


Unexpectedly, Creation of Temporary Water Bodies Has Increased the Availability of Food and Nesting Sites for Bees (Apiformes)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
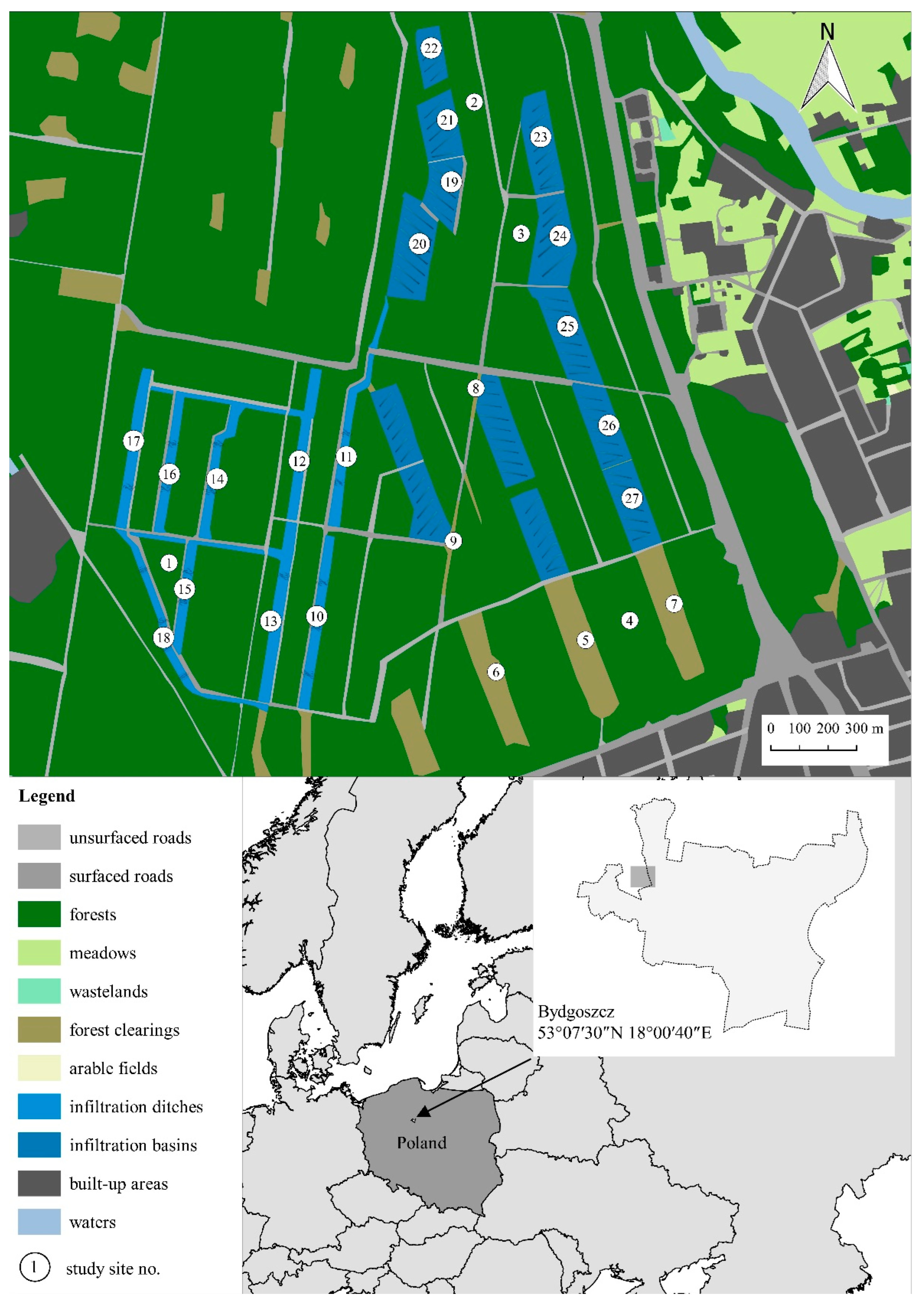
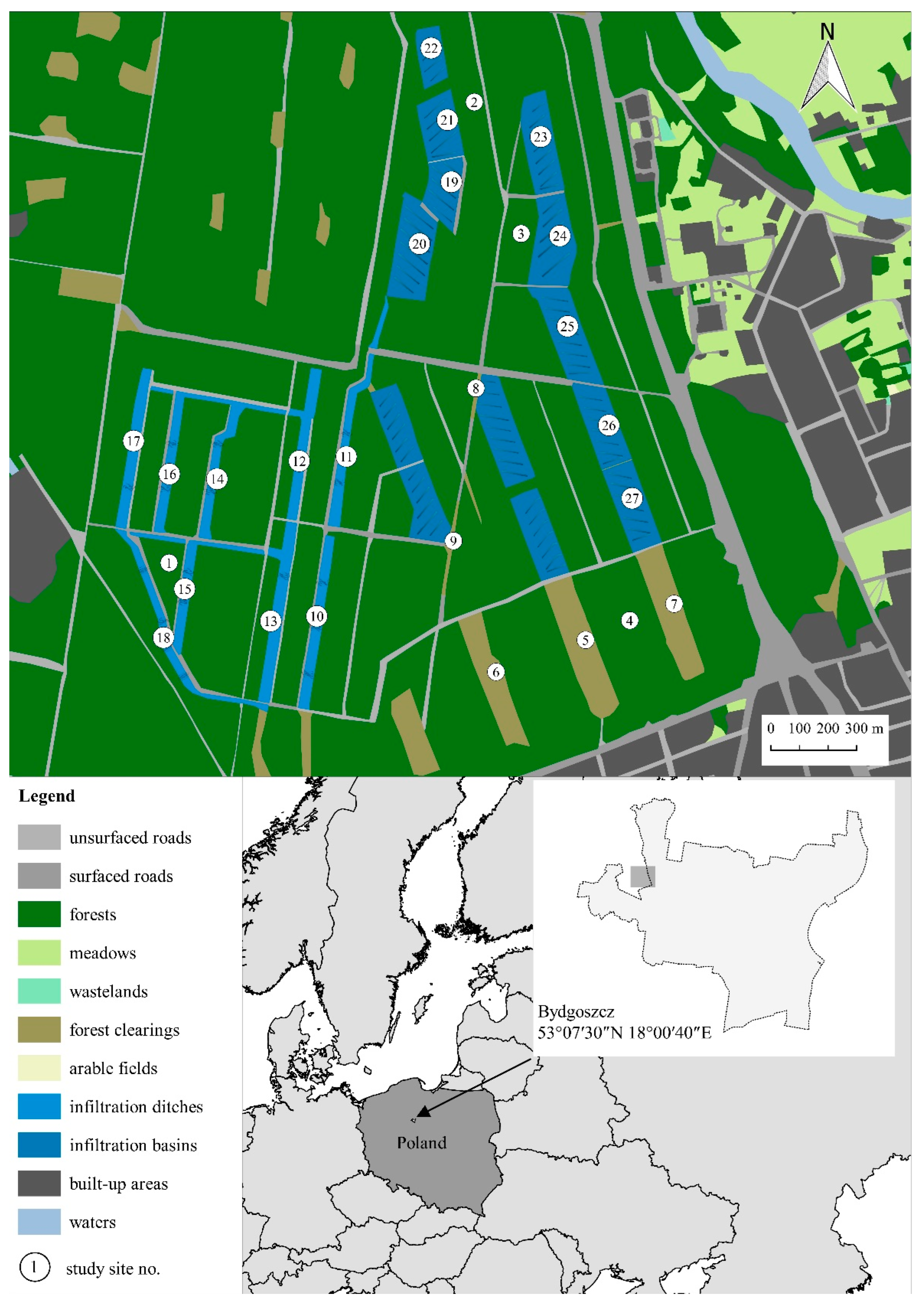
2.1. Study Area
2.2. Bee Sampling
2.3. Floristic Records
2.4. Environmental Variables at the Level of Habitat and Landscape
2.5. Statistical Analyses
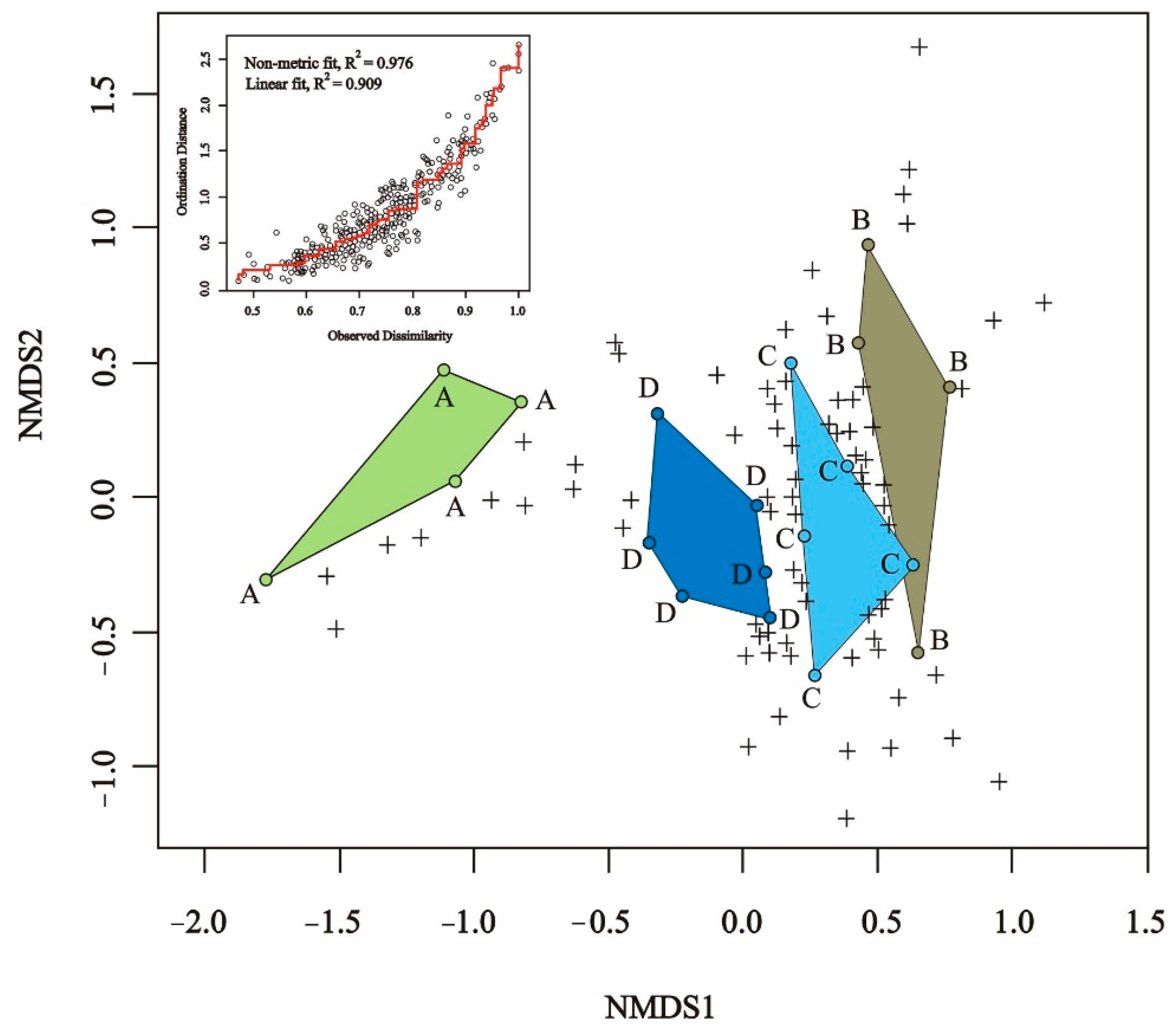
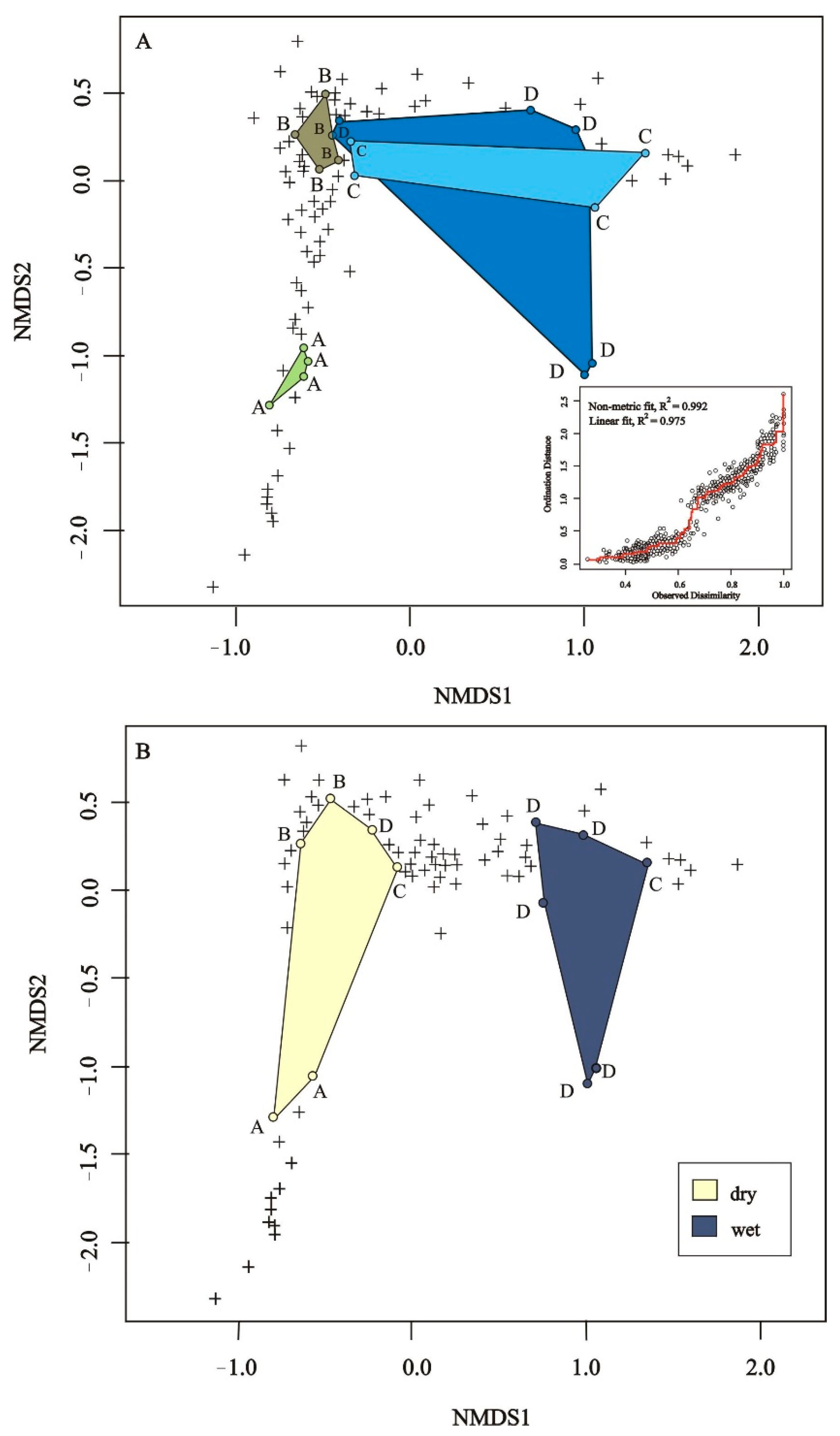
3. Results
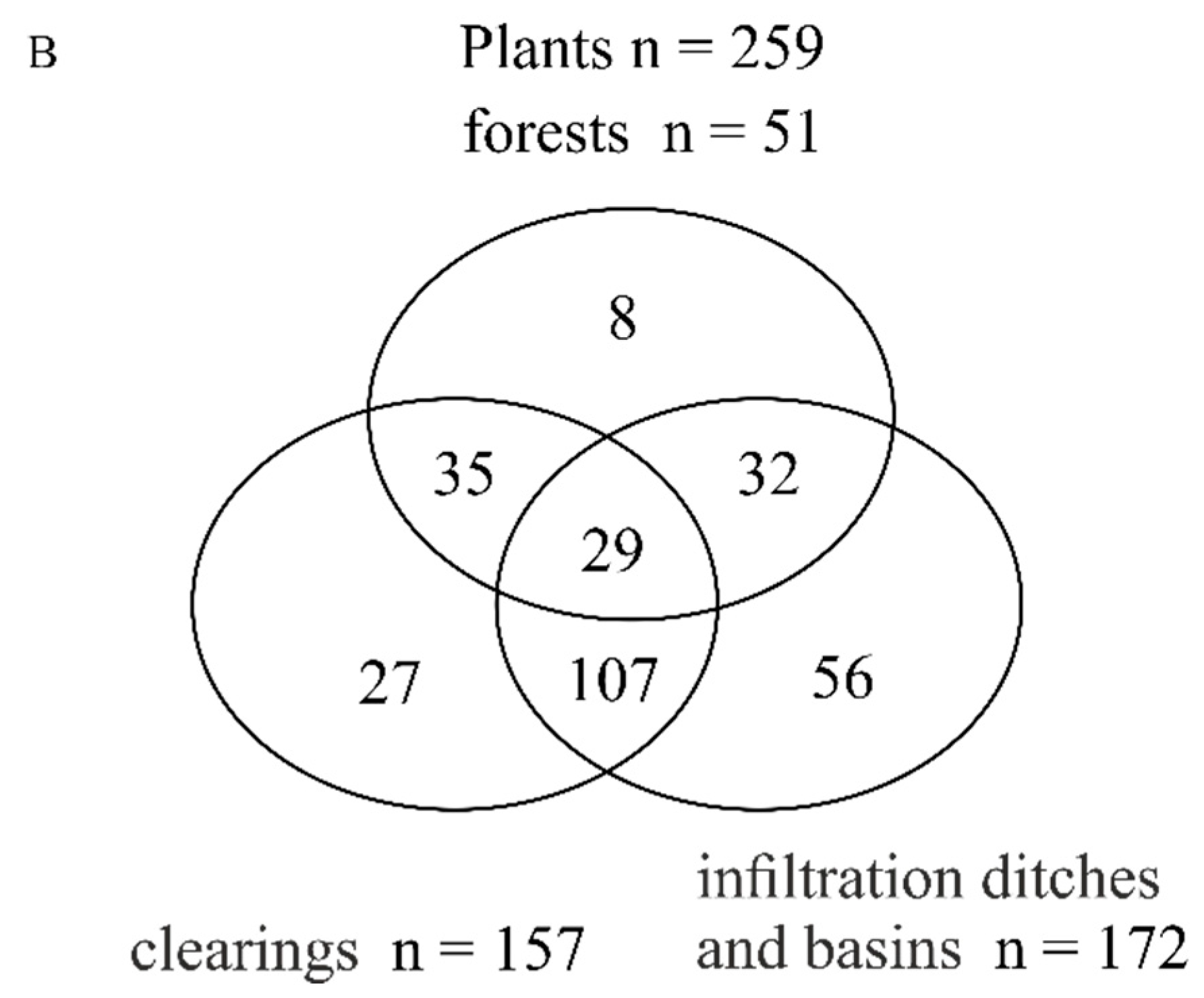
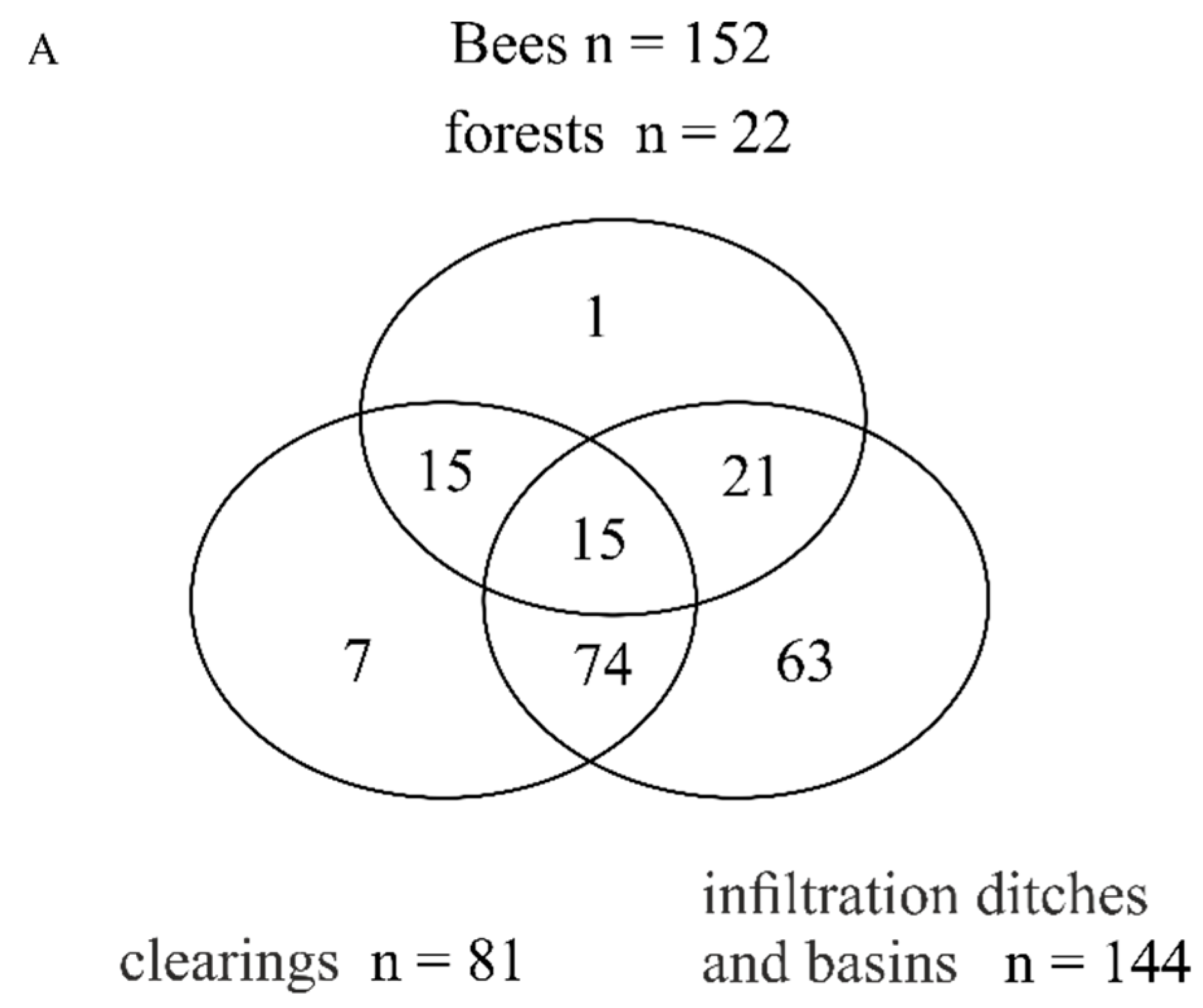
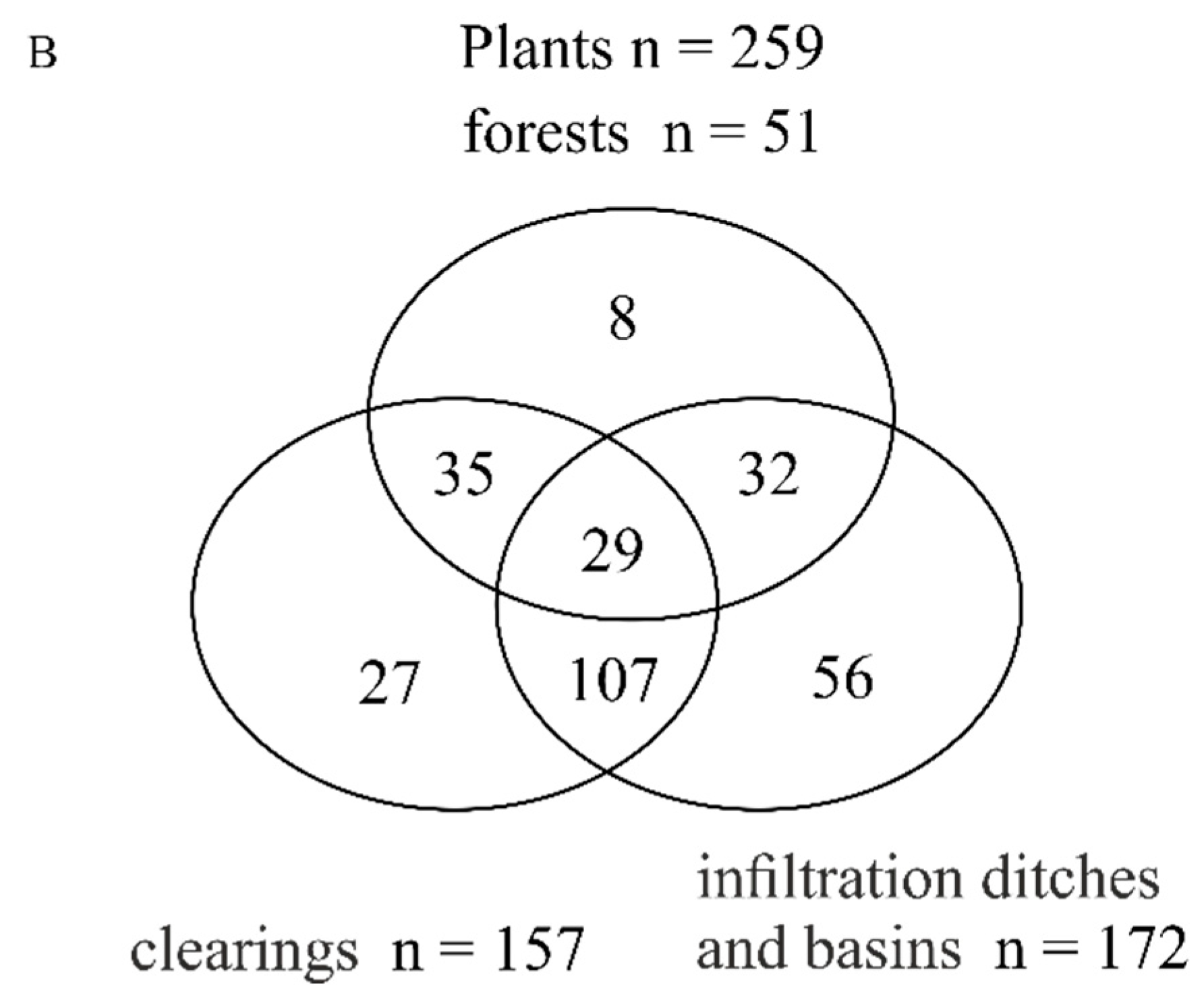
3.1. Total Species Richness of Bees and Plants
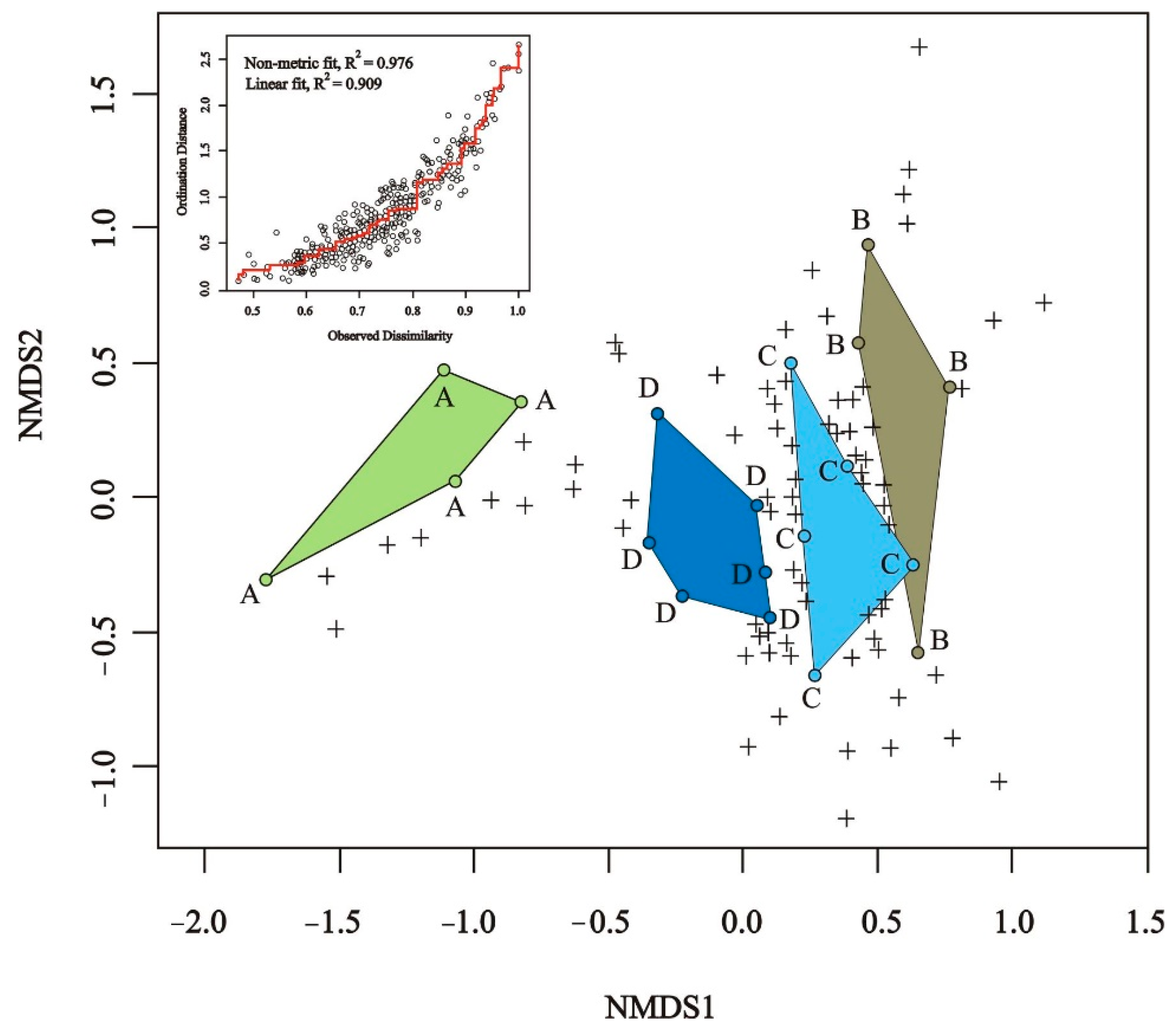
3.2. Bee Species Composition
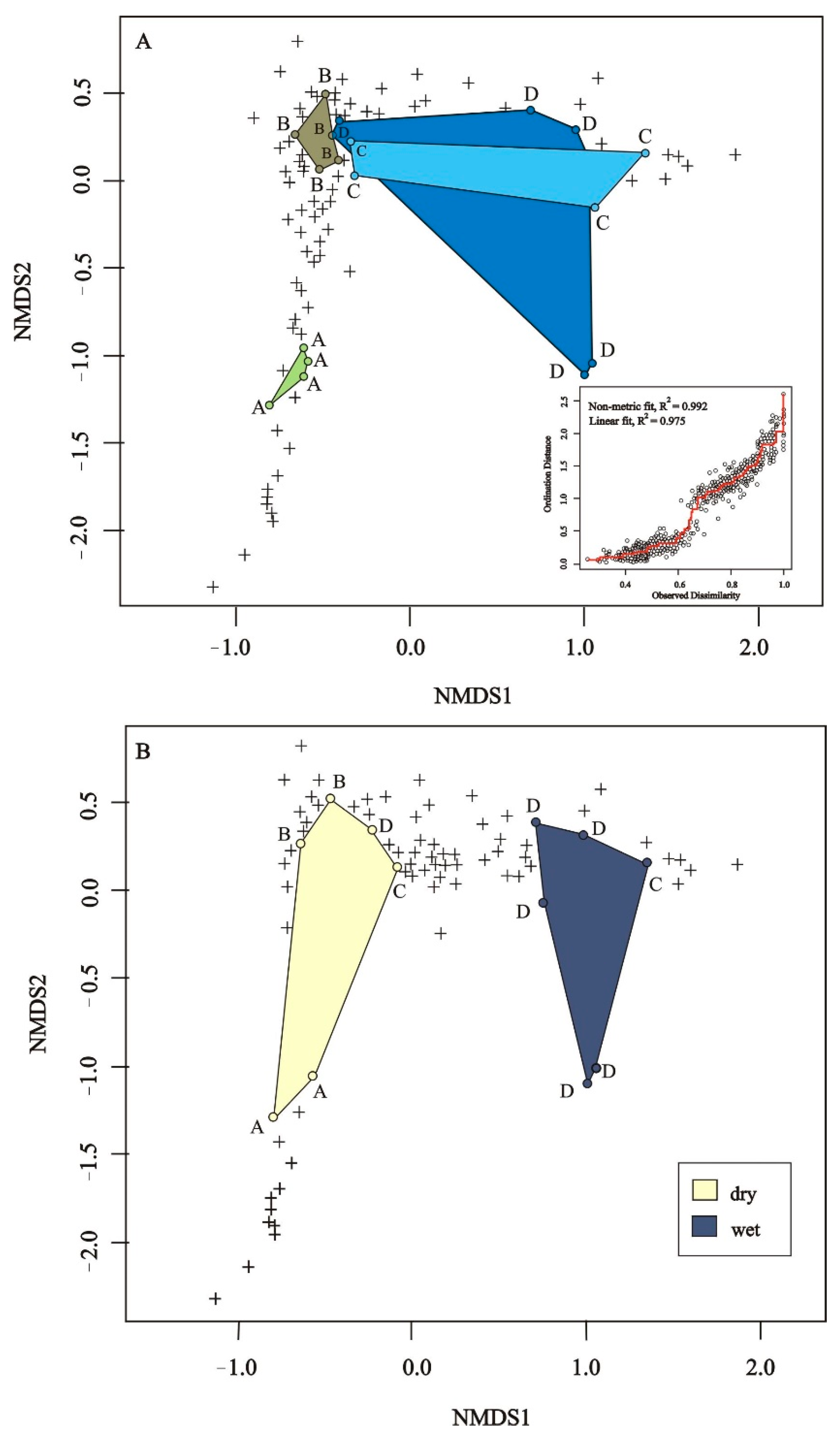
3.3. Plant Species Composition
3.4. Changes in Bee Abundance and Species Richness
3.5. Factors Affecting Bee Species Richness and Abundance
4. Discussion
4.1. Infiltration Basins as New Habitats for Bees
4.2. Changes in Natural Resources of Plants and Bees and Habitat Preferences of Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Bai, J.; Wu, X.; Briggs, J.M. Global change and the ecology of cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef] [PubMed]
- Somme, L.; Moquet, L.; Quinet, M.; Vanderplanck, M.; Michez, D.; Lognay, G.; Jacquemart, A.-L. Food in a row: Urban trees offer valuable floral resources to pollinating insects. Urban Ecosyst. 2016, 19, 1149–1161. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [PubMed]
- Kowarik, I. Novel urban ecosystems, biodiversity, and conservation. Environ. Pollut. 2011, 159, 1974–1983. [Google Scholar] [CrossRef]
- Sattler, T.; Obrist, M.K.; Duelli, P.; Moretti, M. Urban arthropod communities: Added value or just a blend of surrounding biodiversity? Landsc. Urban Plan. 2011, 103, 347–361. [Google Scholar] [CrossRef]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in cities needs space: A meta-analysis of factors determining intraurban biodiversity variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar]
- Theodorou, P.; Radzevičiūtė, R.; Lentendu, G.; Kahnt, B.; Husemann, M.; Bleidorn, C.; Settele, J.; Schweiger, O.; Grosse, I.; Wubet, T.; et al. Urban areas as hotspots for bees and pollination but not a panacea for all insects. Nat. Commun. 2020, 11, 576. [Google Scholar] [CrossRef] [PubMed]
- Normandin, É.; Vereecken, N.J.; Buddle, C.M.; Fournier, V. Taxonomic and functional trait diversity of wild bees in different urban settings. PeerJ 2017, 5, e3051. [Google Scholar]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar]
- Steffan-Dewenter, I.; Tscharntke, T. Resource overlap and possible competition between honey bees and wild bees in central Europe. Oecologia 2000, 1222, 288–296. [Google Scholar] [CrossRef]
- Kearns, C.A.; Oliveras, D.M. Environmental factors affecting bee diversity in urban and remote grassland plots in Boulder, Colorado. J. Insect Conserv. 2009, 13, 655–665. [Google Scholar]
- Buchholz, S.; Gathof, A.K.; Grossmann, A.J.; Kowarik, I.; Fischer, L.K. Wild bees in urban grasslands: Urbanisation, functional diversity and species traits. Landsc. Urban Plan. 2020, 196, 103731. [Google Scholar]
- Twerd, L.; Sobieraj-Betlińska, A.; Szefer, P. Roads, railways, and power lines: Are they crucial for bees in urban woodlands? Urban For. Urban Green. 2021, 61, 127120. [Google Scholar]
- Potts, S.G.; Vulliamy, B.; Dafni, A.; Ne’eman, G.; O’Toole, C.; Roberts, S.; Willmer, P. Response of plant-pollinator communities to fire: Changes in diversity, abundance and floral reward structure. Oikos 2003, 101, 103–112. [Google Scholar]
- Tommasi, D.; Miro, A.; Higo, H.A.; Winston, M.L. Bee diversity and abundance in an urban setting. Can. Entomol. 2004, 136, 851–869. [Google Scholar]
- Larson, J.L.; Kesheimer, A.J.; Potter, D.A. Pollinator assemblages on dandelions and white clover in urban and suburban lawns. J. Insect Conserv. 2014, 18, 863–873. [Google Scholar] [CrossRef]
- Hicks, D.M.; Ouvrard, P.; Baldock, K.C.R.; Mark, M.; Goddard, A.; Kunin, W.E.; Mitschunas, N.; Memmott, J.; Morse, H.; Nikolitsi, M.; et al. Food for pollinators: Quantifying the nectar and pollen resources of urban flower meadows. PLoS ONE 2016, 11, e0158117. [Google Scholar]
- Martins, K.T.; Gonzales, A.; Lechowicz, M.J. Patterns of pollinator turnover and increasing diversity associated with urban habitats. Urban Ecosyst. 2017, 20, 1359–1371. [Google Scholar]
- Twerd, L.; Banaszak-Cibicka, W.; Sobieraj-Betlińska, A.; Waldon-Rudzionek, B.; Hoffmann, R. Contributions of phenological groups of wild bees as an indicator of food availability in urban wastelands. Ecol. Indic. 2021, 126, 107616. [Google Scholar] [CrossRef]
- Haaland, C.; Gyllin, M. Butterflies and bumblebees in greenways and sown wildflower strips in Southern Sweden. J. Insect Conserv. 2010, 14, 125–132. [Google Scholar] [CrossRef]
- Matteson, K.C.; Langellotto, G.A. Determinates of inner city butterfly and bee species richness. Urban Ecosyst. 2010, 13, 333–347. [Google Scholar]
- Gunnarsson, B.; Federsel, L.M. Bumble bees in the city: Abundance, species richness and diversity in two urban habitats. J. Insect Conserv. 2014, 18, 1185–1191. [Google Scholar]
- Ellis, E.C.; Antill, E.C.; Kreft, H. All is not loss: Plant biodiversity in the anthropocene. PLoS ONE 2012, 7, e30535. [Google Scholar]
- Smith, R.M.; Thompson, K.; Hodgson, J.G.; Warren, P.H.; Gaston, K.J. Urban domestic gardens (IX): Composition and richness of the vascular plant flora, and implications for native biodiversity. Biol. Conserv. 2006, 129, 312–322. [Google Scholar]
- Bartomeus, I.; Fründ, J.; Williams, N.M. Invasive plants as novel food resources, the pollinators’ perspective. In Biological Invasions and Animal Behavior; Weis, J.S., Sol, D., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 119–132. [Google Scholar]
- Matteson, K.C.; Langellotto, G.A. Small scale additions of native plants fail to increase beneficial insect richness in urban gardens. Insect Conserv. Divers. 2011, 4, 89–98. [Google Scholar]
- Hanley, M.E.; Awbi, A.J.; Franco, M. Going native? Flower use by bumblebees in English urban gardens. Ann. Bot. 2014, 113, 799–806. [Google Scholar] [CrossRef]
- Stewart, A.B.; Sritongchuay, T.; Teartisup, P.; Kaewsomboon, S.; Bumrungsri, S. Habitat and landscape factors influence pollinators in a tropical megacity, Bangkok, Thailand. PeerJ 2018, 6, e5335. [Google Scholar]
- Shochat, E.; Lerman, S.B.; Anderies, J.M.; Warren, P.S.; Faeth, S.H.; Nilon, C.H. Invasion, competition, and biodiversity loss in urban ecosystems. BioScience 2010, 60, 199–208. [Google Scholar] [CrossRef]
- Herzon, I.; Helenius, J. Agricultural drainage ditches, their biological importance and functioning. Biol. Conserv. 2008, 141, 1171–1183. [Google Scholar]
- Hopwood, J.L. The contribution of roadside grassland restorations to native bee conservation. Biol. Conserv. 2008, 141, 2632–2640. [Google Scholar] [CrossRef]
- Hopwood, J.; Winkler, L.; Deal, B.; Chivvis, M. Use of Roadside Prairie Plantings by Native Bees; Living Roadway Trust Fund. 2010. Available online: www.academia.edu/16996737/Use_of_roadside_prairie_plantings_by_native_bees (accessed on 15 October 2021).
- Moroń, D.; Skórka, P.; Lenda, M.; Rożej-Pabijan, E.; Wantuch, M.; Kajzer-Bonk, J.; Celary, W.; Mielczarek, Ł.E.; Tryjanowski, P. Railway embankments as new habitat for pollinators in an agricultural landscape. PLoS ONE 2014, 9, e101297. [Google Scholar] [CrossRef]
- Heneberg, P.; Bogusch, P.; Řezáč, M. Off-road motorcycle circuits support long-term persistence of bees and wasps (Hymenoptera: Aculeata) of open landscape at newly formed refugia within otherwise afforested temperate landscape. Ecol. Eng. 2016, 93, 187–198. [Google Scholar] [CrossRef]
- Hill, B.; Bartomeus, I. The potential of electricity transmission corridors in forested areas as bumblebee habitat. R. Soc. Open Sci. 2016, 3, 160525. [Google Scholar] [CrossRef]
- Heneberg, P.; Bogusch, P.; Řezáč, M. Roadside verges can support spontaneous establishment of steppe-like habitats hosting diverse assemblages of bees and wasps (Hymenoptera: Aculeata) in an intensively cultivated central European landscape. Biodivers. Conserv. 2017, 26, 843–864. [Google Scholar] [CrossRef]
- Moroń, D.; Tryjanowski, P.; Celary, W.; Lenda, M.; Skórka, P. Railway lines affect spatial turnover of pollinator communities in an agricultural landscape. Biodivers. Res. 2017, 23, 1090–1097. [Google Scholar] [CrossRef]
- Wagner, D.L.; Metzler, K.J.; Frye, H. Importance of transmission line corridors for conservation of native bees and other wildlife. Biol. Conserv. 2019, 235, 147–156. [Google Scholar] [CrossRef]
- Krauss, J.; Alfert, T.; Steffan-Dewenter, I. Habitat area but not habitat age determines wild bee richness in limestone quarries. J. Appl. Ecol. 2009, 46, 194–202. [Google Scholar] [CrossRef]
- Tropek, R.; Spitzer, L.; Konvicka, M. Two groups of epigeic arthropods differ in colonising of piedmont quarries: The necessity of multi-taxa and life-history traits approaches in the monitoring studies. Community Ecol. 2008, 9, 177–184. [Google Scholar] [CrossRef]
- Tropek, R.; Kadlec, T.; Karesova, P.; Spitzer, L.; Kocarek, P.; Malenovsky, I.; Banar, P.; Tuf, I.H.; Hejda, M.; Konvicka, M. Spontaneous succession in limestone quarries as an effective restoration tool for endangered arthropods and plants. J. Appl. Ecol. 2010, 47, 139–147. [Google Scholar] [CrossRef]
- Heneberg, P.; Bogusch, P.; Řehounek, J. Sandpits provide critical refuge for bees and wasps (Hymenoptera: Apocrita). J. Insect Conserv. 2013, 17, 473–490. [Google Scholar] [CrossRef]
- Twerd, L.; Banaszak-Cibicka, W.; Sandurska, E. What features of sand quarries affect their attractiveness for bees? Acta Oecol. 2019, 96, 56–64. [Google Scholar] [CrossRef]
- Twerd, L.; Szefer, P.; Sobieraj-Betlińska, A.; Olszewski, P. The conservation value of Aculeata communities in sand quarries changes during ecological succession. Glob. Ecol. Conserv. 2021, 28, e01693. [Google Scholar] [CrossRef]
- Twerd, L. Ecology of Wild Bees (Hymenoptera: Apoidea: Apiformes) in the Conditions Influence of the Soda Industry; Wydawnictwo Uniwersytetu Kazimierza Wielkiego: Bydgoszcz, Poland, 2020; pp. 1–258. [Google Scholar]
- Boroń, M.; Brodziak, R.; Bylka, J.; Sozański, M.M.; Urbaniak, A. The methodical concept for operation control of infiltration water intake. Ochr. Sr. 2015, 37, 29–33. [Google Scholar]
- Banaszak, J. Studies on methods of censusing the numbers of bees (Hymenoptera, Apoidea). Pol. Ecol. Stud. 1980, 6, 355–366. [Google Scholar]
- Geslin, B.; Le Féon, V.; Kuhlmann, M.; Vaissière, B.E.; Dajoz, I. The bee fauna of large parks in downtown Paris, France. Ann. Soc. Entomol. Fr. 2016, 51, 487–493. [Google Scholar] [CrossRef]
- Bossert, S. Recognition and identification of bumblebee species in the Bombus lucorum-complex (Hymenoptera, Apidae)—A review and outlook. Dtsch. Entomol. Z. 2015, 62, 19–28. [Google Scholar] [CrossRef]
- Wolf, S.; Rohde, M.; Moritz, R.F.A. The reliability of morphological traits in the differentiation of Bombus terrestris and B. lucorum (Hymenoptera: Apidae). Apidologie 2010, 41, 45–53. [Google Scholar] [CrossRef]
- Banaszak, J.; Twerd, L. Importance of thermophilous habitats for protection of wild bees (Apiformes). Community Ecol. 2018, 19, 239–247. [Google Scholar] [CrossRef]
- Banaszak-Cibicka, W.; Twerd, L.; Fliszkiewicz, M.; Giejdasz, K.; Langowska, A. City parks vs. natural areas—is it possible to preserve a natural level of bee richness and abundance in a city park? Urban Ecosyst. 2018, 21, 599–613. [Google Scholar] [CrossRef] [Green Version]
- Barkman, J.J.; Doing, H.; Segal, S. Kritische Bemerkungen und Vorschläge zur quantitativen Vegetationsanalyse. Acta Bot. Neerl. 1964, 13, 394–419. [Google Scholar] [CrossRef]
- Rutkowski, L. Klucz Do Oznaczania Roślin Naczyniowych Polski Niżowej; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2004; pp. 1–816. [Google Scholar]
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Flowering plants and pteridophytes of Poland. A checklist. In Biodiversity of Poland; Mirek, Z., Ed.; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2002; Volume 1, pp. 1–442. [Google Scholar]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von Pflanzen in Mitteleuropa. Scr. Geobot. 1992, 18, 1–258. [Google Scholar]
- QGIS 2.18.21 Essen Development Team. QGIS: A Free and Open-Source Geographic Information System; Open-Source Geospatial Foundation Project. Available online: Qgis-polska.org/pliki (accessed on 25 April 2021).
- Kujawa, K. Wpływ Struktury Zadrzewień Oraz Struktury Krajobrazu Rolniczego na Zgrupowania Ptaków Lęgowych w Zadrzewieniach; Akademia Rolnicza im. Augusta Cieszkowskiego w Poznaniu: Poznań, Poland, 2006; pp. 1–160. [Google Scholar]
- Bhattacharya, M.; Primack, R.B.; Gerwein, J. Are roads and railroads barriers to bumblebee movement in a temperate suburban conservation area? Biol. Conserv. 2003, 109, 37–45. [Google Scholar] [CrossRef]
- Farwig, N.; Bailey, D.; Bochud, E.; Herrmann, J.D.; Kindler, E.; Reusser, N.; Schüepp, C.; Schmidt-Entling, M.H. Isolation from forest reduces pollination, seed predationand insect scavenging in Swiss farmland. Landsc. Ecol. 2009, 24, 919–927. [Google Scholar] [CrossRef]
- Fischer, L.K.; Eichfeld, J.; Kowarik, I.; Buchholz, S. Disentangling urban habitat and matrix effects on wild bee species. PeerJ 2016, 4, e2729. [Google Scholar] [CrossRef] [PubMed]
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Sample; User’s Guide and Applications. 2006. Available online: Purl.oclc.org/estimates (accessed on 15 August 2021).
- Chao, A. Non-parametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘Vegan’. Community Ecology Package. R Package. 2011. Available online: Cran.r-project.org/package=vegan (accessed on 5 October 2021).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and User’s Guide to Canoco for Windows; Software for Canonical Community Ordination; Microcomputer Power: Ithaca, NY, USA, 1998. [Google Scholar]
- Novák, J.; Prach, K. Vegetation succession in basalt quarries: Pattern on a landscape scale. Appl. Veg. Sci. 2003, 6, 111–116. [Google Scholar] [CrossRef]
- Trnková, R.; Řehounková, K.; Prach, K. Spontaneous succession of vegetation on acidic bedrock in quarries in the Czech Republic. Preslia 2010, 82, 333–343. [Google Scholar]
- Brändle, M.; Durka, W.; Altmoos, M. Diversity of surface dwelling beetle assemblages in open-cast lignite mines in Central Germany. Biodiver. Conserv. 2000, 9, 1297. [Google Scholar] [CrossRef]
- Tichanek, F.; Tropek, R.J. Conservation value of post-mining headwaters: Drainage channels at a lignite spoil heap harbour threatened stream dragonflies. J. Insect Conserv. 2015, 19, 975. [Google Scholar] [CrossRef]
- Harabis, F. High diversity of odonates in post-mining areas: Meta-analysis uncovers potential pitfalls associated with the formation and management of valuable habitats. Ecol. Eng. 2016, 90, 438–446. [Google Scholar] [CrossRef]
- Hodecek, J.; Kuras, T.; Šipoš, J.; Dolný, A. Post-industrial areas as successional habitats: Long-term changes of functional diversity in beetle communities. Basic Appl. Ecol. 2015, 16, 629–640. [Google Scholar] [CrossRef]
- Steffan-Dwenter, I.; Tscharntke, T. Succession of bee communities on fallows. Ecography 2001, 24, 83–93. [Google Scholar] [CrossRef]
- Griffin, S.R.; Bruninga-Socolar, B.; Morgan, A.; Kerr, M.A.; Gibbs, J.; Winfree, R. Wild bee community change over a 26-year chronosequence of restored tallgrass prairie. Restor. Ecol. 2016, 25, 650–660. [Google Scholar] [CrossRef]
- Exeler, N.; Kratochwil, A.; Hochkirch, A. Restoration of riverine inland sand dune complexes: Implications for the conservation of wild bees. J. Appl. Ecol. 2009, 46, 1097–1105. [Google Scholar] [CrossRef]
- Tarrant, S.; Ollerton, J.; Rahman, M.L.; Tarrant, J.; McCollin, D. Grassland restoration on landfill sites in the East Midlands, United Kingdom: An evaluation of floral resources and pollinating insects. Restor. Ecol. 2013, 21, 560–568. [Google Scholar] [CrossRef]
- McKinney, A.M.; Goodell, K. Shading by invasive shrub reduces seed production and pollinator services in a native herb. Biol. Invasions. 2010, 12, 2751–2763. [Google Scholar] [CrossRef]
- Osborne, J.L.; Williams, I.H.; Corbet, S.A. Bees, pollination and habitat change in the European community. Bee World 1991, 72, 99–116. [Google Scholar] [CrossRef]
- Cane, J.H.; Tepedino, V.J. Causes and extent of declines among native North American invertebrate pollinators: Detection, evidence, and consequences. Conserv. Ecol. 2001, 5, 1. [Google Scholar] [CrossRef]
- Wulf, M.; Naaf, T. Herb layer response to broadleaf tree species with different leaf litter quality and canopy structure in temperate forests. J. Veg. Sci. 2009, 20, 517–526. [Google Scholar] [CrossRef]
- Wu, P.; Axmacher, J.C.; Song, X.; Zhang, X.; Xu, H.; Chen, C.; Yu, Z.; Liu, Y. Effects of plant diversity, vegetation composition, and habitat type on different functional trait groups of wild bees in rural Beijing. J. Insect Sci. 2018, 18, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huston, M. General hypothesis of species-diversity. Am. Nat. 1979, 113, 81–101. [Google Scholar] [CrossRef]
- Haddad, N.M.; Tilman, D.; Haarstad, J.; Ritchie, M.; Knops, J.M.H. Contrasting effects of plant richness and composition on insect communities: A field experiment. Am. Nat. 2001, 158, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, B.A.; Porter, E.E. Does herbivore diversity depend on plant diversity? The case of California butterflies. Am. Nat. 2003, 161, 40–49. [Google Scholar] [CrossRef]
- Bogusch, P.; Blažej, L.; Trýzna, M.; Heneberg, P. Forgotten role of fires in Central European forests: Critical importance of early post-fire successional stages for bees and wasps (Hymenoptera: Aculeata). Eur. J. For. Res. 2015, 134, 153–166. [Google Scholar] [CrossRef]
- Heneberg, P.; Bogusch, P.; Řezáč, M. Numerous drift sand “specialists” among bees and wasps (Hymenoptera: Aculeata) nest in wetlands that spontaneously form de novo in arable fields. Ecol. Eng. 2018, 117, 133–139. [Google Scholar] [CrossRef]
- Cane, J.H. Soils of ground-nesting bees (Hymenoptera: Apoidea): Texture, moisture, cell depth and climate. J. Kans. Entomol. Soc. 1991, 64, 406–413. [Google Scholar]
- Řehounková, K.; Prach, K. Spontaneous vegetation succession in disused gravel-sand pits: Role of local site and landscape factors. J. Veg. Sci. 2006, 17, 583–590. [Google Scholar] [CrossRef]
- Walker, L.R.; del Moral, R. (Eds.) Primary Succession and Ecosystem Rehabilitation; Cambridge University Press: Cambridge, UK, 2003; pp. 1–442. [Google Scholar]
- Prach, K.; Pysek, P.; Jarosık, V. Climate and pH as determinants of vegetation succession in Central European man-made habitats. J. Veg. Sci. 2007, 18, 701–710. [Google Scholar] [CrossRef]
- Prach, K.; Řehounková, K.; Lencová, K.; Jírová, A.; Konvalinková, P.; Mudrák, O.; Študent, V.; Vaněček, Z.; Tichy, L.; Petřík, P.; et al. Vegetation succession in restoration of disturbed sites in Central Europe: The direction of succession and species richness across 19 seres. Appl. Veg. Sci. 2014, 17, 193–200. [Google Scholar] [CrossRef]
- Potts, S.G.; Vulliamy, B.; Roberts, S.; O’Toole, C.; Dafni, A.; Ne’eman, G.; Willmer, P. Role of nesting resources in organising diverse bee communities in a Mediterranean landscape. Ecol. Entomol. 2005, 30, 78–85. [Google Scholar] [CrossRef]
- Ziaja, M.; Denisow, B.; Wrzesień, M.; Wójcik, T. Availability of food resources for pollinators in three types of lowland meadows. J. Apic. Res. 2018, 57, 467–478. [Google Scholar] [CrossRef]
- Vickruck, J.L.; Best, L.R.; Gavin, M.P.; Devries, J.H.; Galpern, P. Pothole wetlands provide reservoir habitat for native bees in prairie croplands. Biol. Conserv. 2019, 232, 43–50. [Google Scholar] [CrossRef]
- Moroń, D.; Szentgyörgyi, H.; Wantuch, M.; Celary, W.; Westphal, C.; Settele, J.; Woyciechowski, M. Diversity of wild bees in wet meadows: Implications for conservation. Wetlands 2008, 28, 975. [Google Scholar] [CrossRef]
- Stewart, R.I.A.; Andersson, G.K.S.; Brönmark, C.; Klatt, B.J.; Hansson, L.-A.; Zülsdorff, V.; Smith, H.G. Ecosystem services across the aquatic–terrestrial boundary: Linking ponds to pollination. Basic Appl. Ecol. 2017, 18, 13–20. [Google Scholar] [CrossRef]
- Twerd, L.; Banaszak-Cibicka, W. Wastelands: Their attractiveness and importance for preserving the diversity of wild bees in urban areas. J. Insect Conserv. 2019, 23, 573–588. [Google Scholar] [CrossRef]
- Banaszak-Cibicka, W.; Ratyńska, H.; Dylewski, Ł. Features of urban green space favourable for large and diverse bee populations (Hymenoptera: Apoidea: Apiformes). Urban For. Urban Green. 2016, 20, 448–452. [Google Scholar] [CrossRef]
- Ejrnæs, R.; Hansen, D.N.; Aude, E. Changing course of secondary succession in abandoned sandy fields. Biol. Conserv. 2003, 109, 343–350. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Higgs, E.S.; Hall, C.A. (Eds.) Novel Ecosystems: Intervening in the New Ecological World Order; Wiley-Blackwell: Oxford, UK, 2013; pp. 1–384. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Habitat Type | R | A | H’ | Environmental Variables | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C1 | C2 | C3 | C4 | C5 | C6 | C7 | C8 | C9 | C10 | C11 | C12 | C13 | C14 | C15 | |||||
| 1 | A | 13 | 18 | 2.40 | 13.17 | 7.54 | 15.69 | 8.13 | 12.32 | 22.50 | 0.00 | 2.55 | 0.00 | 934.00 | 4.77 | 0 | 100 | 0 | 0 |
| 2 | A | 5 | 67 | 0.31 | 8.67 | 7.32 | 15.02 | 3.50 | 8.12 | 13.37 | 1.33 | 2.14 | 0.00 | 517.00 | 5.99 | 0 | 100 | 0 | 0 |
| 3 | A | 12 | 32 | 2.13 | 9.51 | 7.21 | 14.84 | 5.51 | 9.17 | 11.08 | 2.55 | 3.32 | 0.00 | 123.00 | 12.39 | 0 | 100 | 0 | 0 |
| 4 | A | 12 | 26 | 2.16 | 9.07 | 8.51 | 17.84 | 5.18 | 7.64 | 20.03 | 5.00 | 5.04 | 0.00 | 395.00 | 11.99 | 0 | 100 | 0 | 0 |
| 5 | B | 35 | 156 | 2.83 | 2.98 | 1.59 | 2.41 | 3.10 | 3.27 | 1.91 | 5.06 | 1.80 | 20.00 | 187.00 | 14.69 | 6 | 10 | 0 | 1 |
| 6 | B | 30 | 150 | 2.65 | 7.61 | 5.34 | 6.71 | 7.76 | 7.30 | 8.99 | 9.71 | 4.62 | 10.00 | 141.00 | 16.91 | 8 | 10 | 0 | 1 |
| 7 | B | 39 | 219 | 2.73 | 4.59 | 3.00 | 5.07 | 4.44 | 5.25 | 0.55 | 6.17 | 2.45 | 20.00 | 160.00 | 15.10 | 6 | 10 | 0 | 0 |
| 8 | B | 23 | 115 | 2.27 | 5.92 | 4.83 | 4.28 | 6.36 | 5.76 | 6.82 | 5.15 | 5.17 | 20.00 | 807.00 | 2.06 | 2 | 50 | 0 | 0 |
| 9 | B | 28 | 208 | 2.34 | 4.65 | 3.28 | 3.07 | 4.89 | 4.41 | 5.31 | 7.20 | 3.45 | 20.00 | 172.00 | 8.97 | 3 | 50 | 0 | 0 |
| 10 | C | 56 | 211 | 3.10 | 5.92 | 4.60 | 1.73 | 6.41 | 5.05 | 9.21 | 8.12 | 6.17 | 414.78 | 788.00 | 7.83 | 4 | 30 | 0 | 1 |
| 11 | C | 48 | 200 | 3.27 | 5.33 | 4.00 | 1.57 | 5.88 | 4.69 | 7.68 | 4.11 | 5.39 | 526.00 | 684.00 | 8.64 | 4 | 30 | 0 | 1 |
| 12 | C | 40 | 119 | 3.13 | 6.16 | 4.65 | 1.15 | 7.33 | 6.13 | 6.31 | 7.53 | 6.13 | 670.00 | 457.00 | 6.35 | 4 | 30 | 0 | 1 |
| 13 | C | 43 | 221 | 2.81 | 7.11 | 5.14 | 1.73 | 7.84 | 6.55 | 9.37 | 9.15 | 6.85 | 685.00 | 924.00 | 7.54 | 4 | 30 | 0 | 1 |
| 14 | C | 38 | 173 | 2.94 | 7.15 | 5.63 | 2.33 | 7.77 | 6.77 | 8.73 | 9.02 | 6.30 | 621.00 | 580.00 | 7.72 | 8 | 30 | 1 | 1 |
| 15 | C | 46 | 194 | 2.99 | 6.41 | 3.21 | 2.28 | 7.01 | 6.69 | 4.51 | 9.20 | 6.16 | 610.00 | 470.00 | 6.52 | 4 | 30 | 1 | 1 |
| 16 | C | 40 | 98 | 3.31 | 4.52 | 1.78 | 3.12 | 4.77 | 4.58 | 4.21 | 7.89 | 2.82 | 600.00 | 573.00 | 7.38 | 8 | 30 | 0 | 1 |
| 17 | C | 39 | 112 | 2.97 | 3.82 | 2.12 | 2.03 | 4.07 | 4.26 | 1.82 | 5.72 | 2.52 | 597.73 | 751.00 | 7.40 | 6 | 30 | 0 | 1 |
| 18 | C | 46 | 211 | 2.94 | 5.75 | 3.35 | 3.12 | 6.06 | 6.14 | 4.19 | 5.71 | 5.55 | 639.00 | 726.00 | 1.17 | 4 | 30 | 1 | 1 |
| 19 | D | 26 | 108 | 2.67 | 5.48 | 3.83 | 3.02 | 5.79 | 5.92 | 3.93 | 5.94 | 5.96 | 842.00 | 880.00 | 3.89 | 4 | 10 | 0 | 1 |
| 20 | D | 41 | 294 | 2.59 | 6.30 | 3.89 | 2.41 | 7.17 | 7.19 | 2.85 | 7.55 | 6.90 | 808.00 | 157.00 | 12.99 | 6 | 10 | 0 | 1 |
| 21 | D | 41 | 329 | 2.60 | 6.84 | 4.55 | 5.34 | 7.01 | 6.94 | 6.31 | 9.84 | 6.79 | 661.33 | 103.00 | 11.70 | 5 | 10 | 1 | 1 |
| 22 | D | 38 | 333 | 2.64 | 4.84 | 2.78 | 1.19 | 5.30 | 4.69 | 5.68 | 6.05 | 4.46 | 508.00 | 644.00 | 3.80 | 7 | 10 | 1 | 1 |
| 23 | D | 36 | 197 | 2.38 | 4.41 | 2.95 | 1.73 | 4.91 | 3.96 | 7.24 | 6.66 | 4.11 | 804.86 | 130.00 | 13.19 | 6 | 10 | 0 | 1 |
| 24 | D | 43 | 141 | 3.31 | 4.40 | 2.72 | 0.59 | 5.09 | 4.54 | 3.71 | 8.21 | 3.55 | 790.00 | 149.00 | 12.29 | 4 | 10 | 0 | 1 |
| 25 | D | 62 | 241 | 3.52 | 5.11 | 3.83 | 0.80 | 5.67 | 4.87 | 6.32 | 9.17 | 4.67 | 1215.00 | 148.00 | 13.84 | 4 | 10 | 0 | 1 |
| 26 | D | 57 | 367 | 3.15 | 5.20 | 3.51 | 0.59 | 5.76 | 4.13 | 9.91 | 7.71 | 5.22 | 579.00 | 117.00 | 12.78 | 4 | 10 | 0 | 1 |
| 27 | D | 56 | 372 | 3.22 | 5.70 | 4.95 | 1.19 | 6.21 | 4.82 | 9.74 | 6.05 | 5.92 | 514.00 | 122.00 | 12.80 | 4 | 10 | 0 | 1 |
| Variable | df | SS | MS | F | R2 | p |
|---|---|---|---|---|---|---|
| BEES | ||||||
| Habitat type | 3 | 2.663 | 0.888 | 4.630 | 0.377 | 0.001 |
| Residuals | 23 | 4.410 | 0.192 | 0.623 | ||
| Total | 26 | 7.073 | 1 | |||
| PLANTS | ||||||
| Habitat type | 7 | 3.448 | 0.493 | 3.877 | 0.337 | 0.001 |
| Soil type (dry vs. wet) | 1 | 2.722 | 2.722 | 21.433 | 0.266 | 0.001 |
| Residuals | 32 | 4.065 | 0.127 | 0.397 | ||
| Total | 40 | 10.235 | 1 | |||
| Habitat | Total Richness | Total Abundance | |||||
|---|---|---|---|---|---|---|---|
| Estimate ± SE | z | p | Estimate ± SE | z | p | ||
| Forests vs. | clearings | 3.46 ± 1.19 | 7.04 | <0.001 | 6.85 ± 1.17 | 12.31 | <0.001 |
| ditches | 4.50 ± 1.18 | 8.90 | <0.001 | 7.60 ± 1.18 | 12.17 | <0.001 | |
| basins | 5.00 ± 1.18 | 9.62 | <0.001 | 6.29 ± 1.14 | 13.81 | <0.001 | |
| Clearings vs. | ditches | 1.30 ± 1.13 | 2.21 | 0.027 | 1.11 ± 1.21 | 0.55 | 0.583 |
| basins | 1.44 ± 1.12 | 3.21 | 0.001 | 0.92 ± 1.22 | 0.45 | 0.741 | |
| Ditches vs. | basins | 1.11 ± 1.10 | 1.05 | 0.293 | 0.83 ± 1.23 | 0.62 | 0.362 |
| Habitat/ Functional Trait | Total Richness | Total Abundance | |||||
|---|---|---|---|---|---|---|---|
| Estimate ± SE | z | p | Estimate ± SE | z | p | ||
| POLYLECTIC | |||||||
| Forests vs. | clearings | 2.39 ± 1.19 | 4.91 | <0.001 | 5.54 ± 1.17 | 10.77 | <0.001 |
| ditches | 3.19 ± 1.18 | 7.03 | <0.001 | 6.30 ± 1.18 | 10.87 | <0.001 | |
| basins | 3.74 ± 1.18 | 8.08 | <0.001 | 5.86 ± 1.14 | 13.41 | <0.001 | |
| Clearings vs. | ditches | 1.33 ± 1.12 | 2.60 | 0.009 | 1.14 ± 1.21 | 0.68 | 0.499 |
| basins | 1.56 ± 1.11 | 4.14 | <0.001 | 1.06 ± 1.21 | 0.30 | 0.764 | |
| Ditches vs. | basins | 1.17 ± 1.09 | 1.87 | 0.061 | 0.95 ± 1.22 | 0.45 | 0.748 |
| OLIGOLECTIC | |||||||
| Clearings vs. | ditches | 1.28 ± 1.47 | 0.63 | 0.527 | 1.66 ± 1.62 | 1.04 | 0.297 |
| basins | 1.33 ± 1.46 | 0.75 | 0.451 | 1.30 ± 1.59 | 0.57 | 0.568 | |
| Ditches vs. | basins | 1.04 ± 1.38 | 0.12 | 0.903 | 0.67 ± 1.51 | 0.53 | 0.669 |
| CLEPTOPARASITIC | |||||||
| Forests vs. | clearings | 11.22 ± 1.83 | 3.99 | <0.001 | 22.42 ± 1.93 | 4.73 | <0.001 |
| ditches | 15.91 ± 1.81 | 4.68 | <0.001 | 24.20 ± 1.87 | 5.08 | <0.001 | |
| basins | 15.08 ± 1.81 | 4.59 | <0.001 | 23.29 ± 1.87 | 5.02 | <0.001 | |
| Clearings vs. | ditches | 1.42 ± 1.23 | 1.66 | 0.098 | 1.08 ± 1.45 | 0.21 | 0.836 |
| basins | 1.34 ± 1.24 | 1.40 | 0.163 | 1.04 ± 1.45 | 0.10 | 0.919 | |
| Ditches vs. | basins | 0.95 ± 1.18 | −0.32 | 0.748 | 0.96 ± 1.37 | −0.12 | 0.902 |
| Habitat/ Functional Trait | Total Richness | Total Abundance | |||||
|---|---|---|---|---|---|---|---|
| Estimate ± SE | z | p | Estimate ± SE | z | p | ||
| SOIL | |||||||
| Forests vs. | clearings | 1.84 ± 1.26 | 2.66 | 0.008 | 6.39 ± 1.23 | 8.80 | <0.001 |
| ditches | 3.27 ± 1.23 | 5.67 | <0.001 | 9.81 ± 1.29 | 8.93 | <0.001 | |
| basins | 3.84 ± 1.23 | 6.54 | <0.001 | 4.30 ± 1.16 | 10.02 | <0.001 | |
| Clearings vs. | ditches | 1.78 ± 1.18 | 3.48 | <0.001 | 1.54 ± 1.35 | 1.43 | 0.154 |
| basins | 2.09 ± 1.17 | 4.60 | <0.001 | 0.67 ± 1.33 | 1.62 | 0.127 | |
| Ditches vs. | basins | 1.17 ± 1.13 | 1.30 | 0.193 | 0.42 ± 1.35 | 2.88 | 0.005 |
| HIVE | |||||||
| Forests vs. | clearings | 3.15 ± 1.33 | 3.98 | <0.001 | 13.54 ± 2.66 | 2.67 | 0.008 |
| ditches | 2.19 ± 1.33 | 2.78 | 0.005 | 5.00 ± 3.27 | 1.36 | 0.174 | |
| basins | 3.70 ± 1.31 | 4.81 | <0.001 | 13.56 ± 3.24 | 2.22 | 0.027 | |
| Clearings vs. | ditches | 0.70 ± 1.19 | −2.08 | 0.038 | 0.37 ± 2.60 | −1.04 | 0.297 |
| basins | 1.18 ± 1.17 | 1.04 | 0.300 | 1.00 ± 2.62 | 0 | 0.999 | |
| Ditches vs. | basins | 1.69 ± 1.16 | 3.59 | <0.001 | 2.71 ± 3.26 | 0.84 | 0.399 |
| CAVITY | |||||||
| Forests vs. | clearings | 20.77 ± 2.75 | 3.00 | 0.003 | 31.97 ± 2.87 | 3.29 | 0.001 |
| ditches | 30.20 ± 2.72 | 3.41 | 0.001 | 112.25 ± 2.82 | 4.55 | <0.001 | |
| basins | 19.42 ± 2.73 | 2.96 | 0.003 | 26.91 ± 2.84 | 3.15 | 0.002 | |
| Clearings vs. | ditches | 1.45 ± 1.28 | 1.50 | 0.133 | 3.51 ± 1.56 | 2.83 | 0.005 |
| basins | 0.94 ± 1.30 | −0.26 | 0.798 | 0.84 ± 1.56 | −0.39 | 0.662 | |
| Ditches vs. | basins | 0.64 ± 1.24 | −2.09 | 0.037 | 0.24 ± 1.49 | −3.58 | <0.001 |
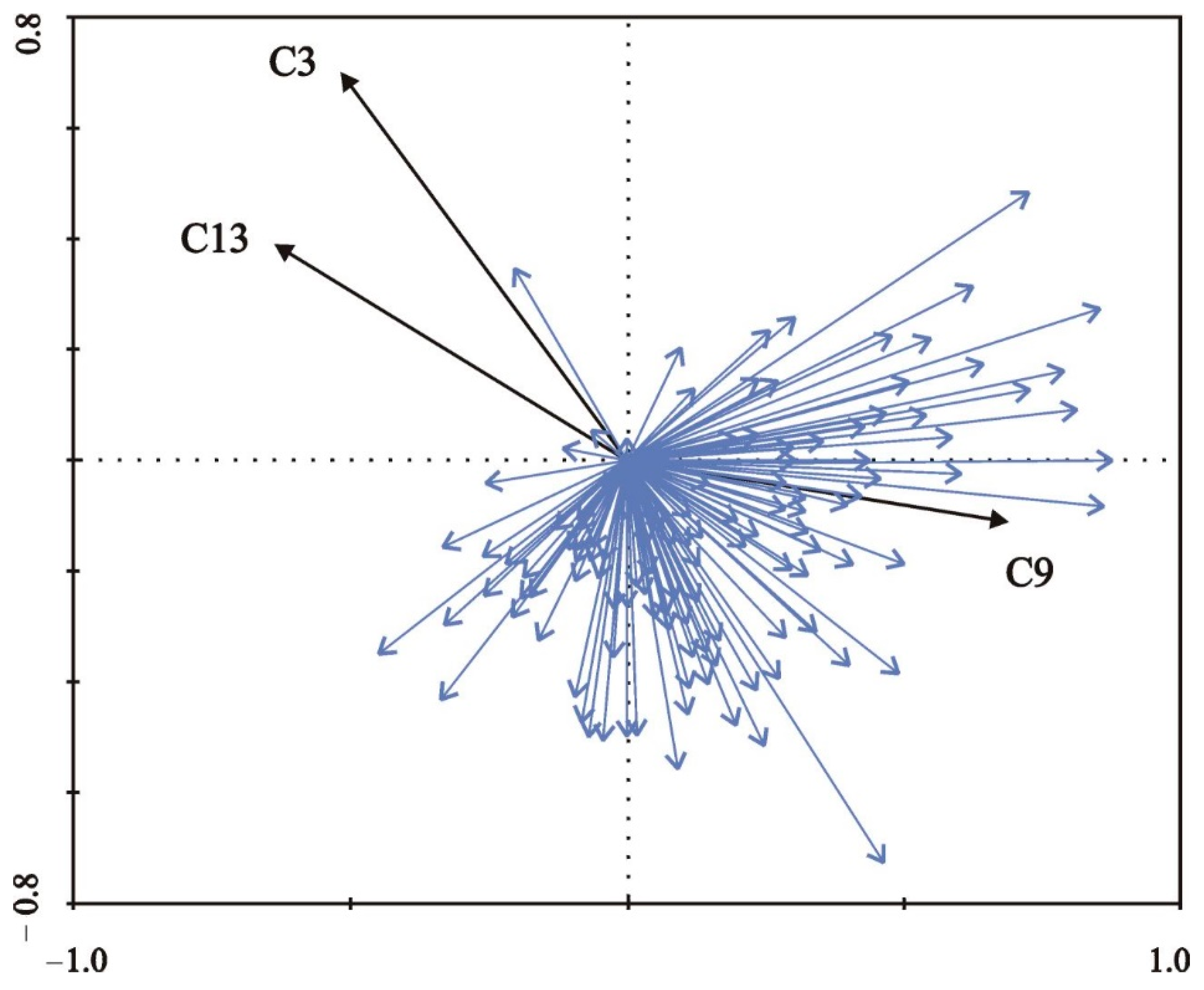
| Variable | RDA | |||
|---|---|---|---|---|
| p | Variance | % of Explained Variance | VIF | |
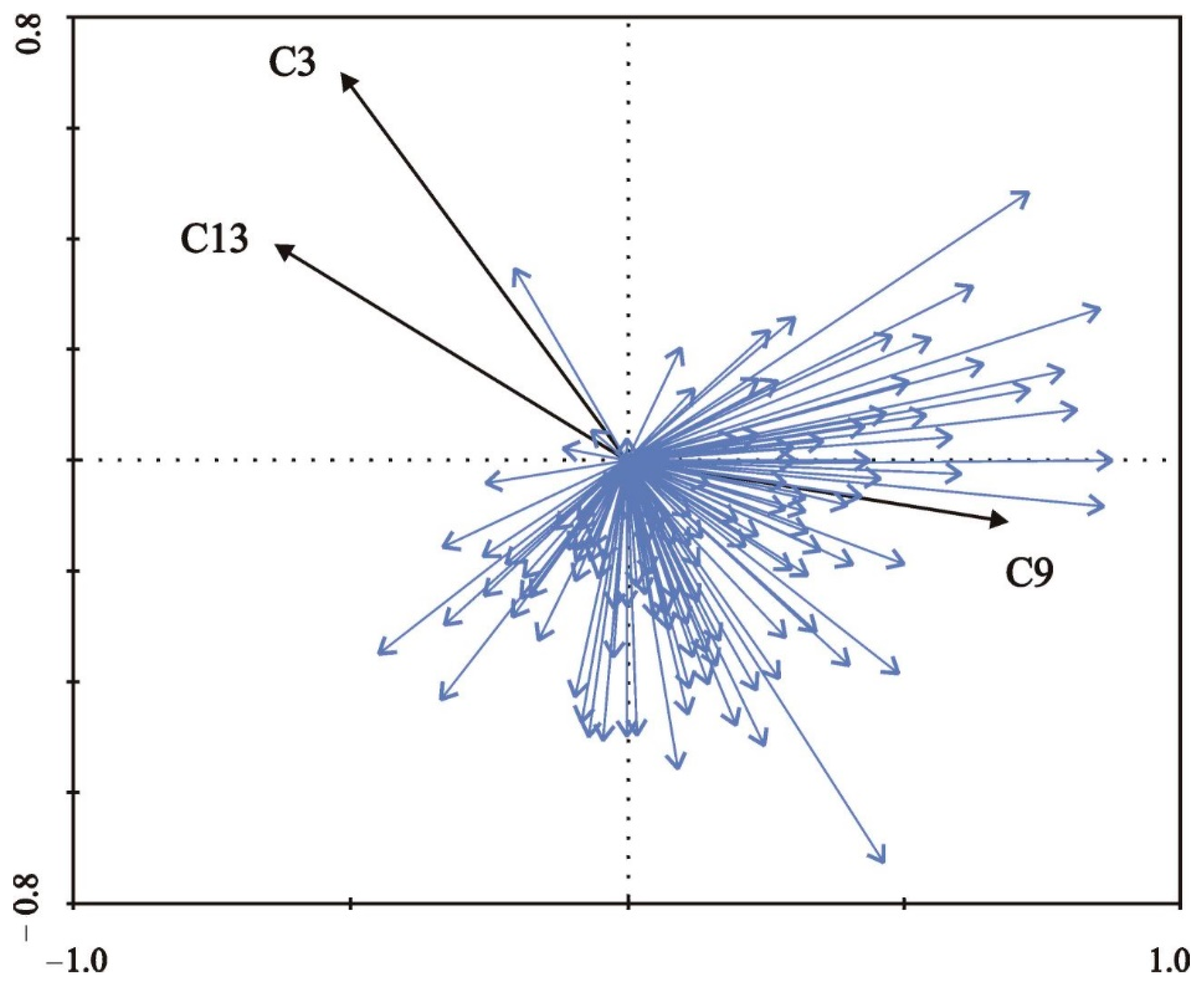
| C3 = mean cover by woody species | 0.001 | 0.12 | 3.55 | 3.884 |
| C9 = mean radius of a belt of predominantly open habitats around sampling sites | 0.001 | 0.10 | 2.80 | 3.427 |
| C13 = mean degree of shading at daytime | 0.022 | 0.05 | 1.87 | 7.923 |
| C14 = presence of water | 0.105 | 0.05 | 1.44 | 1.494 |
| C10 = site area (m2) | 0.248 | 0.04 | 1.19 | 4.013 |
| C11 = coefficient of border development | 0.149 | 0.04 | 1.35 | 5.918 |
| C12 = habitat barriers | 0.320 | 0.03 | 1.13 | 3.884 |
| C15 = presence of scarps | 0.350 | 0.03 | 1.13 | 6.226 |
| C7 = mean cover by plant species typical of dry soils | 0.738 | 0.03 | 0.75 | 2.801 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Twerd, L.; Sobieraj-Betlińska, A.; Kilińska, B.; Waldon-Rudzionek, B.; Hoffmann, R.; Banaszak, J. Unexpectedly, Creation of Temporary Water Bodies Has Increased the Availability of Food and Nesting Sites for Bees (Apiformes). Forests 2022, 13, 1410. https://doi.org/10.3390/f13091410
Twerd L, Sobieraj-Betlińska A, Kilińska B, Waldon-Rudzionek B, Hoffmann R, Banaszak J. Unexpectedly, Creation of Temporary Water Bodies Has Increased the Availability of Food and Nesting Sites for Bees (Apiformes). Forests. 2022; 13(9):1410. https://doi.org/10.3390/f13091410
Chicago/Turabian StyleTwerd, Lucyna, Anna Sobieraj-Betlińska, Barbara Kilińska, Barbara Waldon-Rudzionek, Renata Hoffmann, and Józef Banaszak. 2022. "Unexpectedly, Creation of Temporary Water Bodies Has Increased the Availability of Food and Nesting Sites for Bees (Apiformes)" Forests 13, no. 9: 1410. https://doi.org/10.3390/f13091410
APA StyleTwerd, L., Sobieraj-Betlińska, A., Kilińska, B., Waldon-Rudzionek, B., Hoffmann, R., & Banaszak, J. (2022). Unexpectedly, Creation of Temporary Water Bodies Has Increased the Availability of Food and Nesting Sites for Bees (Apiformes). Forests, 13(9), 1410. https://doi.org/10.3390/f13091410