Stomatal Limitation Is Able to Modulate Leaf Coloration Onset of Temperate Deciduous Tree




Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.1.1. Study Site and Climate Chambers
2.1.2. Tree Material
2.1.3. Treatments and Environment Controls
2.2. Phenology Observation
2.3. Leaf Gas Exchange Parameters and CO2 Response Curve Measurements
2.4. Leaf Chlorophyll Content and Water Content Measurements
2.5. Statistical Analysis
3. Results
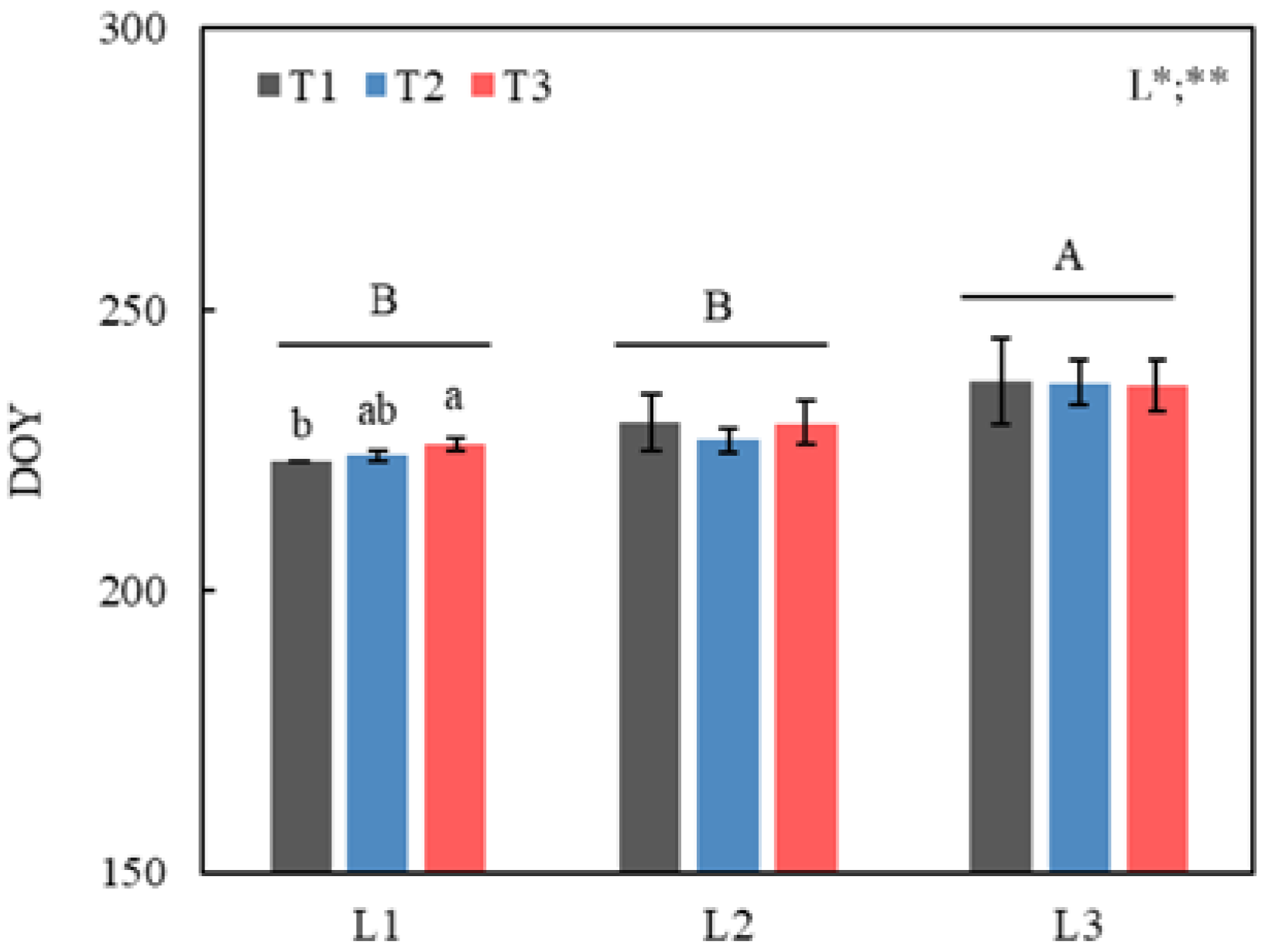
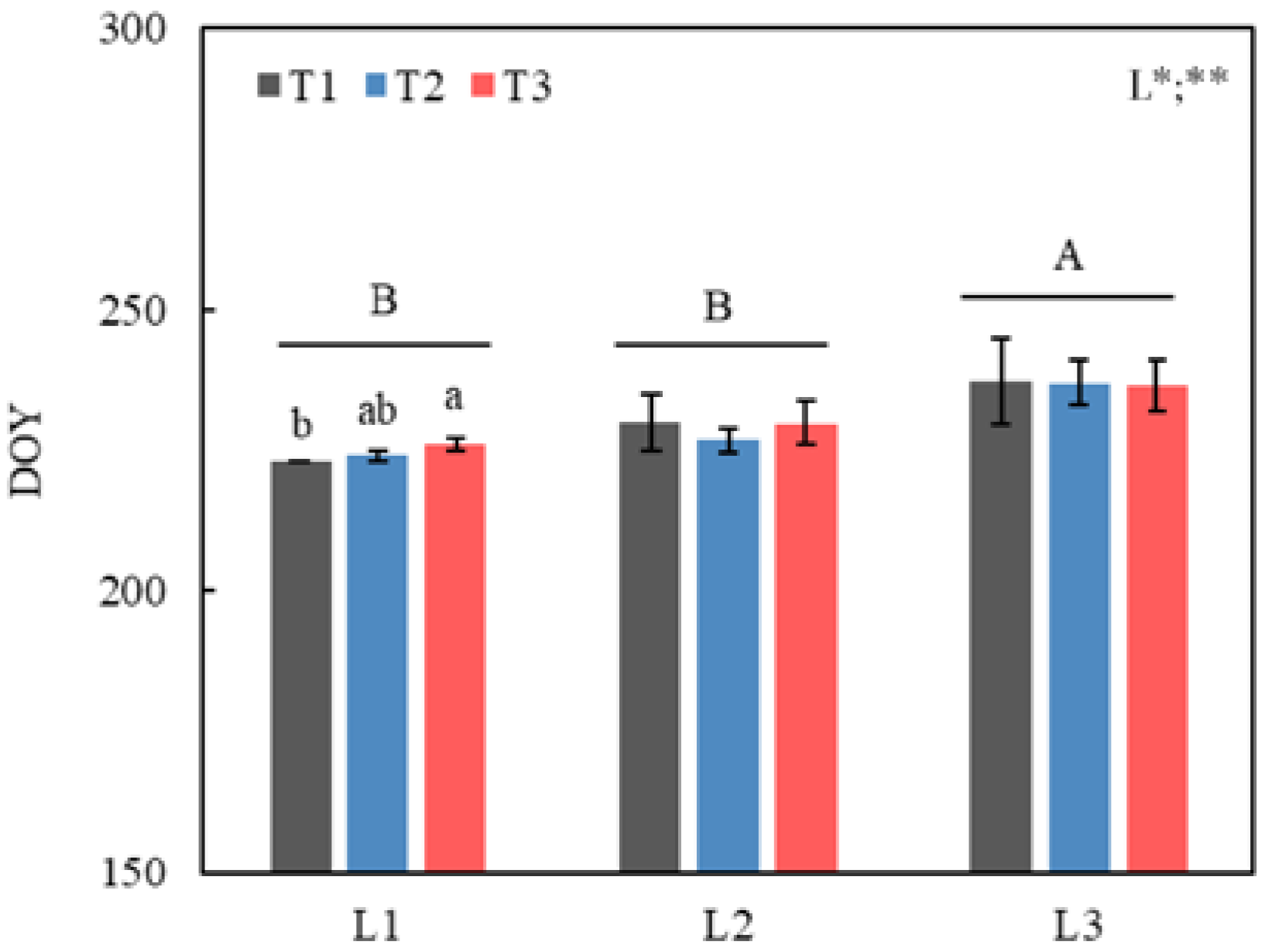
3.1. LCO under Different Temperature and Photoperiod Treatments
3.2. Leaf Gas Exchange Parameters under Different Temperature and Photoperiod Treatments
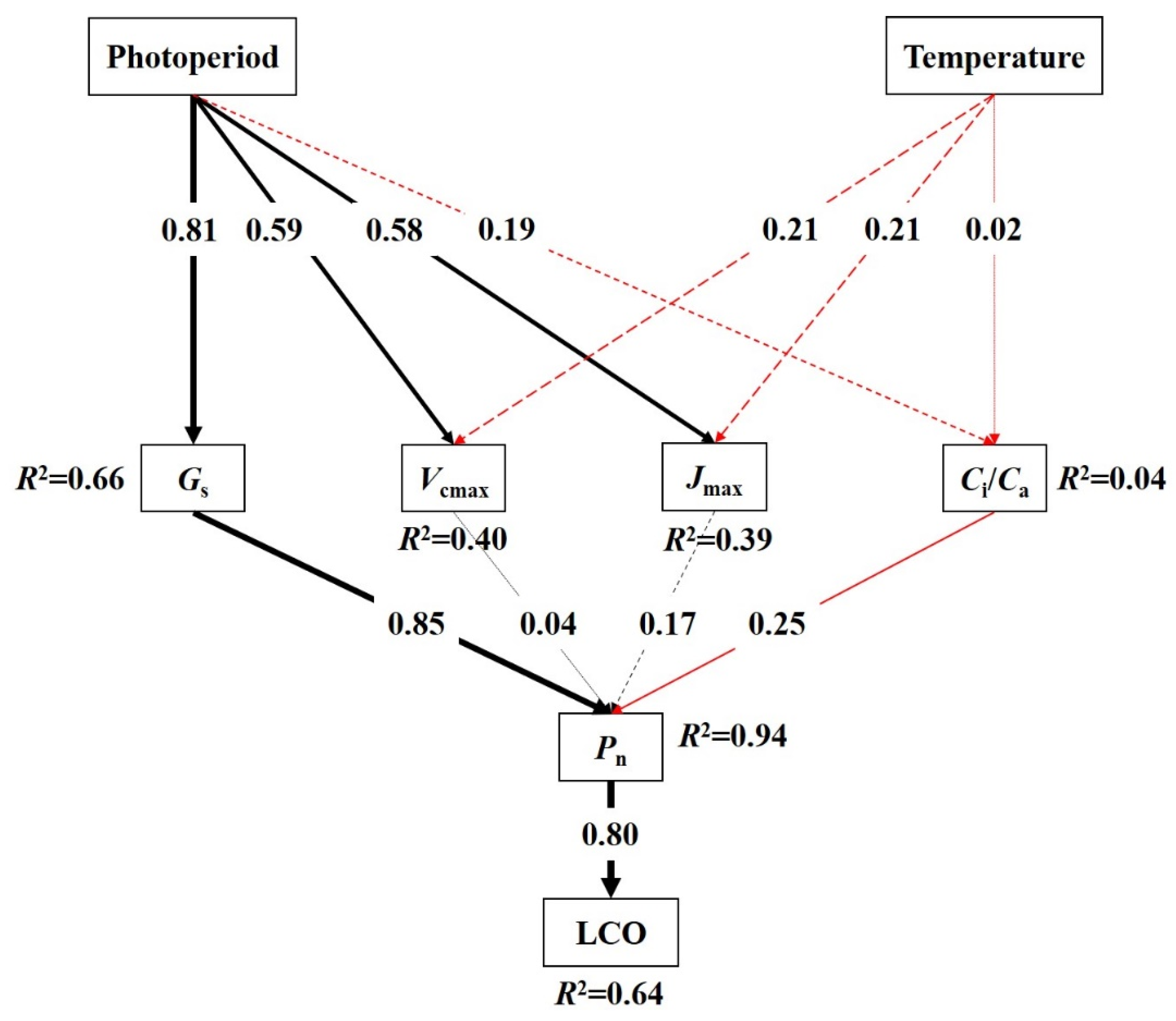
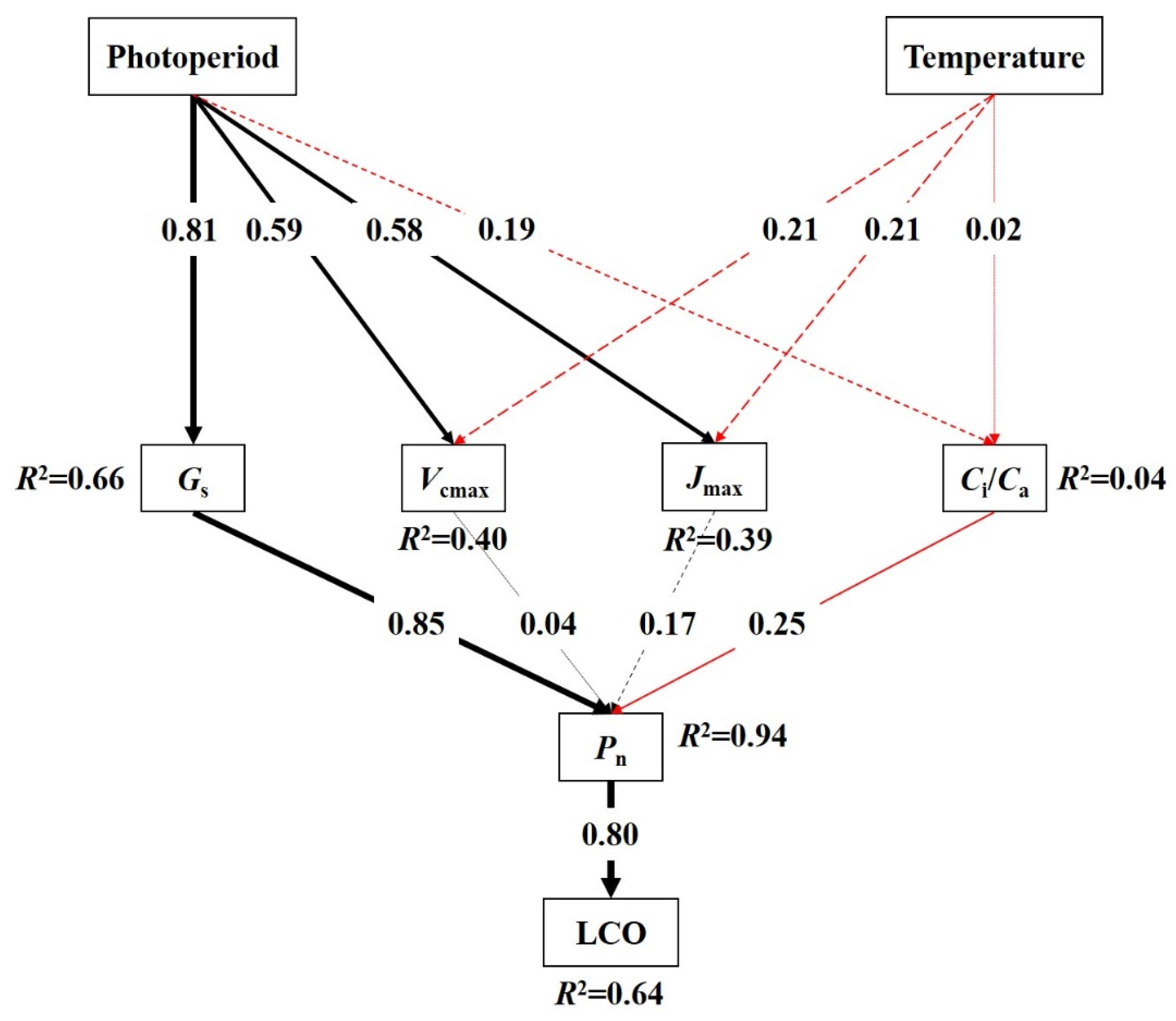
3.3. Regulation Mechanisms of Temperature and Photoperiod on LCO
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ge, Q.; Wang, H.; Rutishauser, T.; Dai, J. Phenological response to climate change in China: A meta-analysis. Glob. Chang. Biol. 2015, 21, 265–274. [Google Scholar] [CrossRef]
- Piao, S.; Liu, Q.; Chen, A.; Janssens, I.A.; Fu, Y.; Dai, J.; Liu, L.; Lian, X.; Shen, M.; Zhu, X. Plant phenology and global climate change: Current progresses and challenges. Glob. Chang. Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Q.; Zhou, G.S.; Zhou, M.Z.; Lü, X.M.; Zhou, L.; Ji, Y.H. Photosynthetically physiological mechanism of Stipa krylovii withered and yellow phenology response to precipitation under the background of warming. J. Appl. Ecol. 2021, 32, 845–852. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, Y.; Jia, Q.; Zhou, G. Increasing temperature shortened the carbon uptake period and decreased the cumulative net ecosystem productivity in a maize cropland in Northeast China. Field Crop Res. 2021, 267, 108150. [Google Scholar] [CrossRef]
- Steinaker, D.F.; Wilson, S.D. Phenology of fine roots and leaves in forest and grassland. J. Ecol. 2008, 96, 1222–1229. [Google Scholar] [CrossRef]
- Huang, L.; Koubek, T.; Weiser, M.; Herben, T.; Cornelissen, H. Environmental drivers and phylogenetic constraints of growth phenologies across a large set of herbaceous species. J. Ecol. 2018, 106, 1621–1633. [Google Scholar] [CrossRef]
- Berra, E.F.; Gaulton, R. Remote sensing of temperate and boreal forest phenology: A review of progress, challenges and opportunities in the intercomparison of in-situ and satellite phenological metrics. Forest Ecol. Manag. 2021, 480, 118663. [Google Scholar] [CrossRef]
- Menzel, A. Trends in phenological phases in Europe between 1951 and 1996. Int. J. Biometeorol. 2000, 44, 76–81. [Google Scholar] [CrossRef]
- Cleland, E.E.; Allen, J.M.; Crimmins, T.M.; Dunne, J.A.; Pau, S.; Travers, S.E.; Zavaleta, E.S.; Wolkovich, E.M. Phenological tracking enables positive species responses to climate change. Ecology 2012, 93, 1765–1771. [Google Scholar] [CrossRef]
- Svystun, T.; Lundströmer, J.; Berlin, M.; Westin, J.; Jönsson, A.M. Model analysis of temperature impact on the Norway spruce provenance specific bud burst and associated risk of frost damage. Forest Ecol. Manag. 2021, 493, 119252. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, F.; Zheng, J.; Lin, J.; Hänninen, H.; Wu, J. Chilling accumulation and photoperiod regulate rest break and bud burst in five subtropical tree species. Forest Ecol. Manag. 2021, 485, 118813. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, X.; Yu, Y.; Donnelly, A. Detecting spatiotemporal changes of peak foliage coloration in deciduous and mixedforests across the Central and Eastern United States. Environ. Res. Lett. 2017, 12, 024013. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Peng, C.; Wang, M.; Luo, Y.; Li, M.; Zhang, K.; Zhang, D.; Zhu, Q. Dynamics of vegetation autumn phenology and its response to multiple environmental factors from 1982 to 2012 on Qinghai-Tibetan Plateau in China. Sci. Total Environ. 2018, 637–638, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhu, Q.; Peng, C.; Zhang, J.; Wang, M.; Zhang, J.; Ding, J.; Zhou, X. Change in autumn vegetation phenology and the climate controls from 1982 to 2012 on the Qinghai–Tibet Plateau. Front. Plant Sci. 2020, 10, 1677. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Dai, J.; Ge, Q. Responses of autumn phenology to climate change and the correlations of plant hormone regulation. Sci. Rep. 2020, 10, 9039. [Google Scholar] [CrossRef]
- Jeong, S.J.; Ho, C.H.; Gim, H.J.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Chang. Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- Wu, C.; Chen, J.M.; Black, T.A.; Price, D.T.; Kurz, W.A.; Desai, A.R.; Gonsamo, A.; Jassal, R.S.; Gough, C.M.; Bohrer, G.; et al. Interannual variability of net ecosystem productivity in forests is explained by carbon flux phenology in autumn. Global Ecol. Biogeogr. 2013, 22, 994–1006. [Google Scholar] [CrossRef]
- Lee, S.; Jeong, S.; Park, C.E.; Kim, J. A Simple method of predicting autumn leaf coloring date using machine learning with spring leaf unfolding date. Asia-Pac. J. Atmos. Sci. 2022, 58, 219–226. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Lang, W.; Chen, X.; Qian, S.; Liu, G.; Piao, S. A new process-based model for predicting autumn phenology: How is leaf senescence controlled by photoperiod and temperature coupling? Agric. For. Meteorol. 2019, 268, 124–135. [Google Scholar] [CrossRef]
- Chu, X.; Man, R.; Zhang, H.; Yuan, W.; Tao, J.; Dang, Q.L. Does climate warming favour early season species? Front. Plant Sci. 2021, 12, 765351. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Fu, Y.H.; Geng, X.; Hao, Z.; Tang, J.; Zhang, X.; Xu, Z.; Hao, F. Influences of shifted vegetation phenology on runoff across a hydroclimatic gradient. Front. Plant Sci. 2022, 12, 802664. [Google Scholar] [CrossRef] [PubMed]
- Keskitalo, J.; Bergquist, G.; Gardestrom, P.; Jansson, S. A cellular timetable of autumn senescence. Plant Physiol. 2005, 139, 1635–1648. [Google Scholar] [CrossRef] [Green Version]
- Saxe, H.; Cannell, M.G.R.; Johnsen, Ø.; Ryan, M.G.; Vourlitis, G. Tree and forest functioning in response to global warming. New Phytol. 2001, 149, 369–399. [Google Scholar] [CrossRef] [PubMed]
- Estiarte, M.; Peñuelas, J. Alteration of the phenology of leaf senescence and fall in winter deciduous species by climate change: Effects on nutrient proficiency. Glob. Chang. Biol. 2015, 21, 1005–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bucher, S.F.; Römermann, C.; Bonser, S. The timing of leaf senescence relates to flowering phenology and functional traits in 17 herbaceous species along elevational gradients. J. Ecol. 2021, 109, 1537–1548. [Google Scholar] [CrossRef]
- Zani, D.; Crowther, T.W.; Mo, L.D.; Renner, S.S.; Zohner, C.M. Increased growing-season productivity drives earlier autumn leaf senescence in temperate trees. Science 2020, 370, 1066–1071. [Google Scholar] [CrossRef]
- Xia, H.; Wang, B. Complete chloroplast genome sequence of the Mongolian oak, Quercus mongolica (Fagaceae). Mitochondrial DNA Part B 2019, 4, 1089–1090. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.X.; Zhou, G.S.; Lü, X.M.; Wang, S.Q.; Zhang, S.Y. Interactive effects of different warming and changing photoperiod on spring phenology of Quercu smongolicus seedlings. Acta Ecol. Sin. 2021, 41, 2816–2825. [Google Scholar] [CrossRef]
- Wan, M.W.; Liu, X.Z. Phenological Observation Methods in China; Science Press: Beijing, China, 1979. [Google Scholar]
- Davis, J.E.; Arkebauer, T.J.; Norman, J.M.; Brandle, J.R. Rapid field measurement of the assimilation rate versus internal CO2 concentration relationship in green ash (Fraxinus pennsylvanica Marsh.): The influence of light intensity. Tree Physiol. 1987, 3, 387–392. [Google Scholar] [CrossRef]
- McDermitt, D.; Norman, J.; Davis, J.; Ball, T.; Arkebauer, T.; Welles, J.; Roerner, S. CO2 response curves can be measured with a field-portable closed-loop photosynthesis system. Ann. Sc. For. 1989, 46, 416s–420s. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Zhou, G.; He, Q.; Zhou, L.; Ji, Y.; Lv, X. Capability of leaf water content and its threshold values in reflection of soil–plant water status in maize during prolonged drought. Ecol. Indic. 2021, 124, 107395. [Google Scholar] [CrossRef]
- Garnier, E.; Laurent, G. Leaf anatomy, specific mass and water content in congeneric annual and perennial grass species. New Phytol. 1994, 128, 725–736. [Google Scholar] [CrossRef]
- Fracheboud, Y.; Luquez, V.; Bjorken, L.; Sjodin, A.; Tuominen, H.; Jansson, S. The control of autumn senescence in European aspen. Plant Physiol. 2009, 149, 1982–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, S.; Dang, Q.L.; Man, R.; Tedla, B. Photoperiod, [CO2] and soil moisture interactively affect phenology in trembling aspen: Implications to climate change-induced migration. Environ. Exp. Bot. 2020, 180, 104269. [Google Scholar] [CrossRef]
- Estrella, N.; Menzel, A. Responses of leaf colouring in four deciduous tree species to climate and weather in Germany. Clim. Res. 2006, 32, 253–267. [Google Scholar] [CrossRef] [Green Version]
- Delpierre, N.; Dufrêne, E.; Soudani, K.; Ulrich, E.; Cecchini, S.; Boé, J.; François, C. Modelling interannual and spatial variability of leaf senescence for three deciduous tree species in France. Agric. For. Meteorol. 2009, 149, 938–948. [Google Scholar] [CrossRef]
- Singh, R.K.; Bhalerao, R.P.; Eriksson, M.E. Growing in time: Exploring the molecular mechanisms of tree growth. Tree Physiol. 2021, 41, 657–678. [Google Scholar] [CrossRef]
- Meng, L.; Zhou, Y.; Gu, L.; Richardson, A.D.; Penuelas, J.; Fu, Y.; Wang, Y.; Asrar, G.R.; De Boeck, H.J.; Mao, J.; et al. Photoperiod decelerates the advance of spring phenology of six deciduous tree species under climate warming. Glob. Chang. Biol. 2021, 27, 2914–2927. [Google Scholar] [CrossRef]
- Doi, H.; Takahashi, M. Latitudinal patterns in the phenological responses of leaf colouring and leaf fall to climate change in Japan. Global Ecol. Biogeogr. 2008, 17, 556–561. [Google Scholar] [CrossRef]
- Fu, Y.H.; Piao, S.; Delpierre, N.; Hao, F.; Hanninen, H.; Liu, Y.; Sun, W.; Janssens, I.A.; Campioli, M. Larger temperature response of autumn leaf senescence than spring leaf-out phenology. Glob. Chang. Biol. 2018, 24, 2159–2168. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Huang, Y.; Wei, Y.; Zhang, W.; Li, T.; Zhang, Q. Inner Mongolian grassland plant phenological changes and their climatic drivers. Sci. Total Environ. 2019, 683, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Ma, X.; Dou, X.; Zhu, J.; Zhao, C. Impacts of climate change on vegetation phenology and net primary productivity in arid Central Asia. Sci. Total Environ. 2021, 796, 149055. [Google Scholar] [CrossRef] [PubMed]
- Gunderson, C.A.; Edwards, N.T.; Walker, A.V.; O’Hara, K.H.; Campion, C.M.; Hanson, P.J. Forest phenology and a warmer climate-growing season extension in relation to climatic provenance. Glob. Chang. Biol. 2012, 18, 2008–2025. [Google Scholar] [CrossRef]
- Morin, X.; Roy, J.; Sonie, L.; Chuine, I. Changes in leaf phenology of three European oak species in response to experimental climate change. New Phytol. 2010, 186, 900–910. [Google Scholar] [CrossRef]
- Pudas, E.; Tolvanen, A.; Poikolainen, J.; Sukuvaara, T.; Kubin, E. Timing of plant phenophases in Finnish Lapland in 1997–2006. Boreal Environ. Res. 2008, 13, 31–43. [Google Scholar]
- Tanino, K.K.; Kalcsits, L.; Silim, S.; Kendall, E.; Gray, G.R. Temperature-driven plasticity in growth cessation and dormancy development in deciduous woody plants: A working hypothesis suggesting how molecular and cellular function is affected by temperature during dormancy induction. Plant Mol. Biol. 2010, 73, 49–65. [Google Scholar] [CrossRef]
- Barr, A.G.; Black, T.A.; Hogg, E.H.; Kljun, N.; Morgenstern, K.; Nesic, Z. Inter-annual variability in the leaf area index of a boreal aspen-hazelnut forest in relation to net ecosystem production. Agric. For. Meteorol. 2004, 126, 237–255. [Google Scholar] [CrossRef]
- Frechette, E.; Chang, C.Y.; Ensminger, I. Variation in the phenology of photosynthesis among eastern white pine provenances in response to warming. Glob. Chang. Biol. 2020, 26, 5217–5234. [Google Scholar] [CrossRef]
- Kong, D.; Zhang, Y.; Wang, D.; Chen, J.; Gu, X. Photoperiod explains the asynchronization between vegetation carbon phenology and vegetation greenness phenology. J. Geophys. Res. Biogeosci. 2020, 125, 1–15. [Google Scholar] [CrossRef]
- Bauerle, W.L.; Oren, R.; Way, D.A.; Qian, S.S.; Stoy, P.C.; Thornton, P.E.; Bowden, J.D.; Hoffman, F.M.; Reynolds, R.F. Photoperiodic regulation of the seasonal pattern of photosynthetic capacity and the implications for carbon cycling. Proc. Natl. Acad. Sci. USA 2012, 109, 8612–8617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoshita, T.; Kume, A.; Hanba, Y.T. Seasonal variations in photosynthetic functions of the urban landscape tree species Gingko biloba: Photoperiod is a key trait. Trees 2020, 35, 273–285. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Temperature (℃) | Relative Humidity (%) | Precipitation (mm) | Irrigation Amount (mL) |
|---|---|---|---|---|
| April | 5.65 | 50.94 | 19.51 | 55 |
| May | 14.02 | 51.41 | 37.64 | 107 |
| June | 19.96 | 65.16 | 87.04 | 247 |
| July | 22.33 | 76.87 | 155.24 | 440 |
| August | 20.28 | 78.68 | 113.22 | 321 |
| September | 13.61 | 69.75 | 54.45 | 154 |
| October | 4.45 | 61.76 | 20.16 | 57 |
| Month | L | T | L × T | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| df | F | P | df | F | P | df | F | P | ||
| Pn | May | 2 | 0.49 | 0.62 | 2 | 4.24 | <0.05 | 4 | 0.33 | 0.86 |
| June | 2 | 2.52 | 0.14 | 2 | 2.58 | 0.13 | 4 | 1.54 | 0.27 | |
| July | 2 | 3.86 | <0.05 | 2 | 5.23 | <0.05 | 4 | 2.44 | 0.10 | |
| August | 2 | 33.39 | <0.01 | 2 | 1.26 | 0.31 | 4 | 1.51 | 0.25 | |
| LCO | 2 | 4.19 | <0.05 | 2 | 1.84 | 0.20 | 4 | 4.06 | <0.05 | |
| Gs | May | 2 | 1.07 | 0.36 | 2 | 8.45 | <0.01 | 4 | 1.04 | 0.42 |
| June | 2 | 7.74 | <0.05 | 2 | 9.23 | <0.01 | 4 | 4.79 | <0.05 | |
| July | 2 | 4.96 | <0.05 | 2 | 2.53 | 0.12 | 4 | 1.85 | 0.18 | |
| August | 2 | 23.86 | <0.01 | 2 | 0.11 | 0.90 | 4 | 1.30 | 0.32 | |
| LCO | 2 | 4.13 | <0.05 | 2 | 1.24 | 0.32 | 4 | 2.62 | 0.09 | |
| Ci | May | 2 | 9.53 | <0.01 | 2 | 3.36 | 0.06 | 4 | 0.45 | 0.77 |
| June | 2 | 0.03 | 0.97 | 2 | 1.55 | 0.26 | 4 | 0.34 | 0.85 | |
| July | 2 | 2.97 | 0.08 | 2 | 1.71 | 0.22 | 4 | 3.02 | 0.05 | |
| August | 2 | 0.84 | 0.45 | 2 | 0.56 | 0.58 | 4 | 1.39 | 0.29 | |
| LCO | 2 | 0.38 | 0.69 | 2 | 0.06 | 0.94 | 4 | 0.16 | 0.95 | |
| Ci/Ca | May | 2 | 9.58 | <0.01 | 2 | 3.45 | 0.06 | 4 | 0.45 | 0.77 |
| June | 2 | 0.05 | 0.95 | 2 | 1.67 | 0.24 | 4 | 0.37 | 0.82 | |
| July | 2 | 2.94 | 0.09 | 2 | 1.69 | 0.22 | 4 | 3.02 | 0.06 | |
| August | 2 | 0.82 | 0.46 | 2 | 0.50 | 0.62 | 4 | 1.40 | 0.28 | |
| LCO | 2 | 0.40 | 0.68 | 2 | 0.05 | 0.95 | 4 | 0.15 | 0.96 | |
| Tr | May | 2 | 4.84 | <0.05 | 2 | 13.18 | <0.01 | 4 | 2.30 | 0.10 |
| June | 2 | 22.35 | <0.01 | 2 | 15.48 | <0.01 | 4 | 7.49 | <0.01 | |
| July | 2 | 11.66 | <0.01 | 2 | 4.37 | <0.05 | 4 | 3.74 | <0.05 | |
| August | 2 | 18.88 | <0.01 | 2 | 0.07 | 0.93 | 4 | 1.48 | 0.26 | |
| LCO | 2 | 5.31 | <0.05 | 2 | 1.01 | 0.39 | 4 | 1.94 | 0.17 | |
| L | T | L × T | |||||||
|---|---|---|---|---|---|---|---|---|---|
| df | F | P | df | F | P | df | F | P | |
| Vcmax | 2 | 5.15 | <0.05 | 2 | 0.82 | 0.46 | 4 | 0.98 | 0.45 |
| Jmax | 2 | 5.30 | <0.05 | 2 | 0.93 | 0.42 | 4 | 0.03 | 1.00 |
| Chl | 2 | 1.26 | 0.31 | 2 | 0.37 | 0.70 | 4 | 0.35 | 0.84 |
| LWC | 2 | 2.74 | 0.09 | 2 | 1.97 | 0.17 | 4 | 0.37 | 0.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.; Zhou, G.; Lv, X.; He, Q.; Zhou, M. Stomatal Limitation Is Able to Modulate Leaf Coloration Onset of Temperate Deciduous Tree. Forests 2022, 13, 1099. https://doi.org/10.3390/f13071099
Yu H, Zhou G, Lv X, He Q, Zhou M. Stomatal Limitation Is Able to Modulate Leaf Coloration Onset of Temperate Deciduous Tree. Forests. 2022; 13(7):1099. https://doi.org/10.3390/f13071099
Chicago/Turabian StyleYu, Hongying, Guangsheng Zhou, Xiaomin Lv, Qijin He, and Mengzi Zhou. 2022. "Stomatal Limitation Is Able to Modulate Leaf Coloration Onset of Temperate Deciduous Tree" Forests 13, no. 7: 1099. https://doi.org/10.3390/f13071099
APA StyleYu, H., Zhou, G., Lv, X., He, Q., & Zhou, M. (2022). Stomatal Limitation Is Able to Modulate Leaf Coloration Onset of Temperate Deciduous Tree. Forests, 13(7), 1099. https://doi.org/10.3390/f13071099