Detection and Quantification of Serpula himantioides in the Wood of Chamaecyparis pisifera Butt Rot Trees by Real-Time PCR


Abstract
:1. Introduction
2. Materials and Methods
2.1. Design of Species-Specific Primer Set
2.2. Verification of Specificity by End-Point PCR
2.3. Verification of Specificity by Amplicon Sequencing
2.4. Real-Time PCR Experiments
3. Results
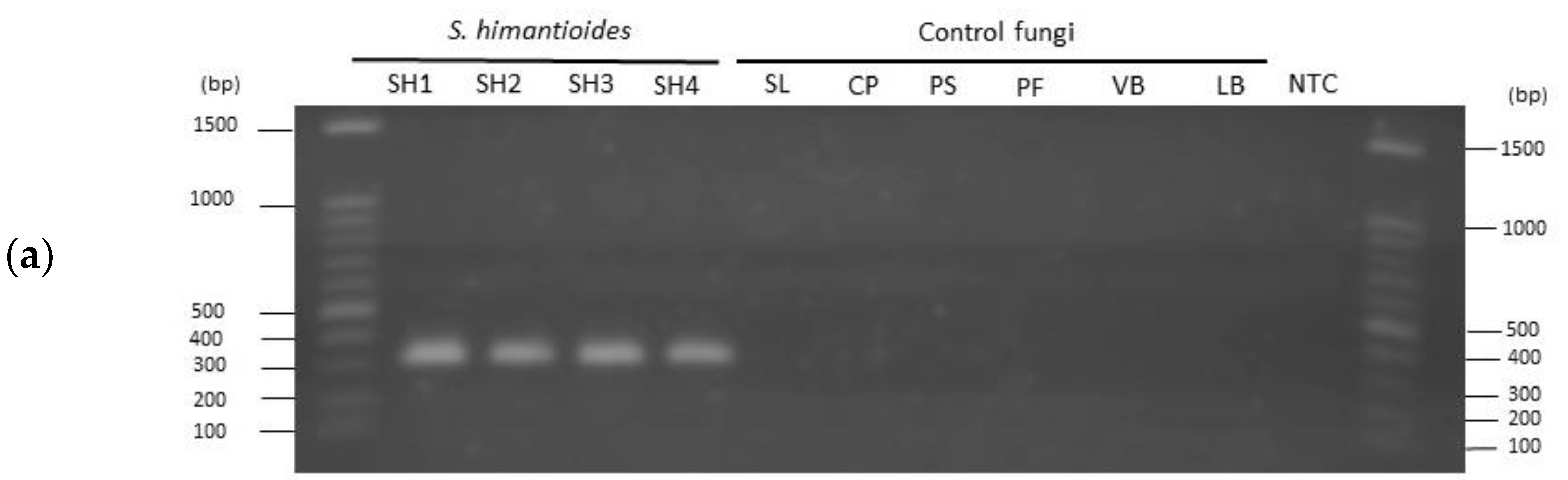
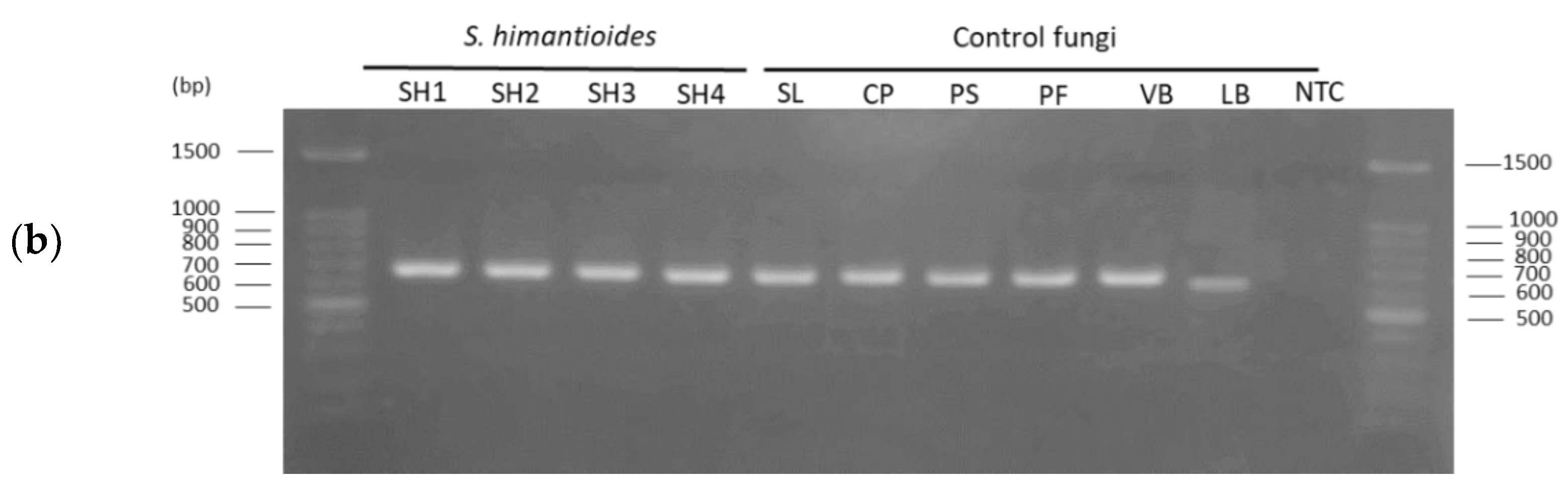
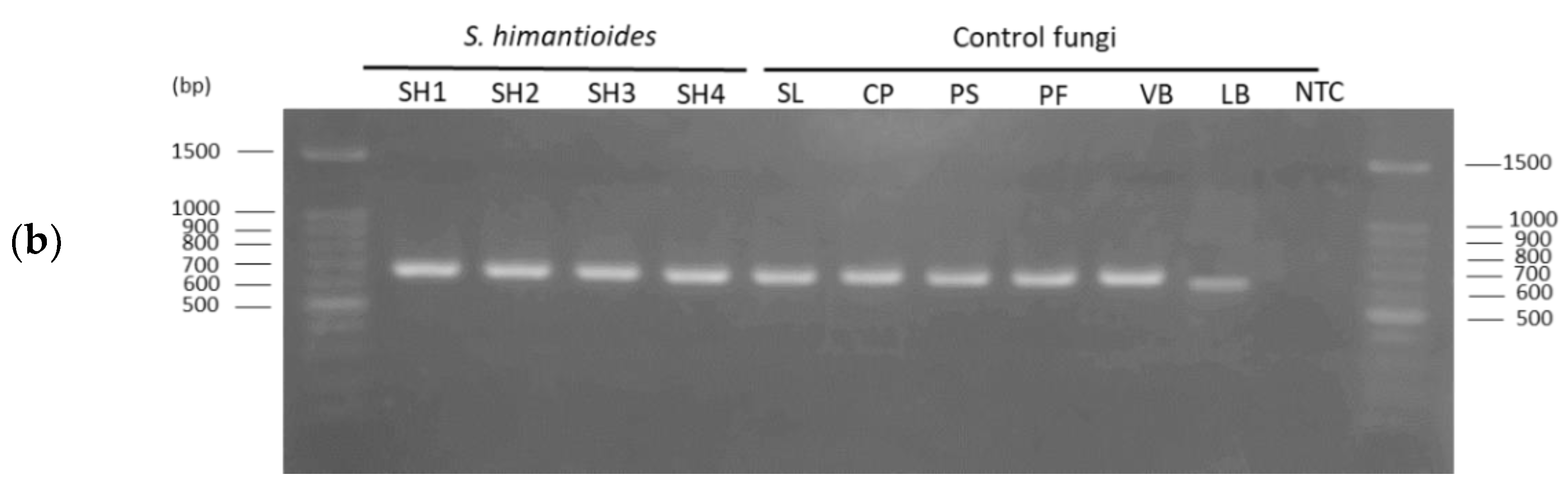
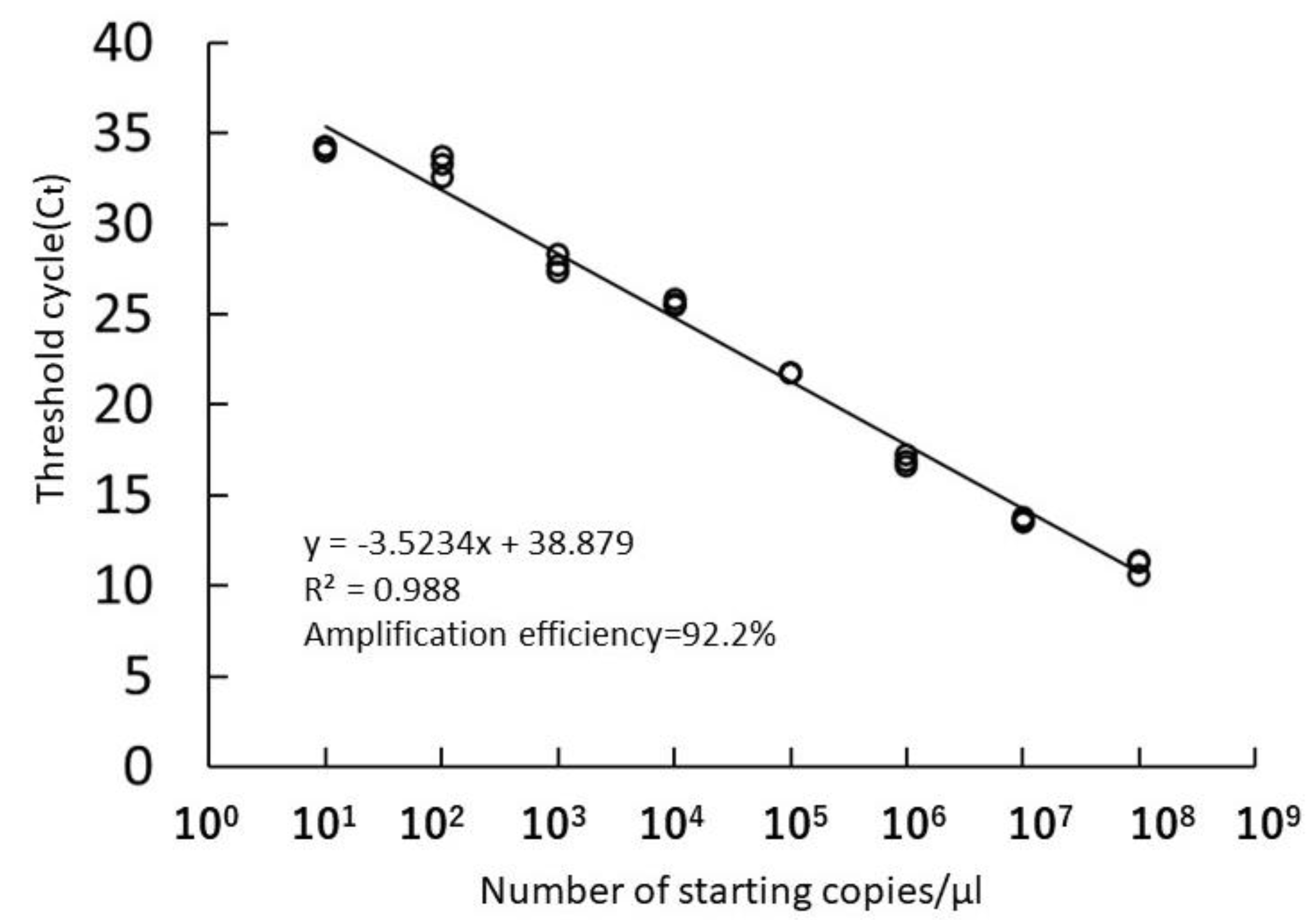
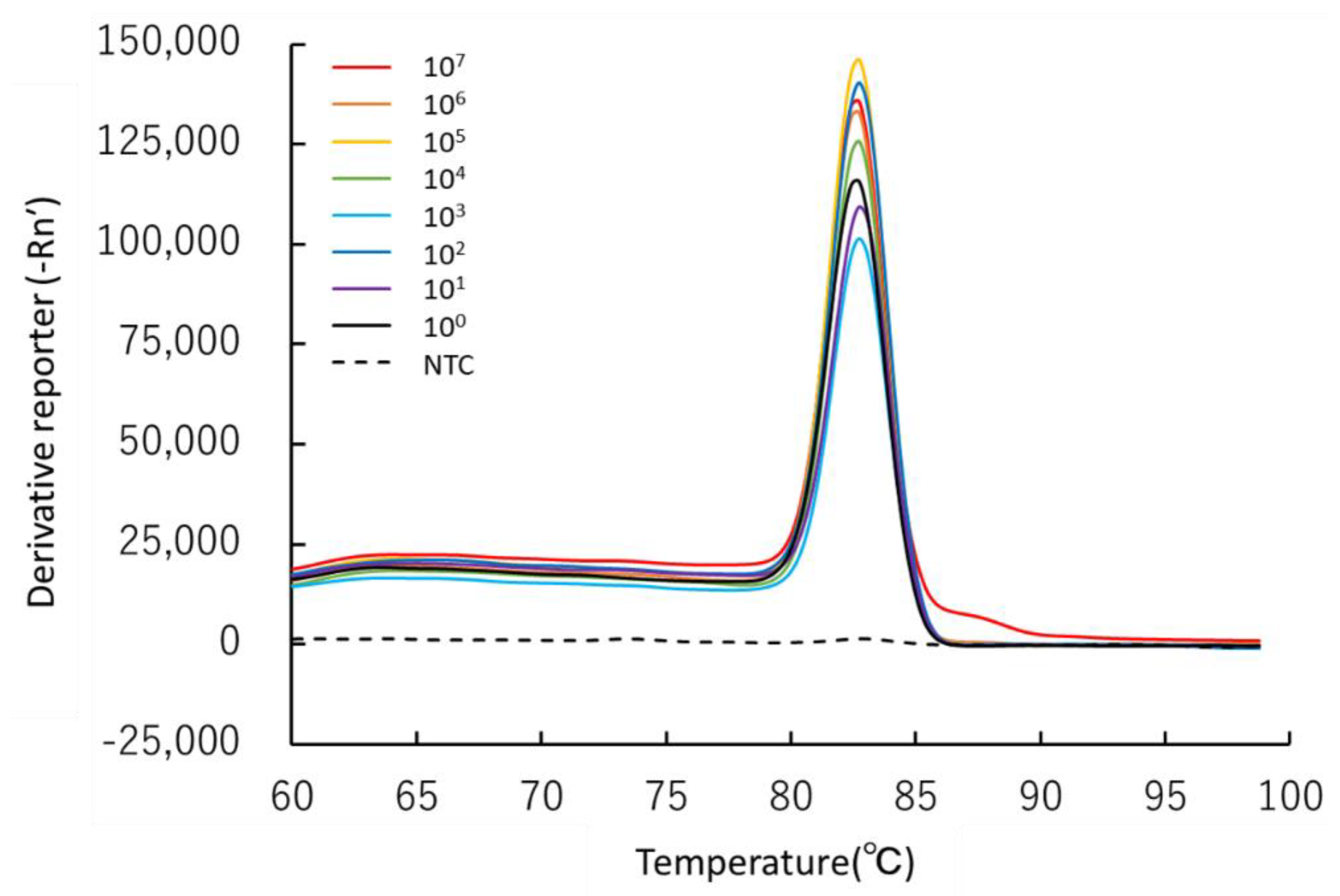
3.1. Specificity of the Newly Developed Primer Set
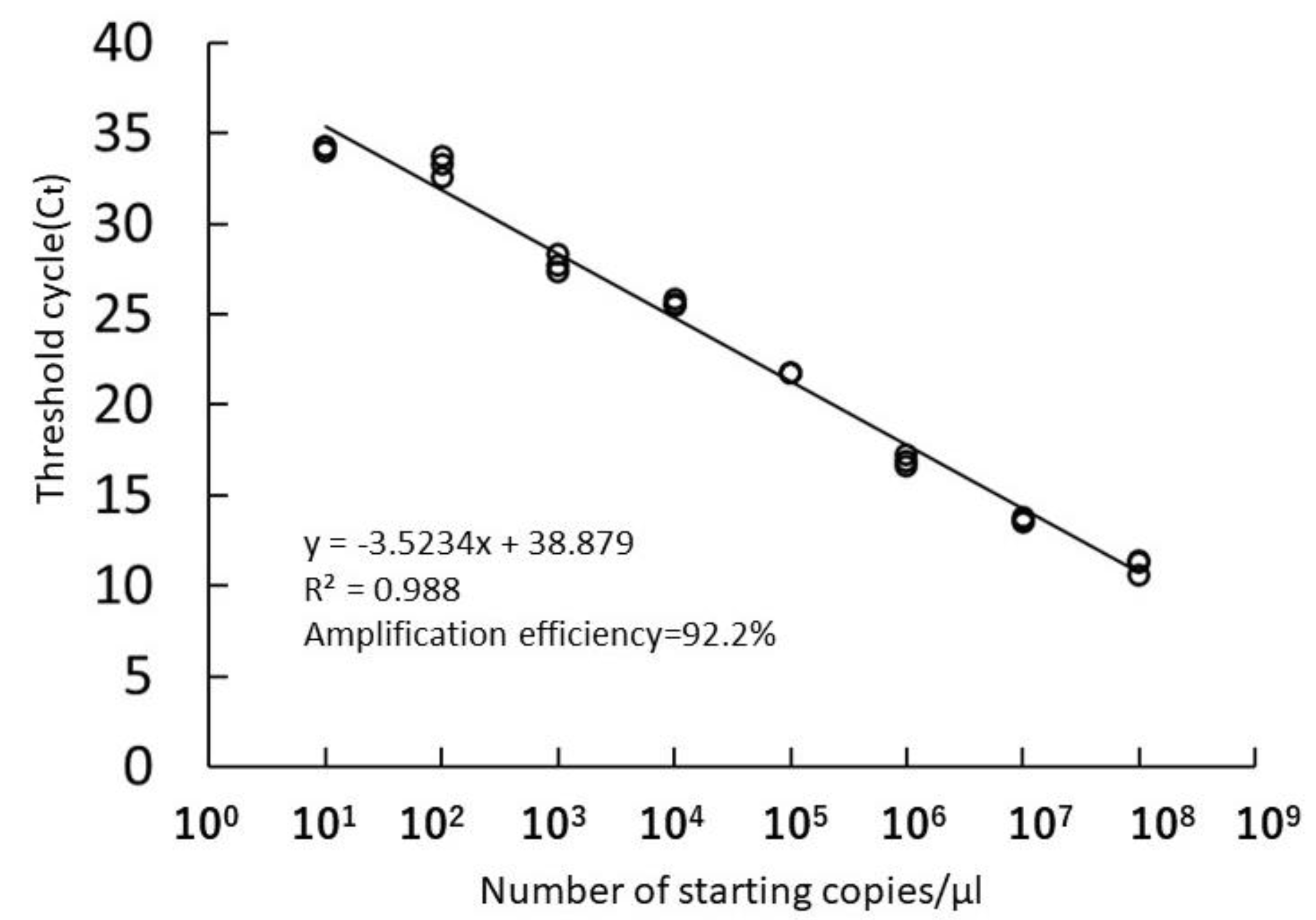
3.2. Sensitivity of the Real-Time PCR Assay for Environmental Samples
4. Discussion
4.1. Specificity of the Designed Primer Set
4.2. Sensitivity of the Real-Time PCR Assay for Environmental Samples
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicolotti, G.; Gonthier, P.; Guglielmo, F. Advances in Detection and Identification of Wood Rotting Fungi in Timber and Standing Trees. In Molecular Identification of Fungi; Gherbawy, Y., Voight, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 251–276. [Google Scholar] [CrossRef]
- Woodward, S.; Stenlid, J.; Karjalainen, R.; Hüttermann, A. Heterobasidion Annosum, Biology, Ecology, Impact and Control; CAB International: Oxon, UK, 1998; pp. xi–xii. [Google Scholar]
- Regue, A.; Bassie, L.; de-Miguel, S.; Colinas, C. Environmental and stand conditions related to Fistulina hepatica heart rot attack on Castanea sativa. For. Pathol. 2019, 49, 7. [Google Scholar] [CrossRef]
- Tomikawa, Y.; Iwase, Y.; Arita, K.; Yamada, H. Nondestructive inspection of a wooden pole using ultrasonic computed tomography. IEEE Trans. Ultrason. Ferroelectr. Freq. Control 1986, 33, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Habermehl, A.; Ridder, H.W.; Seidl, P. Computerized tomographic systems as tools for diagnosing urban tree health. Int. Symp. Urban Tree Health 1999, 496, 261–268. [Google Scholar] [CrossRef]
- Nicolotti, G.; Socco, L.V.; Martinis, R.; Godio, A.; Sambuelli, L. Application and comparison of three tomographic techniques for detection of decay in trees. J. Arbor. 2003, 29, 66–78. [Google Scholar] [CrossRef]
- Muller, U.; Bammer, R.; Halmschlager, E.; Stollberger, R.; Wimmer, R. Detection of fungal wood decay using magnetic resonance imaging. Holz. Als Roh-Und Werkst. 2001, 59, 190–194. [Google Scholar] [CrossRef]
- Sambuelli, L.; Socco, L.V.; Godio, A.; Nicolotti, G.; Martinis, R. Ultrasonic, electric and radar measurements for living tree assessment. Boll. Di Geofis. Teor. Ed Appl. 2003, 44, 253–279. [Google Scholar]
- Guglielmo, F.; Bergemann, S.E.; Gonthier, P.; Nicolotti, G.; Garbelotto, M. A multiplex PCR-based method for the detection and early identification of wood rotting fungi in standing trees. J. Appl. Microbiol. 2007, 103, 1490–1507. [Google Scholar] [CrossRef]
- Schena, L.; Nigro, F.; Ippolito, A.; Gallitelli, D. Real-time quantitative PCR: A new technology to detect and study phytopathogenic and antagonistic fungi. Eur. J. Plant. Pathol. 2004, 110, 893–908. [Google Scholar] [CrossRef]
- Carlsen, T.; Engh, I.B.; Decock, C.; Rajchenberg, M.; Kauserud, H. Multiple cryptic species with divergent substrate affinities in the Serpula himantioides species complex. Fungal Biol. 2011, 115, 54–61. [Google Scholar] [CrossRef]
- Seehann, G. Butt rot in conifers caused by Serpula himantioides. Karst. For. Pathol. 1986, 16, 207–217. [Google Scholar]
- Koch, J.; Thomsen, I.M. Serpula himantioides, Heterobasidion annosum and Calocera viscosa as butt rot fungi in a Danish Douglas-fir stand. For. Pathol. 2003, 33, 1–6. [Google Scholar] [CrossRef]
- Whitney, R.D. Root-rotting fungi in white spruce, black spruce, and balsam fir in northern Ontario. Can. J. For. Res. 1995, 25, 1209–1230. [Google Scholar] [CrossRef]
- Rizzo, D.M.; Harrington, T.C. Root and butt rot fungi on balsam fir and red spruce in the White Mountains, New Hampshire. Plant Dis. 1988, 72, 329–331. [Google Scholar] [CrossRef]
- Haraguchi, R.; Hirao, T.; Yamada, T. Butt-rot damage situation of Chamaecyparis pisifera in the University of Tokyo Chichibu Forest. Kanto J. For. Res. 2017, 68, 181–184. (In Japanese) [Google Scholar]
- Abe, Y. Some properties of Chamaecyparis pisifera butt rot fungus Serpula himantioides. Trans. JPN For. Soc. 2002, 113, 87. (In Japanese) [Google Scholar]
- Schmidt, O. Indoor wood-decay basidiomycetes: Damage, causal fungi, physiology, identification and characterization, prevention and control. Mycol. Prog. 2007, 6, 261–279. [Google Scholar] [CrossRef]
- Boehm, J.; Hahn, A.; Schubert, R.; Bahnweg, G.; Adler, N.; Nechwatal, J.; Oehlmann, R.; Osswald, W. Real-time quantitative PCR: DNA determination in isolated spores of the mycorrhizal fungus Glomus mosseae and monitoring of Phytophthora infestans and Phytophthora citricola in their respective host plants. J. Phytopathol. 1999, 147, 409–416. [Google Scholar] [CrossRef]
- Luchi, N.; Capretti, P.; Pinzani, P.; Orlando, C.; Pazzagli, M. Real-time PCR detection of Biscogniauxia mediterranea in symptomless oak tissue. Lett. Appl. Microbiol. 2005, 41, 61–68. [Google Scholar] [CrossRef]
- Luchi, N.; Capretti, P.; Vettraino, A.M.; Vannini, A.; Pinzani, P.; Pazzagli, M. Early detection of Biscogniauxia nummularia in symptomless European beech (Fagus sylvatica L.) by TaqMan (TM) quantitative real-time PCR. Lett. Appl. Microbiol. 2006, 43, 33–38. [Google Scholar] [CrossRef]
- Filion, M.; St-Arnaud, M.; Jabaji-Hare, S.H. Direct quantification of fungal DNA from soil substrate using real-time PCR. J. Microbiol. Methods 2003, 53, 67–76. [Google Scholar] [CrossRef]
- Berry, S.D.; Fargette, M.; Spaull, V.W.; Morand, S.; Cadet, P. Detection and quantification of root-knot nematode (Meloidogyne javanica), lesion nematode (Pratylenchus zeae) and dagger nematode (Xiphinema elongatum) parasites of sugarcane using real-time PCR. Mol. Cell. Probes 2008, 22, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Schweigkofler, W.; O’Donnell, K.; Garbelotto, M. Detection and quantification of airborne conidia of Fusarium circinatum, the causal agent of pine pitch canker, from two California sites by using a real-time PCR approach combined with a simple spore trapping method. Appl. Environ. Microbiol. 2004, 70, 3512–3520. [Google Scholar] [CrossRef] [PubMed]
- Luchi, N.; Ghelardini, L.; Belbahri, L.; Quartier, M.; Santini, A. Rapid Detection of Ceratocystis platani Inoculum by Quantitative Real-Time PCR Assay. Appl. Environ. Microbiol. 2013, 79, 5394–5404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonthier, P.; Guglielmo, F.; Sillo, F.; Giordano, L.; Garbelotto, M. A molecular diagnostic assay for the detection and identification of wood decay fungi of conifers. For. Pathol. 2015, 45, 89–101. [Google Scholar] [CrossRef]
- Horisawa, S.; Sakuma, Y.; Doi, S. Qualitative and quantitative PCR methods using species-specific primer for detection and identification of wood rot fungi. J. Wood Sci. 2009, 55, 133–138. [Google Scholar] [CrossRef]
- Eikenes, M.; Hietala, A.; Alfredsen, G.; Fossdal, C.G.; Solheim, H. Comparison of chitin, ergosterol and Real-Time PCR based assays for monitoring colonization of Trametes versicolor in birch wood. Holzforschung 2005, 59, 568–573. [Google Scholar] [CrossRef]
- Schmidt, O.; Moreth, U. Species-specific PCR primers in the rDNA-ITS region as a diagnostic tool for Serpula lacrymans. Mycol. Res. 2000, 104, 69–72. [Google Scholar] [CrossRef]
- OligoEvaluator™. Available online: http://www.oligoevaluator.com/ (accessed on 7 June 2021).
- National Center for Biotechnology Information Home Page. Available online: http://www.ncbi.nlm.nih.gov/BLAST/ (accessed on 7 June 2021).
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In Pcr Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Ihrmark, K.; Bodeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandstrom-Durling, M.; Clemmensen, K.E.; et al. New primers to amplify the fungal ITS2 region—evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Polme, S.; Koljalg, U.; Yorou, N.S.; Wijesundera, R.; Villarreal Ruiz, L.; Vasco-Palacios, A.M.; Pham Quang, T.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef]
- Gweon, H.S.; Oliver, A.; Taylor, J.; Booth, T.; Gibbs, M.; Read, D.S.; Griffiths, R.I.; Schonrogge, K. PIPITS: An automated pipeline for analyses of fungal internal transcribed spacer sequences from the Illumina sequencing platform. Methods Ecol. Evol. 2015, 6, 973–980. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; De Wit, P.; Sanchez-Garcia, M.; Ebersberger, I.; de Sousa, F.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 4, 914–919. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Hoiland, K.; Kjoller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi—Recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef]
- Bradley, I.M.; Pinto, A.J.; Guest, J.S. Design and Evaluation of Illumina MiSeq-Compatible, 18S rRNA Gene-Specific Primers for Improved Characterization of Mixed Phototrophic Communities. Appl. Environ. Microbiol. 2016, 82, 5878–5891. [Google Scholar] [CrossRef]
- Skrede, I.; Engh, I.B.; Binder, M.; Carlsen, T.; Kauserud, H.; Bendiksby, M. Evolutionary history of Serpulaceae (Basidiomycota): Molecular phylogeny, historical biogeography and evidence for a single transition of nutritional mode. BMC Evol. Biol. 2011, 11, 230. [Google Scholar] [CrossRef] [PubMed]
- Kauserud, H.; Hogberg, N.; Knudsen, H.; Elborne, S.A.; Schumacher, T. Molecular phylogenetics suggest a North American link between the anthropogenic dry rot fungus Serpula lacrymans and its wild relative S-himantioides. Mol. Ecol. 2004, 13, 3137–3146. [Google Scholar] [CrossRef]
- Ota, H.; Matsushita, N.; Haraguchi, R.; Yamada, T.; Fukuda, K. A new cryptic species of the dry rot fungus Serpula himantioides found in Japan. JPN For. Soc. Congr. 2019, 130, 172. (In Japanese) [Google Scholar] [CrossRef]
- Prevost-Boure, N.C.; Christen, R.; Dequiedt, S.; Mougel, C.; Lelievre, M.; Jolivet, C.; Shahbazkia, H.R.; Guillou, L.; Arrouays, D.; Ranjard, L. Validation and Application of a PCR Primer Set to Quantify Fungal Communities in the Soil Environment by Real-Time Quantitative PCR. PLoS ONE 2011, 6, e24166. [Google Scholar] [CrossRef]
- Thonar, C.; Erb, A.; Jansa, J. Real-time PCR to quantify composition of arbuscular mycorrhizal fungal communities—marker design, verification, calibration and field validation. Mol. Ecol. Resour. 2012, 12, 219–232. [Google Scholar] [CrossRef]
- van der Wal, A.; Gunnewiek, P.K.; de Hollander, M.; de Boer, W. Fungal diversity and potential tree pathogens in decaying logs and stumps. For. Ecol. Manag. 2017, 406, 266–273. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Basidiomycetes | Ascomycetes (%) | Other Fungi (%) | |||
|---|---|---|---|---|---|---|
| S. himantioides (%) | S. lacrymans (%) | Serpula spp. (%) | Besides Serpula (%) | |||
| DWS_1 | 45.83 | 0.00 | 52.57 | 0.06 | 1.49 | 0.05 |
| DWS_2 | 47.78 | 0.00 | 51.86 | 0.05 | 0.26 | 0.05 |
| DWS_3 | 48.58 | 0.00 | 45.34 | 3.52 | 0.51 | 2.06 |
| DWS_4 | 47.16 | 0.00 | 50.25 | 1.29 | 0.22 | 1.08 |
| DWS_5 | 49.08 | 0.00 | 48.88 | 0.72 | 1.29 | 0.04 |
| DWS_6 | 27.54 | 0.00 | 29.23 | 18.32 | 24.40 | 0.51 |
| DWS_7 | 16.52 | 0.17 | 16.94 | 19.79 | 37.98 | 8.60 |
| DWS_8 | 13.36 | 0.31 | 13.61 | 31.61 | 33.15 | 7.96 |
| DWS_9 | 7.37 | 0.24 | 7.52 | 42.25 | 31.73 | 10.90 |
| Sample Name | Basidiomycetes | Ascomycetes (%) | Other Fungi (%) | |||
|---|---|---|---|---|---|---|
| S. himantioides (%) | S. lacrymans (%) | Serpula spp. (%) | Besides Serpula (%) | |||
| DWS_1 | 91.02 | 0.00 | 8.98 | 0.00 | 0.00 | 0.00 |
| DWS_2 | 90.86 | 0.00 | 9.14 | 0.00 | 0.00 | 0.00 |
| DWS_3 | 92.05 | 0.00 | 7.95 | 0.00 | 0.00 | 0.00 |
| DWS_4 | 90.52 | 0.00 | 9.48 | 0.00 | 0.00 | 0.00 |
| DWS_5 | 91.83 | 0.00 | 8.17 | 0.00 | 0.00 | 0.00 |
| DWS_6 | 92.22 | 0.00 | 7.78 | 0.00 | 0.00 | 0.00 |
| DWS_7 | 91.40 | 0.00 | 8.60 | 0.00 | 0.00 | 0.00 |
| DWS_8 | 92.03 | 0.00 | 7.97 | 0.00 | 0.00 | 0.00 |
| DWS_9 | 87.34 | 0.00 | 12.66 | 0.00 | 0.00 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haraguchi, R.; Hirao, T.; Yamada, T. Detection and Quantification of Serpula himantioides in the Wood of Chamaecyparis pisifera Butt Rot Trees by Real-Time PCR. Forests 2022, 13, 1429. https://doi.org/10.3390/f13091429
Haraguchi R, Hirao T, Yamada T. Detection and Quantification of Serpula himantioides in the Wood of Chamaecyparis pisifera Butt Rot Trees by Real-Time PCR. Forests. 2022; 13(9):1429. https://doi.org/10.3390/f13091429
Chicago/Turabian StyleHaraguchi, Ryusei, Toshihide Hirao, and Toshihiro Yamada. 2022. "Detection and Quantification of Serpula himantioides in the Wood of Chamaecyparis pisifera Butt Rot Trees by Real-Time PCR" Forests 13, no. 9: 1429. https://doi.org/10.3390/f13091429
APA StyleHaraguchi, R., Hirao, T., & Yamada, T. (2022). Detection and Quantification of Serpula himantioides in the Wood of Chamaecyparis pisifera Butt Rot Trees by Real-Time PCR. Forests, 13(9), 1429. https://doi.org/10.3390/f13091429