
Molecular Mechanisms Regulating the Columnar Tree Architecture in Apple



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
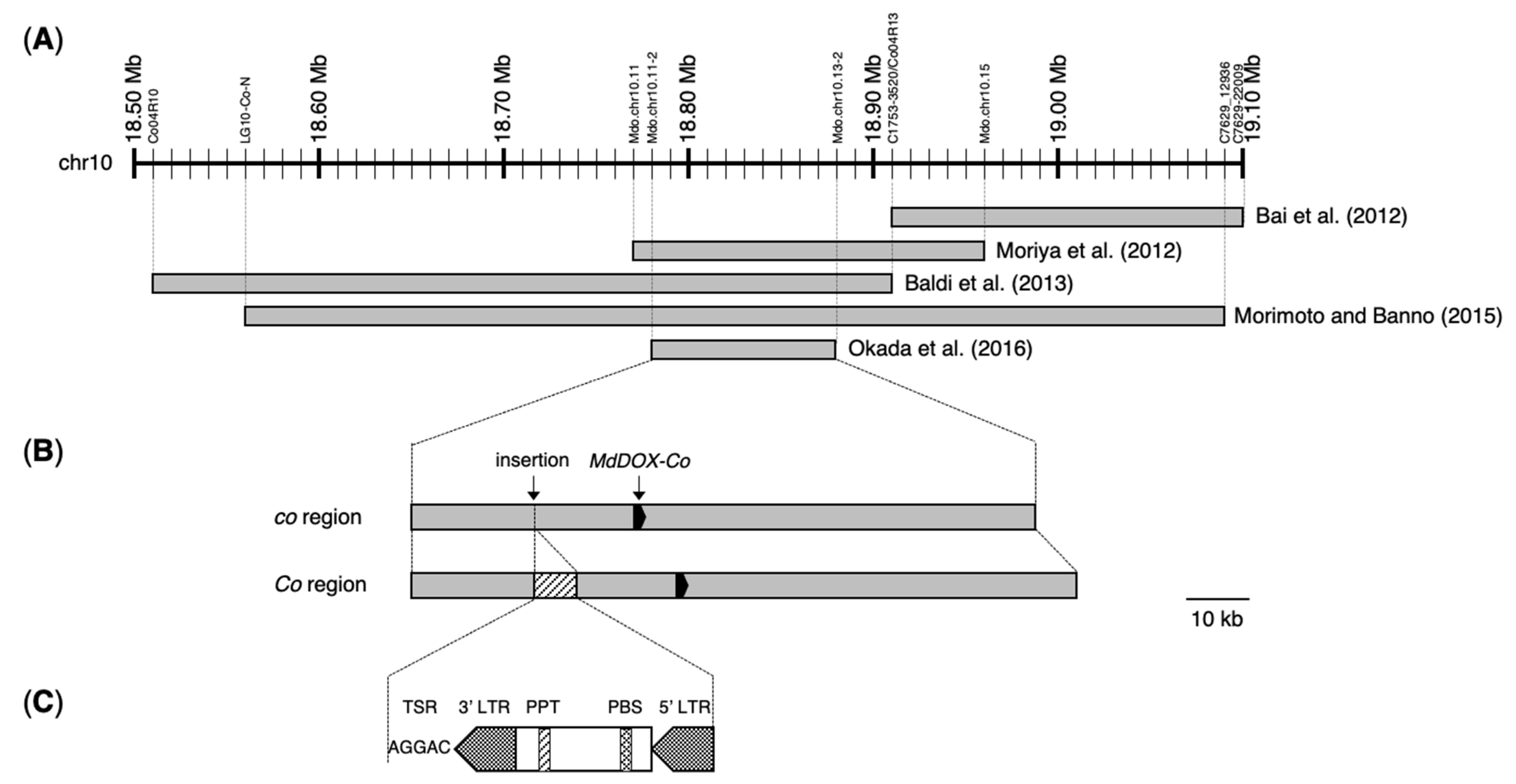
2. Fine Mapping of the Co Locus
3. Identification of Mutation in ‘McIntosh Wijcik’
4. Exploration of Co Candidate Genes
5. Expression Analysis of MdDOX-Co
6. Phenotypes of Transgenic Plants Overexpressing MdDOX-Co
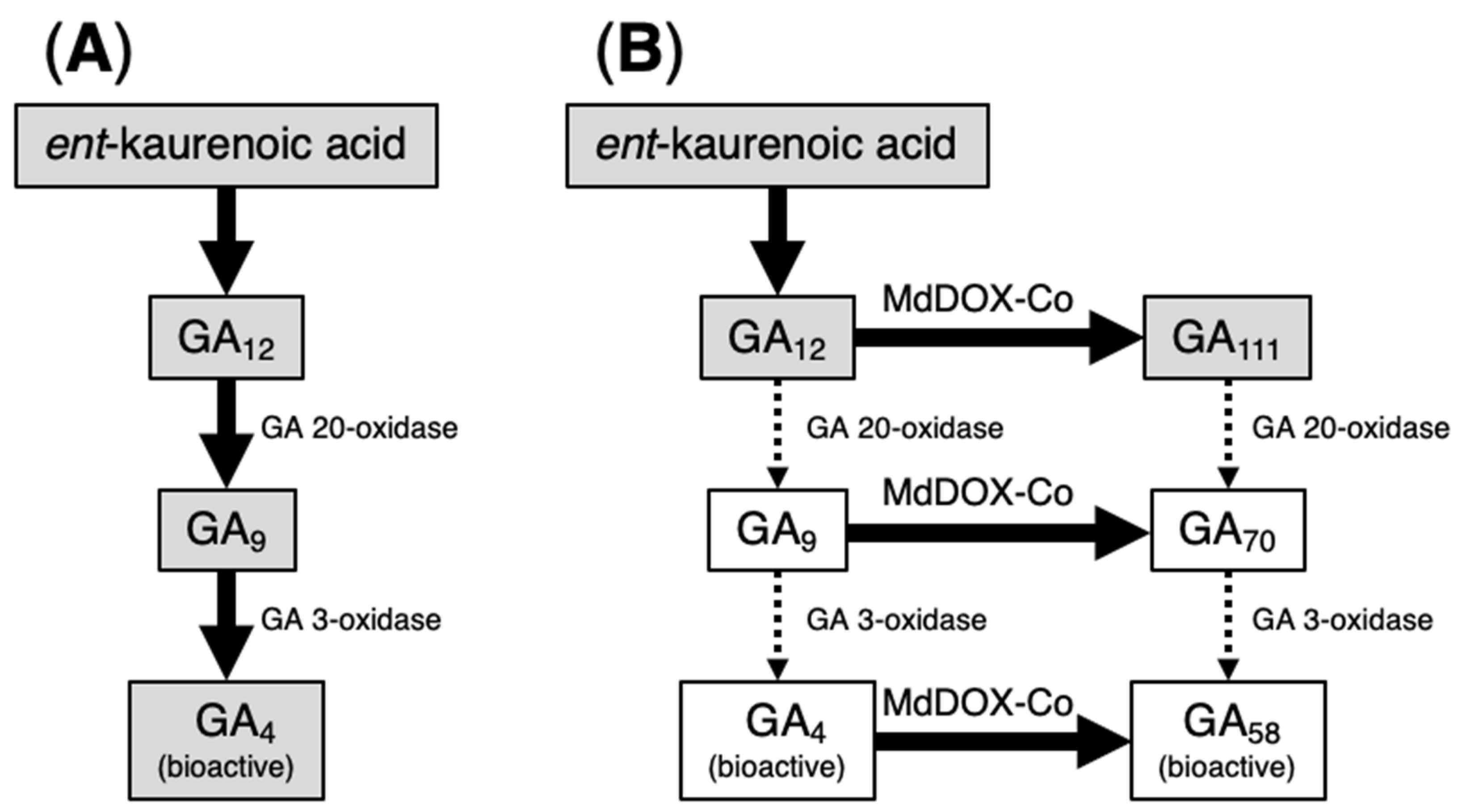
7. Characterization of MdDOX-Co
8. Functions of MdDOX-Co and the Mechanisms of the Columnar Growth Phenotype
9. Modifier Genes and Other Genes Involved in the Columnar Growth Phenotype
10. Marker-Assisted Selection (MAS) Systems for Selecting Columnar Apples
11. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Costes, E.; Lauri, P.É.; Regnard, J.L. Analyzing fruit tree architecture: Implications for tree management and fruit production. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons, Inc: Oxford, UK, 2006; Volume 32, pp. 1–61. [Google Scholar]
- Petersen, R.; Krost, C. Tracing a key player in the regulation of plant architecture: The columnar growth habit of apple trees (Malus × domestica). Planta 2013, 238, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Hollender, C.A.; Dardick, C. Molecular basis of angiosperm tree architecture. New Phytol. 2015, 206, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.L.; Hollender, C.A. Branching out: New insights into the genetic regulation of shoot architecture in trees. Curr. Opin. Plant Biol. 2019, 47, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, L.; Singh, R.; Brown, S.; Dardick, C.; Xu, K. Exploring DNA variant segregation types in pooled genome sequencing enables effective mapping of weeping trait in Malus. J. Exp. Bot. 2018, 69, 1499–1516. [Google Scholar] [CrossRef] [PubMed]
- Lapins, K.O. Inheritance of compact growth type in apple. J. Am. Soc. Hortic. Sci. 1976, 101, 133–135. [Google Scholar]
- Talwara, S.; Grout, B.W.W.; Toldam-Andersen, T.B. Modification of leaf morphology and anatomy as a consequence of columnar architecture in domestic apple (Malus × domestica Borkh.) trees. Sci. Hortic. 2013, 164, 310–315. [Google Scholar] [CrossRef]
- Costes, E.; Gion, J.M. Genetics and genomics of tree architecture. Adv. Bot. Res. 2015, 74, 157–200. [Google Scholar]
- Okada, K. Columnar growth phenotype in apple results from a deficiency of bioactive gibberellin. Kajitsu Nippon 2021, 76, 70–73. (In Japanese) [Google Scholar]
- Okada, K. Molecular mechanism and maker-assisted selection of columnar apples. Agric. Biotechnol. 2019, 3, 8–12. (In Japanese) [Google Scholar]
- Tobutt, K.R. Breeding columnar apples at East Malling. Acta Hortic. 1985, 159, 63–68. [Google Scholar] [CrossRef]
- Fisher, D.V. The ‘Wijcik spur McIntosh’. Fruit Var. J. 1995, 49, 212–213. [Google Scholar]
- Kelsey, D.F.; Brown, S.K. ‘McIntosh Wijcik’: A columnar mutation of ‘McIntosh’ apple providing useful in physiology and breeding research. Fruit Var. J. 1992, 46, 83–87. [Google Scholar]
- Sassa, H. Molecular mechanism of the S-RNase-based gametophytic self-incompatibility in fruit trees of Rosaceae. Breed. Sci. 2016, 66, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Inomata, Y.; Kudo, K.; Wada, M.; Masuda, T.; Bessho, H.; Suzuki, K. The influence of the training system on characteristics of tree growth, fruit productivity and dry matter production of columnar-type apple tree ‘Maypole’. Hortic. Res. 2004, 3, 387–392. (In Japanese) [Google Scholar] [CrossRef][Green Version]
- Blazek, J.; Krelinova, J. Tree growth and some other characteristics of new columnar apple cultivars bred in Holovousy, Czech Republic. Hortic. Sci. 2011, 38, 11–20. [Google Scholar] [CrossRef]
- Guitton, B.; Kelner, J.J.; Velasco, R.; Gardiner, S.E.; Chagne, D.; Costes, E. Genetic control of biennial bearing in apple. J. Exp. Bot. 2012, 63, 131–149. [Google Scholar] [CrossRef]
- Iwanami, H.; Moriya-Tanaka, Y.; Honda, C.; Hanada, T.; Wada, M. Apple thinning strategy based on a model predicting flower-bud formation. Sci. Hortic. 2019, 256, 108529. [Google Scholar] [CrossRef]
- Conner, P.J.; Brown, S.K.; Weeden, N.F. Randomly amplified polymorphic DNA-based genetic linkage maps of three apple cultivars. J. Am. Soc. Hortic. Sci. 1997, 122, 350–359. [Google Scholar] [CrossRef]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The genome of the domesticated apple (Malus × domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef]
- Bai, T.; Zhu, Y.; Fernandez-Fernandez, F.; Keulemans, J.; Brown, S.; Xu, K. Fine genetic mapping of the Co locus controlling columnar growth habit in apple. Mol. Genet. Genom. 2012, 287, 437–450. [Google Scholar] [CrossRef][Green Version]
- Moriya, S.; Okada, K.; Haji, T.; Yamamoto, T.; Abe, K. Fine mapping of Co, a gene controlling columnar growth habit located on apple (Malus × domestica Borkh.) linkage group 10. Plant Breed. 2012, 131, 641–647. [Google Scholar] [CrossRef]
- Baldi, P.; Wolters, P.J.; Komjanc, M.; Viola, R.; Velasco, R.; Salvi, S. Genetic and physical characterisation of the locus controlling columnar habit in apple (Malus × domestica Borkh.). Mol. Breed. 2013, 31, 429–440. [Google Scholar] [CrossRef]
- Morimoto, T.; Banno, K. Genetic and physical mapping of Co, a gene controlling the columnar trait of apple. Tree Genet. Genomes 2015, 11, 807. [Google Scholar] [CrossRef]
- Okada, K.; Wada, M.; Moriya, S.; Katayose, Y.; Fujisawa, H.; Wu, J.; Kanamori, H.; Kurita, K.; Sasaki, H.; Fujii, H.; et al. Expression of a putative dioxygenase gene adjacent to an insertion mutation is involved in the short internodes of columnar apples (Malus × domestica). J. Plant Res. 2016, 129, 1109–1126. [Google Scholar] [CrossRef]
- Wolters, P.J.; Schouten, H.J.; Velasco, R.; Si-Ammour, A.; Baldi, P. Evidence for regulation of columnar habit in apple by a putative 2OG-Fe(II) oxygenase. New Phytol. 2013, 200, 993–999. [Google Scholar] [CrossRef]
- Wolters, P.J. Evidence for regulation of columnar habit in apple by a putative 2OG-Fe (II) oxygenase (vol 200, pg 993, 2013). New Phytol. 2015, 207, 928. [Google Scholar]
- Otto, D.; Petersen, R.; Brauksiepe, B.; Braun, P.; Schmidt, E.R. The columnar mutation (“Co gene”) of apple (Malus × domestica) is associated with an integration of a Gypsy-like retrotransposon. Mol. Breed. 2014, 33, 863–880. [Google Scholar] [CrossRef]
- Petersen, R.; Djozgic, H.; Rieger, B.; Rapp, S.; Schmidt, E.R. Columnar apple primary roots share some features of the columnar-specific gene expression profile of aerial plant parts as evidenced by RNA-Seq analysis. BMC Plant Biol. 2015, 15, 34. [Google Scholar] [CrossRef]
- Okada, K.; Wada, M.; Takebayashi, Y.; Kojima, M.; Sakakibara, H.; Nakayasu, M.; Mizutani, M.; Nakajima, M.; Moriya, S.; Shimizu, T.; et al. Columnar growth phenotype in apple results from gibberellin deficiency by ectopic expression of a dioxygenase gene. Tree Physiol. 2020, 40, 1205–1216. [Google Scholar] [CrossRef]
- Wada, M.; Iwanami, H.; Moriya, S.; Hanada, T.; Moriya-Tanaka, Y.; Honda, C.; Shimizu, T.; Abe, K.; Okada, K. A root-localized gene in normal apples is ectopically expressed in aerial parts of columnar apples. Plant Growth Regul. 2018, 85, 389–398. [Google Scholar] [CrossRef]
- Sun, X.; Wen, C.; Hou, H.; Huo, H.; Zhu, J.; Dai, H.; Zhang, Y. Genes involved in strigolactone biosyntheses and their expression analyses in columnar apple and standard apple. Biol. Plant. 2020, 64, 68–76. [Google Scholar] [CrossRef]
- Sun, X.; Wen, C.; Xu, J.; Wang, Y.; Zhu, J.; Zhang, Y. The apple columnar gene candidate MdCoL and the AP2/ERF factor MdDREB2 positively regulate ABA biosynthesis by activating the expression of MdNCED6/9. Tree Physiol. 2021, 41, 1065–1076. [Google Scholar] [CrossRef]
- Wang, L.; Yu, B.; Zhao, Y.; Li, Y.; Guo, J.; Zhu, Y. A putative 2OG-Fe(II) oxygenase’s response to gibberellin deficiency is related to the internodal growth of columnar apples. Acta Physiol. Plant. 2021, 43, 70. [Google Scholar] [CrossRef]
- Watanabe, D.; Takahashi, I.; Jaroensanti-Tanaka, N.; Miyazaki, S.; Jiang, K.; Nakayasu, M.; Wada, M.; Asami, T.; Mizutani, M.; Okada, K.; et al. The apple gene responsible for columnar tree shape reduces the abundance of biologically active gibberellin. Plant J. 2021, 105, 1026–1034. [Google Scholar] [CrossRef] [PubMed]
- Kawai, Y.; Ono, E.; Mizutani, M. Evolution and diversity of the 2-oxoglutarate-dependent dioxygenase superfamily in plants. Plant J. 2014, 78, 328–343. [Google Scholar] [CrossRef]
- Matsuda, J.; Okabe, S.; Hashimoto, T.; Yamada, Y. Molecular cloning of Hyoscyamine 6-beta-hydroxylase, a 2-oxoglutarate- dependent dioxygenase, from cultured roots of Hyoscyamus niger. J. Biol. Chem. 1991, 266, 9460–9464. [Google Scholar] [CrossRef]
- Nakanishi, H.; Yamaguchi, H.; Sasakuma, T.; Nishizawa, N.K.; Mori, S. Two dioxygenase genes, Ids3 and Ids2, from Hordeum vulgare are involved in the biosynthesis of mugineic acid family phytosiderophores. Plant Mol. Biol. 2000, 44, 199–207. [Google Scholar] [CrossRef]
- Spartz, A.K.; Gray, W.M. Plant hormone receptors: New perceptions. Genes Dev. 2008, 22, 2139–2148. [Google Scholar] [CrossRef]
- Bedini, A.; Mercy, L.; Schneider, C.; Franken, P.; Lucic-Mercy, E. Unraveling the initial plant hormone signaling, metabolic mechanisms and plant defense triggering the endomycorrhizal symbiosis behavior. Front. Plant Sci. 2018, 9, 1800. [Google Scholar] [CrossRef]
- Faizan, M.; Faraz, A.; Sami, F.; Siddiqui, H.; Yusuf, M.; Gruszka, D.; Hayat, S. Role of strigolactones: Signalling and crosstalk with other phytohormones. Open Life Sci. 2020, 15, 217–228. [Google Scholar] [CrossRef]
- Sun, X.; Wen, C.; Zhu, J.; Dai, H.; Zhang, Y. Apple columnar gene MdDMR6 increases the salt stress tolerance in transgenic tobacco seedling and apple calli. J. Plant Growth Regul. 2021, 40, 187–196. [Google Scholar] [CrossRef]
- Dougherty, L.; Bai, T.; Brown, S.; Xu, K. Exploring DNA variant segregation types enables mapping loci for recessive phenotypic suppression of columnar growth in apple. Front. Plant Sci. 2020, 11, 692. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, J.; Dai, H. Characterization of transcriptional differences between columnar and standard apple trees using RNA-Seq. Plant Mol. Biol. Report. 2012, 30, 957–965. [Google Scholar] [CrossRef]
- Krost, C.; Petersen, R.; Schmidt, E.R. The transcriptomes of columnar and standard type apple trees (Malus × domestica)—A comparative study. Gene 2012, 498, 223–230. [Google Scholar] [CrossRef]
- Krost, C.; Petersen, R.; Lokan, S.; Brauksiepe, B.; Braun, P.; Schmidt, E.R. Evaluation of the hormonal state of columnar apple trees (Malus × domestica) based on high throughput gene expression studies. Plant Mol. Biol. 2013, 81, 211–220. [Google Scholar] [CrossRef]
- Tobutt, K.R. Breeding columnar apples at East Malling. Sci. Hortic. 1984, 35, 72–77. [Google Scholar] [CrossRef]
- Cmejlova, J.; Vavra, R.; Cmejla, R. A rapid real-time PCR assay for detecting columnar growth in apples (Malus × domestica). Plant Breed. 2020, 139, 1244–1250. [Google Scholar] [CrossRef]
- Claeys, H.; De Bodt, S.; Inze, D. Gibberellins and DELLAs: Central nodes in growth regulatory networks. Trends Plant Sci. 2014, 19, 231–239. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okada, K.; Honda, C. Molecular Mechanisms Regulating the Columnar Tree Architecture in Apple. Forests 2022, 13, 1084. https://doi.org/10.3390/f13071084
Okada K, Honda C. Molecular Mechanisms Regulating the Columnar Tree Architecture in Apple. Forests. 2022; 13(7):1084. https://doi.org/10.3390/f13071084
Chicago/Turabian StyleOkada, Kazuma, and Chikako Honda. 2022. "Molecular Mechanisms Regulating the Columnar Tree Architecture in Apple" Forests 13, no. 7: 1084. https://doi.org/10.3390/f13071084
APA StyleOkada, K., & Honda, C. (2022). Molecular Mechanisms Regulating the Columnar Tree Architecture in Apple. Forests, 13(7), 1084. https://doi.org/10.3390/f13071084