2018, 18, 10. [Google Scholar] [CrossRef]
- Baye, T.M.; Abebe, T.; Wilke, R.A. Genotype-environment interactions and their translational implications. Pers. Med. 2011, 8, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Li, Y.; Cao, W.; He, X.Y.; Chen, W.; Xu, S. Prediction of Suitable Habitat for Lycophytes and Ferns in Northeast China: A Case Study on Athyrium brevifrons. Chin. Geogr. Sci. 2019, 29, 1011–1023. [Google Scholar] [CrossRef]
- Wan, J.Z.; Wang, C.J.; Han, S.J.; Yu, J.H. Planning the priority protected areas of endangered orchid species in northeastern China. Biodivers. Conserv. 2014, 23, 1395–1409. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Thuiller, W.; Miaud, C. Prediction and validation of the potential global distribution of a problematic alien invasive species—The American bullfrog. Divers. Distrib. 2007, 13, 476–485. [Google Scholar] [CrossRef]
- Kumar, P. Assessment of impact of climate change on Rhododendrons in Sikkim Himalayas using Maxent modelling: Limitations and challenges. Biodivers. Conserv. 2012, 21, 1251–1266. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M.; et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Bátori, Z.; Vojtkó, A.; Farkas, T.; Szabó, A.; Havadtői, K.; Vojtkó, A.E.; Tölgyesi, C.; Cseh, V.; Erdős, L.; Maák, I.E.; et al. Large- and small-scale environmental factors drive distributions of cool-adapted plants in karstic microrefugia. Ann. Bot. 2017, 119, 301–309. [Google Scholar] [CrossRef]
- Scherrer, D.; Körner, C. Topographically controlled thermal-habitat differentiation buffers alpine plant diversity against climate warming. J. Biogeogr. 2011, 38, 406–416. [Google Scholar] [CrossRef]
- Ronikier, M.; Schneeweiss, G.M.; Schönswetter, P. The extreme disjunction between Beringia and Europe in Ranunculus glacialis s. l. (Ranunculaceae) does not coincide with the deepest genetic split—A story of the importance of temperate mountain ranges in arctic-alpine phylogeography. Mol. Ecol. 2021, 21, 5561–5578. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, D.; Barik, S.K.; Upadhaya, K. Habitat distribution modelling for reintroduction of Ilex khasiana Purk., a critically endangered tree species of northeastern India. Ecol. Eng. 2012, 40, 37–43. [Google Scholar] [CrossRef]
- Spitale, D. The interaction between elevational gradient and substratum reveals how bryophytes respond to the climate. J. Veg. Sci. 2016, 27, 844–853. [Google Scholar] [CrossRef]
- Maciel-Silva, A.S.; Válio, I.F.M. Reproductive phenology of bryophytes in tropical rain forests: The sexes never sleep. Bryologist 2011, 114, 708–719. [Google Scholar] [CrossRef]
- Cleavitt, N. Comparative ecology of a lowland and a subalpine species of Mnium in the northern Rocky Mountains. Plant Ecol. 2004, 174, 205–216. [Google Scholar] [CrossRef]
- Alatalo, J.M.; Jägerbrand, A.K.; Molau, U. Climate change and climatic events: Community-, functional- and species-level responses of bryophytes and lichens to constant, stepwise, and pulse experimental warming in an alpine tundra. Alp. Bot. 2014, 124, 81–91. [Google Scholar] [CrossRef]
- Zheng, B.X.; Xu, Q.Q.; Shen, Y.P. The relationship between climate change and Quaternary glacial cycles on the Qinghai-Tibetan Plateau: Review and speculation. Quat. Int. 2002, 97–98, 93–101. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef]
- Zhao, H.X.; Zhang, H.; Xu, C.G. Study on Taiwania cryptomerioides under climate change: MaxEnt modeling for predicting the potential geographical distribution. Glob. Ecol. Conserv. 2020, 24, e01313. [Google Scholar] [CrossRef]
- Zheng, H.-Y.; Guo, X.-L.; Price, M.; He, X.-J.; Zhou, S.-D. Effects of Mountain Uplift and Climatic Oscillations on Phylogeography and Species Divergence of Chamaesium (Apiaceae). Front. Plant Sci. 2021, 12, 673200. [Google Scholar] [CrossRef]
- Lister, A.M. The impact of Quaternary Ice Ages on mammalian evolution. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 221–241. [Google Scholar] [CrossRef] [PubMed]
- Botts, E.A.; Erasmus, B.F.N.; Alexander, G.J. Small range size and narrow niche breadth predict range contractions in South African frogs. Glob. Ecol. Biogeogr. 2013, 22, 567–576. [Google Scholar] [CrossRef]
- Dubos, N.; Frederique, M.; Grinand, C.; Nourtier, M.; Deso, G.; Jean-Michel, P.; Razafimanahaka, J.; Andriantsimanarilafy, R.; Rakotondrasoa, E.; Razafindraibe, P.; et al. Are narrow-ranging species doomed to extinction? Projected dramatic decline in future climate suitability of two highly threatened species. Perspect. Ecol. Conserv. 2021, 20, 18–28. [Google Scholar] [CrossRef]
- Chevin, L.-M.; Lande, R.; Mace, G.M. Adaptation, Plasticity, and Extinction in a Changing Environment: Towards a Predictive Theory. PLoS Biol. 2010, 8, e1000357. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Sgrò, C.M. Climate change and evolutionary adaptation. Nature 2011, 470, 479–485. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, T.; Pan, C.; Bian, T.; Wang, Q.; Kou, J.; Zhou, B. Response of a Sylvan Moss Species (Didymodon validus Limpr.) with a Narrow Distribution Range to Climate Change. Forests 2023, 14, 2227. https://doi.org/10.3390/f14112227
Wu T, Pan C, Bian T, Wang Q, Kou J, Zhou B. Response of a Sylvan Moss Species (Didymodon validus Limpr.) with a Narrow Distribution Range to Climate Change. Forests. 2023; 14(11):2227. https://doi.org/10.3390/f14112227
Chicago/Turabian StyleWu, Tingting, Chuntong Pan, Tao Bian, Qiaoxin Wang, Jin Kou, and Bangwei Zhou. 2023. "Response of a Sylvan Moss Species (Didymodon validus Limpr.) with a Narrow Distribution Range to Climate Change" Forests 14, no. 11: 2227. https://doi.org/10.3390/f14112227
APA StyleWu, T., Pan, C., Bian, T., Wang, Q., Kou, J., & Zhou, B. (2023). Response of a Sylvan Moss Species (Didymodon validus Limpr.) with a Narrow Distribution Range to Climate Change. Forests, 14(11), 2227. https://doi.org/10.3390/f14112227