
Quercus petraea (Matt.) Liebl. from the Thayatal National Park in Austria: Selection of Potentially Drought-Tolerant Phenotypes
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- Are the Thayatal National Park’s sessile oaks of autochthonous origin?
- Is there evidence of genetic structure among sessile oaks across the Thayatal National Park? If so, how is it reflected in patterns of genetic diversity?
- Do the sessile oaks in the Thayatal National Park exhibit variations in drought tolerance differences, and if so, how many potentially drought-tolerant phenotypes can be identified among the studied trees?
2. Material and Methods
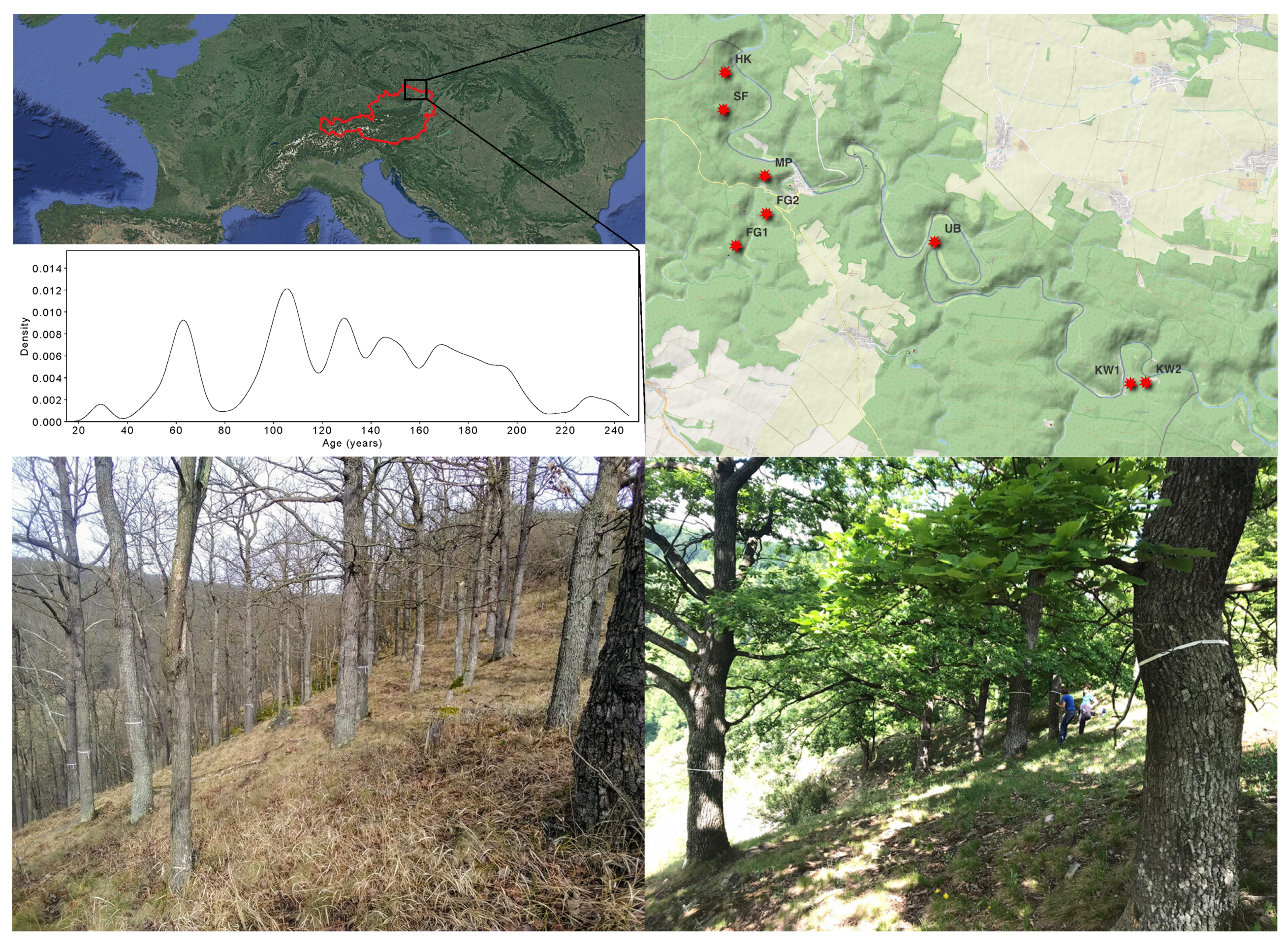
2.1. Study Area and Tree Selection
2.2. Tree Characterization, Material Collection and Processing
2.3. Genotyping for Analysis of Autochthony, Genetic Diversity and Genetic Structure
2.4. Wood Core Analyses: Age, Drought Tolerance, and Water Use Efficiency
3. Results
3.1. Autochthony
3.2. Genetic Diversity and Genetic Structure
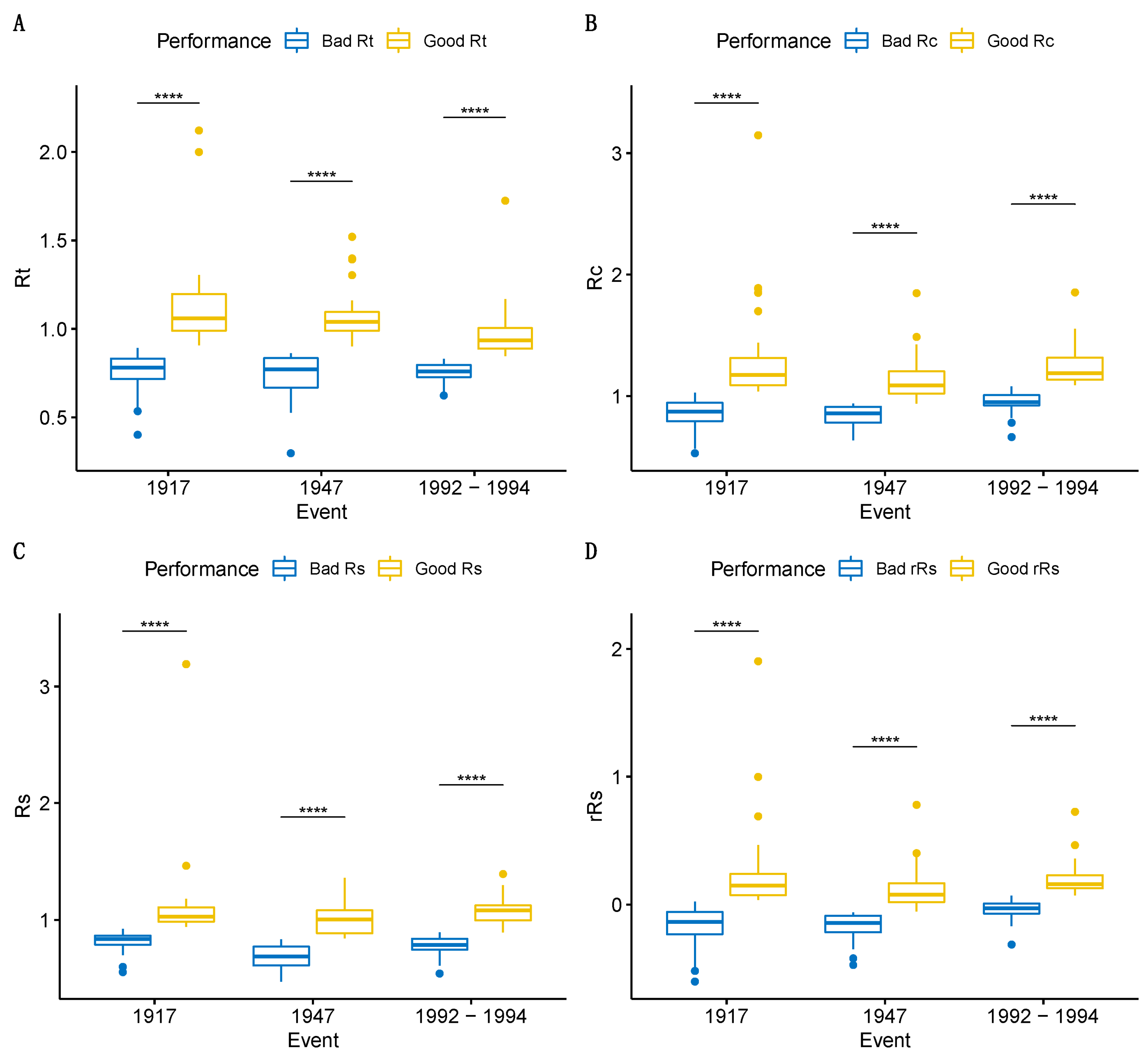
3.3. Drought Tolerance
4. Discussion
4.1. Autochthony
4.2. Patterns of Genetic Structure and Diversity Linked with Age Structure
4.3. Response to Extreme Drought Years and Selection of the Best Drought-Adapted Phenotypes
5. Conclusions and Outlook Related to Breeding and Conservation
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chakraborty, D.; Móricz, N.; Rasztovits, E.; Dobor, L.; Schueler, S. Provisioning Forest and Conservation Science with High-Resolution Maps of Potential Distribution of Major European Tree Species under Climate Change. Ann. For. Sci. 2021, 78, 26. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How Much Does Climate Change Threaten European Forest Tree Species Distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef] [PubMed]
- Illés, G.; Móricz, N. Climate Envelope Analyses Suggests Significant Rearrangements in the Distribution Ranges of Central European Tree Species. Ann. For. Sci. 2022, 79, 35. [Google Scholar] [CrossRef]
- Bader, M.K.-F.; Scherrer, D.; Zweifel, R.; Körner, C. Less Pronounced Drought Responses in Ring-Porous than in Diffuse-Porous Temperate Tree Species. Agric. For. Meteorol. 2022, 327, 109184. [Google Scholar] [CrossRef]
- Scherrer, D.; Bader, M.K.-F.; Körner, C. Drought-Sensitivity Ranking of Deciduous Tree Species Based on Thermal Imaging of Forest Canopies. Agric. For. Meteorol. 2011, 151, 1632–1640. [Google Scholar] [CrossRef]
- Vanhellemont, M.; Sousa-Silva, R.; Maes, S.L.; Van Den Bulcke, J.; Hertzog, L.; De Groote, S.R.E.; Van Acker, J.; Bonte, D.; Martel, A.; Lens, L.; et al. Distinct Growth Responses to Drought for Oak and Beech in Temperate Mixed Forests. Sci. Total Environ. 2019, 650, 3017–3026. [Google Scholar] [CrossRef] [PubMed]
- Bose, A.K.; Scherrer, D.; Camarero, J.J.; Ziche, D.; Babst, F.; Bigler, C.; Bolte, A.; Dorado-Liñán, I.; Etzold, S.; Fonti, P.; et al. Climate Sensitivity and Drought Seasonality Determine Post-Drought Growth Recovery of Quercus Petraea and Quercus Robur in Europe. Sci. Total Environ. 2021, 784, 147222. [Google Scholar] [CrossRef] [PubMed]
- Zweifel, R.; Rigling, A.; Dobbertin, M. Species-Specific Stomatal Response of Trees to Drought—A Link to Vegetation Dynamics? J. Veg. Sci. 2009, 20, 442–454. [Google Scholar] [CrossRef]
- Günthardt-Goerg, M.S.; Kuster, T.M.; Arend, M.; Vollenweider, P. Foliage Response of Young Central European Oaks to Air Warming, Drought and Soil Type: Oak Foliage Response to Air Warming, Drought and Soil pH. Plant Biol. 2013, 15, 185–197. [Google Scholar] [CrossRef]
- Kuster, T.M.; Arend, M.; Günthardt-Goerg, M.S.; Schulin, R. Root Growth of Different Oak Provenances in Two Soils under Drought Stress and Air Warming Conditions. Plant Soil 2013, 369, 61–71. [Google Scholar] [CrossRef]
- Vollenweider, P.; Menard, T.; Arend, M.; Kuster, T.M.; Günthardt-Goerg, M.S. Structural Changes Associated with Drought Stress Symptoms in Foliage of Central European Oaks. Trees 2016, 30, 883–900. [Google Scholar] [CrossRef]
- POWO Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 28 August 2023).
- Saleh, D.; Chen, J.; Leplé, J.-C.; Leroy, T.; Truffaut, L.; Dencausse, B.; Lalanne, C.; Labadie, K.; Lesur, I.; Bert, D.; et al. Genome-Wide Evolutionary Response of European Oaks during the Anthropocene. Evol. Lett. 2022, 6, 4–20. [Google Scholar] [CrossRef] [PubMed]
- European Commission a European Green Deal. Available online: https://commission.europa.eu/strategy-and-policy/priorities-2019-2024/european-green-deal_en (accessed on 28 August 2023).
- Jalonen, R.; Valette, M.; Boshier, D.; Duminil, J.; Thomas, E. Forest and Landscape Restoration Severely Constrained by a Lack of Attention to the Quantity and Quality of Tree Seed: Insights from a Global Survey. Conserv. Lett. 2018, 11, e12424. [Google Scholar] [CrossRef]
- Gaisberger, H.; Fremout, T.; Kettle, C.J.; Vinceti, B.; Kemalasari, D.; Kanchanarak, T.; Thomas, E.; Serra-Diaz, J.M.; Svenning, J.-C.; Slik, F.; et al. Tropical and Subtropical Asia’s Valued Tree Species under Threat. Conserv. Biol. 2022, 36, e13873. [Google Scholar] [CrossRef] [PubMed]
- Pörtner, H.-O.; Roberts, D.C.; Tignor, M.M.B.; Poloczanska, E.S.; Mintenbeck, K.; Alegría, A.; Craig, M.; Langsdorf, S.; Löschke, S.; Möller, V.; et al. (Eds.) IPCC, 2022: Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022; p. 3056. [Google Scholar]
- Steckel, M.; Del Río, M.; Heym, M.; Aldea, J.; Bielak, K.; Brazaitis, G.; Černý, J.; Coll, L.; Collet, C.; Ehbrecht, M.; et al. Species Mixing Reduces Drought Susceptibility of Scots Pine (Pinus sylvestris L.) and Oak (Quercus robur L., Quercus petraea (Matt.) Liebl.)—Site Water Supply and Fertility Modify the Mixing Effect. For. Ecol. Manag. 2020, 461, 117908. [Google Scholar] [CrossRef]
- Bolte, A.; Ammer, C.; Löf, M.; Madsen, P.; Nabuurs, G.-J.; Schall, P.; Spathelf, P.; Rock, J. Adaptive Forest Management in Central Europe: Climate Change Impacts, Strategies and Integrative Concept. Scand. J. For. Res. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Twardek, W.M.; Taylor, J.J.; Rytwinski, T.; Aitken, S.N.; MacDonald, A.L.; Van Bogaert, R.; Cooke, S.J. The Application of Assisted Migration as a Climate Change Adaptation Tactic: An Evidence Map and Synthesis. Biol. Conserv. 2023, 280, 109932. [Google Scholar] [CrossRef]
- Sniezko, R.A.; Dana Nelson, C. Chapter 10 - Resistance Breeding against Tree Pathogens. In Forest Microbiology; Asiegbu, F.O., Kovalchuk, A., Eds.; Academic Press: Cambridge, MA, USA, 2022; Volume 2, pp. 159–175. [Google Scholar]
- Polle, A.; Chen, S.L.; Eckert, C.; Harfouche, A. Engineering Drought Resistance in Forest Trees. Front. Plant Sci. 2019, 9, 1875. [Google Scholar] [CrossRef]
- Baldi, P.; La Porta, N. Toward the Genetic Improvement of Drought Tolerance in Conifers: An Integrated Approach. Forests 2022, 13, 2016. [Google Scholar] [CrossRef]
- Schoppa, F.N.; Gregorius, H.-R. Is Autochthony an Operational Concept? In Genetic Response of Forest Systems to Changing Environmental Conditions; Müller-Starck, G., Schubert, R., Eds.; Forestry Sciences; Springer: Dordrecht, The Netherlands, 2001; pp. 173–185. ISBN 978-94-015-9839-2. [Google Scholar]
- Petit, R.J.; Csaikl, U.M.; Bordacs, S.; Burg, K.; Coart, E.; Cottrell, J.; van Dam, B.; Deans, J.D.; Dumolin-Lapegue, S.; Fineschi, S.; et al. Chloroplast DNA Variation in European White Oaks Phylogeography and Patterns of Diversity Based on Data from over 2600 Populations. For. Ecol. Manag. 2002, 156, 5–26. [Google Scholar] [CrossRef]
- Petit, R.J.; Brewer, S.; Bordács, S.; Burg, K.; Cheddadi, R.; Coart, E.; Cottrell, J.; Csaikl, U.M.; van Dam, B.; Deans, J.D.; et al. Identification of Refugia and Post-Glacial Colonisation Routes of European White Oaks Based on Chloroplast DNA and Fossil Pollen Evidence. For. Ecol. Manag. 2002, 156, 49–74. [Google Scholar] [CrossRef]
- Degen, B.; Yanbaev, Y.; Mader, M.; Ianbaev, R.; Bakhtina, S.; Schroeder, H.; Blanc-Jolivet, C. Impact of Gene Flow and Introgression on the Range Wide Genetic Structure of Quercus robur (L.) in Europe. Forests 2021, 12, 1425. [Google Scholar] [CrossRef]
- Fortini, P.; Di Pietro, R.; Proietti, E.; Cardoni, S.; Quaranta, L.; Simeone, M.C. Dissecting the Continuum and Unravelling the Phylogeographic Knot of Plastid DNA in European White Oaks (Quercus Sect. Quercus): Ancient Signatures and Multiple Diversity Reservoirs. Eur. J. For. Res. 2023, 1–21. [Google Scholar] [CrossRef]
- Huntley, B.; Birks, H.J.B. An Atlas of Past and Present Pollen Maps for Europe: 0–13,000 Years Ago: The Overlay Maps; Cambridge University Press: Cambridge, UK, 1983. [Google Scholar]
- Brewer, S.; Cheddadi, R.; De Beaulieu, J.L.; Reille, M. The Spread of Deciduous Quercus throughout Europe since the Last Glacial Period. For. Ecol. Manag. 2002, 156, 27–48. [Google Scholar] [CrossRef]
- Svenning, J.; Normand, S.; Kageyama, M. Glacial Refugia of Temperate Trees in Europe: Insights from Species Distribution Modelling. J. Ecol. 2008, 96, 1117–1127. [Google Scholar] [CrossRef]
- Stewart, J.R.; Lister, A.M. Cryptic Northern Refugia and the Origins of the Modern Biota. Trends Ecol. Evol. 2001, 16, 608–613. [Google Scholar] [CrossRef]
- Neophytou, C.; Michiels, H.-G. Upper Rhine Valley: A Migration Crossroads of Middle European Oaks. For. Ecol. Manag. 2013, 304, 89–98. [Google Scholar] [CrossRef]
- Semerikova, S.A.; Isakov, I.Y.; Semerikov, V.L. Chloroplast DNA Variability and Phylogeography of Pedunculate Oak Quercus robur L. in the Eastern Part of the Range. Genetika 2021, 57, 56–71. [Google Scholar] [CrossRef]
- Bordács, S.; Popescu, F.; Slade, D.; Csaikl, U.M.; Lesur, I.; Borovics, A.; Kézdy, P.; König, A.O.; Gömöry, D.; Brewer, S.; et al. Chloroplast DNA Variation of White Oaks in Northern Balkans and in the Carpathian Basin. For. Ecol. Manag. 2002, 156, 197–209. [Google Scholar] [CrossRef]
- Csaikl, U.M.; Glaz, I.; Baliuckas, V.; Petit, R.J.; Jensen, J.S. Chloroplast DNA Variation of White Oak in the Baltic Countries and Poland. For. Ecol. Manag. 2002, 156, 211–222. [Google Scholar] [CrossRef]
- Csaikl, U.M.; Burg, K.; Fineschi, S.; König, A.O.; Mátyás, G.; Petit, R.J. Chloroplast DNA Variation of White Oaks in the Alpine Region. For. Ecol. Manag. 2002, 156, 131–145. [Google Scholar] [CrossRef]
- König, A.O.; Ziegenhagen, B.; Van Dam, B.C.; Csaikl, U.M.; Coart, E.; Degen, B.; Burg, K.; De Vries, S.M.G.; Petit, R.J. Chloroplast DNA Variation of Oaks in Western Central Europe and Genetic Consequences of Human Influences. For. Ecol. Manag. 2002, 156, 147–166. [Google Scholar] [CrossRef]
- Tutková-van Loo, M.; Burg, K. Chloroplast Haplotype Diversity of White Oak Species in Slovakia and the Czech Republic: Results from PCR-RFLP Analysis and Phylogeographic Interpretations. For. Genet. 2004, 10, 125–137. [Google Scholar]
- Rodriguez-Zaccaro, F.D.; Groover, A. Wood and Water: How Trees Modify Wood Development to Cope with Drought. Plants People Planet 2019, 1, 346–355. [Google Scholar] [CrossRef]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of Tree Resilience: Effects of Successive Low-Growth Episodes in Old Ponderosa Pine Forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Aldea, J.; Ruiz-Peinado, R.; Del Río, M.; Pretzsch, H.; Heym, M.; Brazaitis, G.; Jansons, A.; Metslaid, M.; Barbeito, I.; Bielak, K.; et al. Timing and Duration of Drought Modulate Tree Growth Response in Pure and Mixed Stands of Scots Pine and Norway Spruce. J. Ecol. 2022, 110, 2673–2683. [Google Scholar] [CrossRef]
- Brendel, O. The Relationship between Plant Growth and Water Consumption: A History from the Classical Four Elements to Modern Stable Isotopes. Ann. For. Sci. 2021, 78, 47. [Google Scholar] [CrossRef]
- Le Provost, G.; Brachi, B.; Lesur, I.; Lalanne, C.; Labadie, K.; Aury, J.-M.; Da Silva, C.; Postolache, D.; Leroy, T.; Plomion, C. Gene Expression and Genetic Divergence in Oak Species Highlight Adaptive Genes to Soil Water Constraints. Plant Physiol. 2022, 190, 2466–2483. [Google Scholar] [CrossRef]
- Wurth-Waitzbauer, C.; Übl, C. 10 Jahre wissenschaftliche Forschung im Nationalpark Thayatal. Wiss. Mitteilungen Niederösterreichisches Landesmus 2010, 21, 19–34. [Google Scholar]
- Roetzel, R. Geologie im Fluss. Erläuterungen zur Geologischen Karte der Nationalparks Thayatal und Podyjí; Geologische Bundesanstalt: Wien, Österreich, 2005. [Google Scholar]
- Roetzel, R.; Fuchs, G.; Batik, P.; Cytroky, P.; Havliček, P. Geologische Karte Der Nationalparks Thayatal Und Podyji; Geologische Bundesanstalt: Wien, Österreich, 2004. [Google Scholar]
- Wrbka, T.; Zmelik, K. Biodiversitätsforschung im Nationalpark Thayatal—Teilbereich Waldvegetation. Endbericht; Universität Wien, Fakultät für Lebenswissenschaften, Department für Naturschutzbiologie, Vegetations- und Landschaftsökologie: Wien, Österreich, 2006; p. 132. [Google Scholar]
- Baillie, M.G.L. A Slice Through Time: Dendrochronology and Precision Dating; Routledge: London, UK; New York, NY, USA, 1995; ISBN 978-0-7134-7654-5. [Google Scholar]
- Stokes, M.A. An Introduction to Tree-Ring Dating; University of Arizona Press: Tucson, AZ, USA, 1996; ISBN 978-0-8165-1680-3. [Google Scholar]
- Deguilloux, M.-F.; Dumolin-Lapègue, S.; Gielly, L.; Grivet, D.; Petit, R.J. A Set of Primers for the Amplification of Chloroplast Microsatellites in Quercus: PRIMER NOTE. Mol. Ecol. Notes 2003, 3, 24–27. [Google Scholar] [CrossRef]
- Weising, K.; Gardner, R.C. A Set of Conserved PCR Primers for the Analysis of Simple Sequence Repeat Polymorphisms in Chloroplast Genomes of Dicotyledonous Angiosperms. Genome 1999, 42, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Steinkellner, H.; Fluch, S.; Turetschek, E.; Lexer, C.; Streiff, R.; Kremer, A.; Burg, K.; Glossl, J. Identification and Characterization of (GA/CT) n- Microsatellite Loci from Quercus Petraea. Plant Mol. Biol. 1997, 33, 1093–1096. [Google Scholar] [CrossRef] [PubMed]
- Kampfer, S.; Lexer, C.; Glössl, J.; Steinkellner, H. Characterization of (GA)n Microsatellite Loci from Quercus Robur. Hereditas 1998, 129, 183–186. [Google Scholar] [CrossRef]
- Meirmans, P.G. Genodive Version 3.0: Easy-to-Use Software for the Analysis of Genetic Data of Diploids and Polyploids. Mol. Ecol. Resour. 2020, 20, 1126–1131. [Google Scholar] [CrossRef] [PubMed]
- Szpiech, Z.A.; Jakobsson, M.; Rosenberg, N.A. ADZE: A Rarefaction Approach for Counting Alleles Private to Combinations of Populations. Bioinformatics 2008, 24, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Jost, L. GST and Its Relatives Do Not Measure Differentiation. Mol. Ecol. 2008, 17, 4015–4026. [Google Scholar] [CrossRef] [PubMed]
- Keenan, K.; McGinnity, P.; Cross, T.F.; Crozier, W.W.; Prodöhl, P.A. diveRsity: An R Package for the Estimation and Exploration of Population Genetics Parameters and Their Associated Errors. Methods Ecol. Evol. 2013, 4, 782–788. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of Population Structure Using Multilocus Genotype Data: Linked Loci and Correlated Allele Frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of Population Structure Using Multilocus Genotype Data: Dominant Markers and Null Alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef]
- Earl, D.A.; vonHoldt, B.M. Structure Harvester: A Website and Program for Visualizing STRUCTURE Output and Implementing the Evanno Method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A Cluster Matching and Permutation Program for Dealing with Label Switching and Multimodality in Analysis of Population Structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, N.A. Distruct: A Program for the Graphical Display of Population Structure: Program Note. Mol. Ecol. Notes 2003, 4, 137–138. [Google Scholar] [CrossRef]
- Huang, K.; Ritland, K.; Guo, S.; Shattuck, M.; Li, B. A Pairwise Relatedness Estimator for Polyploids. Mol. Ecol. Resour. 2014, 14, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. Gplots: Various R Programming Tools for Plotting Data; R Package Version 3.1.3. 2022. Available online: https://cran.r-project.org/package=gplots (accessed on 8 September 2023).
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Lynch, M.; Walsh, B. Genetics and Analysis of Quantitative Traits; Sinauer Associates, Inc.: Sunderland, MA, USA, 1998; ISBN 978-0-87893-481-2. [Google Scholar]
- Nakagawa, S.; Schielzeth, H. Repeatability for Gaussian and Non-Gaussian Data: A Practical Guide for Biologists. Biol. Rev. 2010, 85, 935–956. [Google Scholar] [CrossRef] [PubMed]
- Stoffel, M.A.; Nakagawa, S.; Schielzeth, H. rptR: Repeatability Estimation and Variance Decomposition by Generalized Linear Mixed-effects Models. Methods Ecol. Evol. 2017, 8, 1639–1644. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Ubierna, N. Carbon Isotope Effects in Relation to CO2 Assimilation by Tree Canopies. In Stable Isotopes in Tree Rings: Inferring Physiological, Climatic and Environmental Responses; Siegwolf, R.T.W., Brooks, J.R., Roden, J., Saurer, M., Eds.; Tree Physiology; Springer International Publishing: Cham, Switzerland, 2022; pp. 291–310. ISBN 978-3-030-92698-4. [Google Scholar]
- van Loo, M.; Ufimov, R.; Grabner, M.; Übl, C.; Watzinger, A.; Irauschek, F.; Konrad, H.; Píšová, S.; Trujillo-Moya, C. Quercus petraea (Matt.) Liebl. from the Thayatal National Park in Austria: Selection of Potentially Drought Tolerant Phenotypes [Dataset]; Dryad: Leicester, UK, 2023. [Google Scholar] [CrossRef]
- Trujillo-Moya, C.; George, J.-P.; Fluch, S.; Geburek, T.; Grabner, M.; Karanitsch-Ackerl, S.; Konrad, H.; Mayer, K.; Sehr, E.M.; Wischnitzki, E.; et al. Drought Sensitivity of Norway Spruce at the Species’ Warmest Fringe: Quantitative and Molecular Analysis Reveals High Genetic Variation among and Within Provenances. G3 Genes Genomes Genet. 2018, 8, 1225–1245. [Google Scholar] [CrossRef]
- Krippel, E. Postglaciálny Vývoj Vegetácie Slovenska, 1st ed.; Veda Publishing House: Bratislava, Slovakia, 1986; p. 307. [Google Scholar]
- Küster, H. Auswirkungen von Klimaschwankungen und menschlicher Landschaftsnutzung auf die Arealverschiebung von Pflanzen und die Ausbildung mitteleuropäischer Wälder. Forstwiss. Cent. Ver. Mit Tharandter Forstl. Jahrb. 1996, 115, 301–320. [Google Scholar] [CrossRef]
- Küster, H. Geschichte der Landschaft in Mitteleuropa: Von der Eiszeit bis zur Gegenwart, 1st ed.; C.H.Beck: München, Germany, 1999; ISBN 978-3-406-45357-1. [Google Scholar]
- Lazišťan, E. Slovensko; Neografia: Martin, Slovakia, 2001; ISBN 80-88892-43-0. [Google Scholar]
- Landsteiner, E. Ein Kampf Um Wald Und Weide—Mit Happy End. Die Geschichte Des Mitterretzbacher Kirchenwaldes (1470–2001). Waldviertel 2012, 61, 144–157. [Google Scholar]
- Fineschi, S.; Taurchini, D.; Grossoni, P.; Petit, R.J.; Vendramin, G.G. Chloroplast DNA Variation of White Oaks in Italy. For. Ecol. Manag. 2002, 156, 103–114. [Google Scholar] [CrossRef]
- Neophytou, C.; Gärtner, S.M.; Vargas-Gaete, R.; Michiels, H.-G. Genetic Variation of Central European Oaks: Shaped by Evolutionary Factors and Human Intervention? Tree Genet. Genomes 2015, 11, 79. [Google Scholar] [CrossRef]
- Mazal, L.; Corenblit, D.; Barsoum, N.; Steiger, J.; Skot, L.; Fumanal, B.; Till-Bottraud, I. Fine-Scale Spatial Genetic Structure and Intra-Specific Interactions of Populus Nigra within a Natural River Corridor along the Lower Allier River (France). Flora 2021, 275, 151763. [Google Scholar] [CrossRef]
- Paluch, J.; Zarek, M.; Kempf, M. The Effect of Population Density on Gene Flow between Adult Trees and the Seedling Bank in Abies Alba Mill. Eur. J. For. Res. 2019, 138, 203–217. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon Isotope Discrimination and Photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Valladares, F.; Martinez-Ferri, E.; Balaguer, L.; Perez-Corona, E.; Manrique, E. Low Leaf-level Response to Light and Nutrients in Mediterranean Evergreen Oaks: A Conservative Resource-use Strategy? New Phytol. 2000, 148, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Doležal, J.; Mazůrek, P.; Klimešová, J. Oak Decline in Southern Moravia: The Association between Climate Change and Early and Late Wood Formation in Oaks. Preslia 2010, 82, 289–306. [Google Scholar]
- Piñar Fuentes, J.C.; Cano-Ortiz, A.; Musarella, C.M.; Quinto Canas, R.; Pinto Gomes, C.J.; Spampinato, G.; del Río, S.; Cano, E. Bioclimatology, Structure, and Conservation Perspectives of Quercus Pyrenaica, Acer Opalus Subsp. Granatensis, and Corylus Avellana Deciduous Forests on Mediterranean Bioclimate in the South-Central Part of the Iberian Peninsula. Sustainability 2019, 11, 6500. [Google Scholar] [CrossRef]
- Hall, S.J.; Asner, G.P. Biological Invasion Alters Regional Nitrogen-Oxide Emissions from Tropical Rainforests. Glob. Change Biol. 2007, 13, 2143–2160. [Google Scholar] [CrossRef]
- Hall, R.D.; D’Auria, J.C.; Silva Ferreira, A.C.; Gibon, Y.; Kruszka, D.; Mishra, P.; Van De Zedde, R. High-Throughput Plant Phenotyping: A Role for Metabolomics? Trends Plant Sci. 2022, 27, 549–563. [Google Scholar] [CrossRef]
- Chen, Z.; Li, S.; Wan, X.; Liu, S. Strategies of Tree Species to Adapt to Drought from Leaf Stomatal Regulation and Stem Embolism Resistance to Root Properties. Front. Plant Sci. 2022, 13, 926535. [Google Scholar] [CrossRef]
- Beguería, S.; Vicente-Serrano, S.M.; Reig, F.; Latorre, B. Standardized precipitation evapotranspiration index (SPEI) revisited: Parameter fitting, evapotranspiration models, tools, datasets and drought monitoring. Int. J. Climatol. 2014, 34, 3001–3023. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Loo, M.; Ufimov, R.; Grabner, M.; Übl, C.; Watzinger, A.; Irauschek, F.; Konrad, H.; Píšová, S.; Trujillo-Moya, C. Quercus petraea (Matt.) Liebl. from the Thayatal National Park in Austria: Selection of Potentially Drought-Tolerant Phenotypes. Forests 2023, 14, 2225. https://doi.org/10.3390/f14112225
van Loo M, Ufimov R, Grabner M, Übl C, Watzinger A, Irauschek F, Konrad H, Píšová S, Trujillo-Moya C. Quercus petraea (Matt.) Liebl. from the Thayatal National Park in Austria: Selection of Potentially Drought-Tolerant Phenotypes. Forests. 2023; 14(11):2225. https://doi.org/10.3390/f14112225
Chicago/Turabian Stylevan Loo, Marcela, Roman Ufimov, Michael Grabner, Christian Übl, Andrea Watzinger, Florian Irauschek, Heino Konrad, Soňa Píšová, and Carlos Trujillo-Moya. 2023. "Quercus petraea (Matt.) Liebl. from the Thayatal National Park in Austria: Selection of Potentially Drought-Tolerant Phenotypes" Forests 14, no. 11: 2225. https://doi.org/10.3390/f14112225
APA Stylevan Loo, M., Ufimov, R., Grabner, M., Übl, C., Watzinger, A., Irauschek, F., Konrad, H., Píšová, S., & Trujillo-Moya, C. (2023). Quercus petraea (Matt.) Liebl. from the Thayatal National Park in Austria: Selection of Potentially Drought-Tolerant Phenotypes. Forests, 14(11), 2225. https://doi.org/10.3390/f14112225