
The Effects of Fire Disturbance on Litter Decomposition and C:N:P Stoichiometry in a Larix gmelinii Forest Ecosystem of Boreal China


Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Data Collection
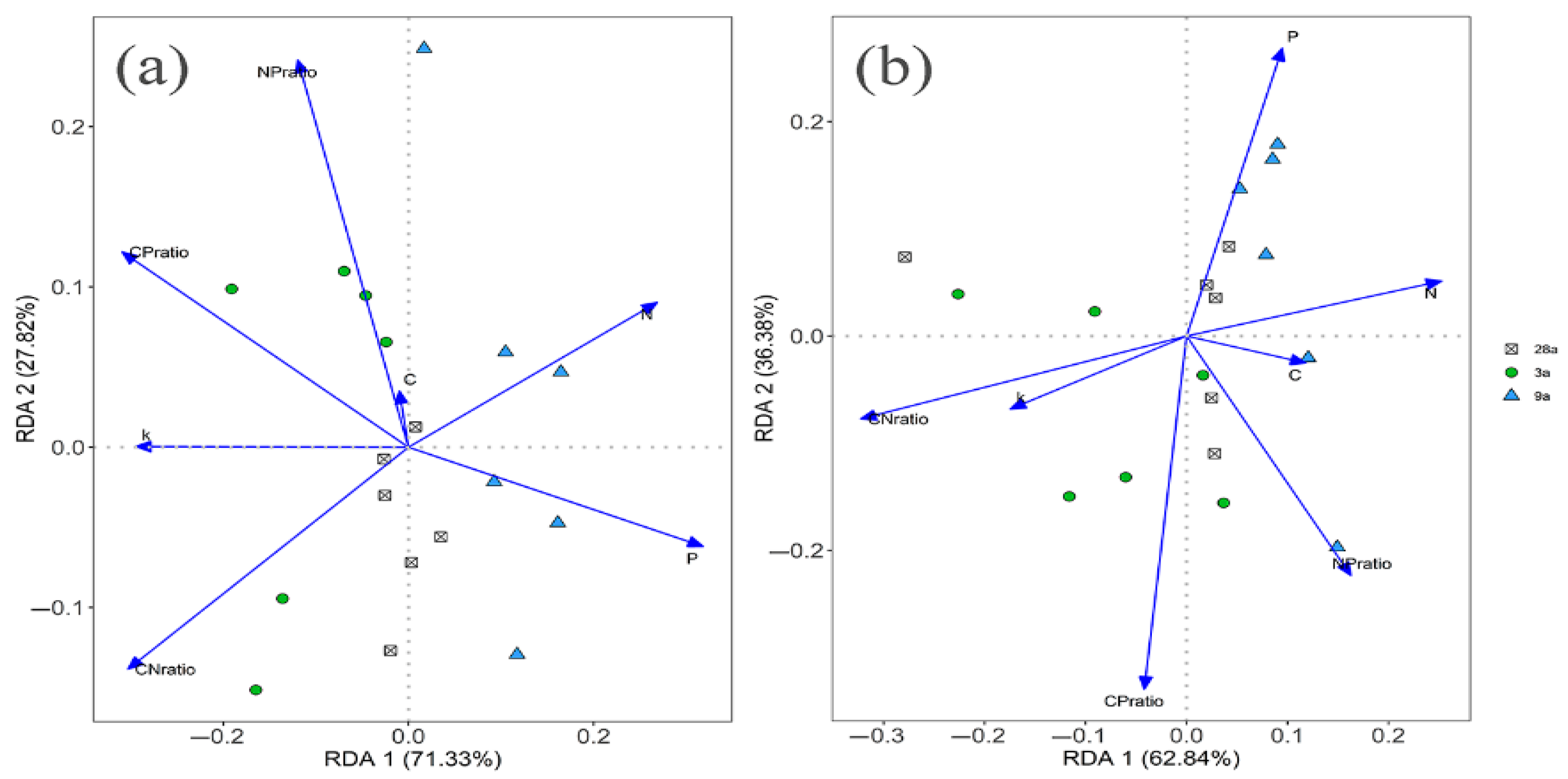
2.4. Data Analysis
2.5. Litter Decomposition Model
3. Results
3.1. Dynamic Change in Litter Decomposition after Fire Disturbance
3.2. Litter C, N, and P Contents and Stoichiometric Ratios
3.3. Litter Stoichiometry and Decomposition Rate
4. Discussion
4.1. Effect of Fire Disturbance on k
4.2. Effect of Fire Disturbance on C, N, and P Stoichiometry
4.3. The Relationship between Litter Nutrient Elements and Decomposition Rate
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disturbance Years | Litter Layer | Control | C Content (g·kg−1) | N Content (g·kg−1) | P Content (g·kg−1) | C:N Ratio | C:P Ratio | N:P Ratio | k |
|---|---|---|---|---|---|---|---|---|---|
| 3 | Upper | Burned | 481.20 ± 0.63 | 3.52 ± 0.04 | 0.56 ± 0.02 | 294.14 ± 58.93 | 953.10 ± 23.89 | 6.31 ± 0.26 | 0.41 ± 0.14 |
| Unburned | 483.49 ± 2.48 | 3.20 ± 0.14 | 0.61 ± 0.03 | 345.68 ± 39.12 | 964.61 ± 119.56 | 6.36 ± 0.58 | 0.33 ± 0.04 | ||
| Lower | Burned | 470.17 ± 1.41 | 2.71 ± 0.18 | 0.85 ± 0.04 | 307.63 ± 89.41 | 611.93 ± 24.53 | 3.36 ± 0.50 | 0.35 ± 0.47 | |
| Unburned | 476.32 ± 2.67 | 3.44 ± 0.15 | 0.74 ± 0.02 | 163.93 ± 5.98 | 767.97 ± 46.17 | 5.71 ± 0.08 | 0.37 ± 0.03 | ||
| p value | B | 0.06558 | 0.17984 | 0.352155 | 0.443 | 0.24216 | 0.01837 * | 0.472 | |
| L | 0.00176 ** | 0.07107 | 0.000174 *** | 0.179 | 0.00366 ** | 0.00217 ** | 0.875 | ||
| L * B | 0.35831 | 0.00489 ** | 0.026756 * | 0.126 | 0.3077 | 0.02184 * | 0.267 | ||
| 9 | Upper | Burned | 481.74 ± 0.57 | 6.39 ± 0.11 | 1.20 ± 0.12 | 101.90 ± 2.62 | 664.55 ± 177.81 | 8.21 ± 1.87 | 0.15 ± 0.03 |
| Unburned | 480.06 ± 1.60 | 5.87 ± 0.74 | 1.26 ± 0.19 | 102.22 ± 12.08 | 541.92 ± 119.68 | 5.94 ± 1.79 | 0.08 ± 0.06 | ||
| Lower | Burned | 482.35 ± 0.29 | 5.09 ± 0.25 | 1.27 ± 0.07 | 114.75 ± 7.17 | 423.69 ± 35.80 | 4.48 ± 0.66 | 0.21 ± 0.01 | |
| Unburned | 480.52 ± 0.98 | 6.33 ± 0.50 | 1.26 ± 0.06 | 90.89 ± 11.36 | 405.20 ± 26.32 | 5.14 ± 0.42 | 0.10 ± 0.06 | ||
| p value | B | 0.114 | 0.4612 | 0.812 | 0.233 | 0.537 | 0.568 | 0.013 * | |
| L | 0.602 | 0.3939 | 0.785 | 0.936 | 0.123 | 0.132 | 0.138 | ||
| L * B | 0.943 | 0.0953 | 0.789 | 0.222 | 0.647 | 0.312 | 0.495 | ||
| 28 | Upper | Burned | 460.79 ± 0.61 | 3.89 ± 0.25 | 0.95 ± 0.04 | 148.42 ± 6.74 | 499.60 ± 20.30 | 4.13 ± 0.28 | 0.14 ± 0.03 |
| Unburned | 467.87 ± 3.63 | 3.55 ± 0.16 | 1.00 ± 0.05 | 159.28 ± 10.77 | 573.33 ± 44.55 | 4.21 ± 0.33 | 0.30 ± 0.05 | ||
| Lower | Burned | 459.01 ± 0.30 | 3.61 ± 0.21 | 0.86 ± 0.14 | 296.33 ± 122.06 | 666.45 ± 71.82 | 5.67 ± 0.79 | 0.23 ± 0.00 | |
| Unburned | 462.86 ± 2.96 | 4.14 ± 0.22 | 0.91 ± 0.03 | 188.32 ± 12.18 | 557.22 ± 19.46 | 4.53 ± 0.23 | 0.22 ± 0.02 | ||
| p value | B | 0.0497 * | 0.6649 | 0.51 | 0.454 | 0.701 | 0.2855 | 0.002 * | |
| L | 0.1891 | 0.4967 | 0.284 | 0.189 | 0.129 | 0.0798 | 0.833 | ||
| L * B | 0.5129 | 0.0765 | 0.999 | 0.363 | 0.074 | 0.2283 | 0.002 * |
| Control | Litter Layer | C Content (g·kg−1) | N Content (g·kg−1) | P Content (g·kg−1) | C:N Ratio | C:P Ratio | N:P Ratio | k | |
|---|---|---|---|---|---|---|---|---|---|
| Burned | Upper | 3 | 481.20 ± 0.63 | 3.52 ± 0.04 | 0.56 ± 0.02 | 294.14 ± 58.93 | 953.10 ± 23.89 | 6.31 ± 0.26 | 0.41 ± 0.14 |
| 9 | 481.74 ± 0.57 | 6.39 ± 0.11 | 1.20 ± 0.12 | 101.90 ± 2.62 | 664.55 ± 177.81 | 8.21 ± 1.87 | 0.15 ± 0.03 | ||
| 28 | 460.79 ± 0.61 | 3.89 ± 0.25 | 0.95 ± 0.04 | 148.42 ± 6.74 | 499.60 ± 20.30 | 4.13 ± 0.28 | 0.14 ± 0.03 | ||
| Lower | 3 | 470.17 ± 1.41 | 2.71 ± 0.18 | 0.85 ± 0.04 | 307.63 ± 89.41 | 611.93 ± 24.53 | 3.36 ± 0.50 | 0.35 ± 0.47 | |
| 9 | 482.35 ± 0.29 | 5.09 ± 0.25 | 1.27 ± 0.07 | 114.75 ± 7.17 | 423.69 ± 35.80 | 4.48 ± 0.66 | 0.21 ± 0.01 | ||
| 28 | 459.01 ± 0.30 | 3.61 ± 0.21 | 0.86 ± 0.14 | 296.33 ± 122.06 | 666.45 ± 71.82 | 5.67 ± 0.79 | 0.23 ± 0.00 | ||
| p value | Y | 0.0000401 *** | 0.838 | 0.778 | 0.767 | 0.1453 | 0.6739 | 0.000 *** | |
| L | 0.163 | 0.217 | 0.505 | 0.39 | 0.0931. | 0.0522 | 0.343 | ||
| L * Y | 0.35 | 0.626 | 0.297 | 0.362 | 0.0126 * | 0.0179 * | 0.141 | ||
| Unburned | Upper | 3 | 483.49 ± 2.48 | 3.20 ± 0.14 | 0.61 ± 0.03 | 345.68 ± 39.12 | 964.61 ± 119.56 | 6.36 ± 0.58 | 0.33 ± 0.04 |
| 9 | 480.06 ± 1.60 | 5.87 ± 0.74 | 1.26 ± 0.19 | 102.22 ± 12.08 | 541.92 ± 119.68 | 5.94 ± 1.79 | 0.08 ± 0.06 | ||
| 28 | 467.87 ± 3.63 | 3.55 ± 0.16 | 1.00 ± 0.05 | 159.28 ± 10.77 | 573.33 ± 44.55 | 4.21 ± 0.33 | 0.30 ± 0.05 | ||
| Lower | 3 | 476.32 ± 2.67 | 3.44 ± 0.15 | 0.74 ± 0.02 | 163.93 ± 5.98 | 767.97 ± 46.17 | 5.71 ± 0.08 | 0.37 ± 0.03 | |
| 9 | 480.52 ± 0.98 | 6.33 ± 0.50 | 1.26 ± 0.06 | 90.89 ± 11.36 | 405.20 ± 26.32 | 5.14 ± 0.42 | 0.10 ± 0.06 | ||
| 28 | 462.86 ± 2.96 | 4.14 ± 0.22 | 0.91 ± 0.03 | 188.32 ± 12.18 | 557.22 ± 19.46 | 4.53 ± 0.23 | 0.22 ± 0.02 | ||
| p value | Y | 0.0000429 *** | 0.71 | 0.496 | 0.435 | 0.0987. | 0.0391 * | 0.000 *** | |
| L | 0.112 | 0.551 | 0.936 | 0.184 | 0.2448 | 0.5468 | 0.968 | ||
| L * Y | 0.983 | 0.856 | 0.581 | 0.081 | 0.4492 | 0.4554 | 0.098 | ||
| Fitting Element | Control | Picture Number | Linear Equation | R Square | p Value |
|---|---|---|---|---|---|
| C | Burned | a | y = 467.37 + 20.67x | 0.03937 | 0.430 |
| Unburned | b | y = 477.42 − 9.74x | 0.01695 | 0.607 | |
| N | Burned | c | y = 6.39 − 8.72x | 0.45138 | 0.002 |
| Unburned | d | y = 6.85 − 10.62x | 0.75055 | 0.000 | |
| P | Burned | e | y = 1.43 − 1.94x | 0.49177 | 0.001 |
| Unburned | f | y = 1.42 − 1.99x | 0.64738 | 0.000 | |
| C:Nratio | Burned | g | y = 21.79 + 753.95x | 0.30394 | 0.018 |
| Unburned | h | y = 63.97 + 485.78x | 0.36926 | 0.007 | |
| C:Pratio | Burned | i | y = 345.12 + 1164.22x | 0.29938 | 0.019 |
| Unburned | j | y = 348.82 + 1251.7x | 0.42756 | 0.003 | |
| N:Pratio | Burned | k | y = 6.34 − 3.92x | 0.03296 | 0.471 |
| Unburned | l | y = 5.25 + 0.3x | 0.00058 | 0.924 |
References
- Kong, J.J.; Yang, J. A Review of effects of fire disturbance on soil environment in boreal coniferous forests. Chin. J. Soil. Sci. 2014, 45, 291–296. [Google Scholar] [CrossRef]
- Hu, T.X.; Hu, H.Q.; Li, F.; Zhao, B.Q.; Wu, S.; Zhu, G.Y.; Sun, L. Long-term effects of post-fire restoration types on nitrogen mineralisation in a Dahurian larch (Larix gmelinii) forest in boreal China. Sci. Total. Environ. 2019, 679, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Heon, J.; Arseneault, D.; Parisien, M.A. Resistance of the boreal forest to high burn rates. Proc. Natl. Acad. Sci. USA 2014, 111, 13888–13893. [Google Scholar] [CrossRef] [PubMed]
- Wardle, D.A.; Nilsson, M.C.; Zackrisson, O. Fire-derived charcoal causes loss of forest humus. Science 2008, 320, 629. [Google Scholar] [CrossRef] [PubMed]
- Pugh, T.A.M.; Arneth, A.; Kautz, M.; Poulter, B.; Smith, B. Important role of forest disturbances in the global biomass turnover and carbon sinks. Nat. Geosci. 2019, 12, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Prescott, C.E.; Vesterdal, L. Decomposition and transformations along the continuum from litter to soil organic matter in forest soils. For. Ecol. Manag. 2021, 498, 119522. [Google Scholar] [CrossRef]
- Prescott, C.E. Litter decomposition: What controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 2010, 101, 133–149. [Google Scholar] [CrossRef]
- Certini, G.; Moya, D.; Lucas-Borja, M.E.; Mastrolonardo, G. The impact of fire on soil-dwelling biota: A review. For. Ecol. Manag. 2021, 488, 118989. [Google Scholar] [CrossRef]
- Petraglia, A.; Cacciatori, C.; Chelli, S.; Fenu, G.; Calderisi, G.; Gargano, D.; Abeli, T.; Orsenigo, S.; Carbognani, M. Litter decomposition: Effects of temperature driven by soil moisture and vegetation type. Plant Soil 2019, 435, 187–200. [Google Scholar] [CrossRef]
- Hopkins, J.R.; Semenova-Nelsen, T.; Sikes, B.A. Fungal community structure and seasonal trajectories respond similarly to fire across pyrophilic ecosystems. Fems Microbiol. Ecol. 2021, 97, fiaa219. [Google Scholar] [CrossRef]
- Pellegrini, A.F.A.; Caprio, A.C.; Georgiou, K.; Finnegan, C.; Hobbie, S.E.; Hatten, J.A.; Jackson, R.B. Low-intensity frequent fires in coniferous forests transform soil organic matter in ways that may offset ecosystem carbon losses. Glob. Chang. Biol. 2021, 27, 3810–3823. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.Y.; Guan, L.L.; Wei, X.H.; Tang, X.L.; Liu, S.G.; Liu, J.X.; Zhang, D.Q.; Yan, J.H. Factors influencing leaf litter decomposition: An intersite decomposition experiment across China. Plant Soil 2008, 311, 61–72. [Google Scholar] [CrossRef]
- Kuruppuarachchi, K.; Seneviratne, G.; Madurapperuma, B.D. Carbon Sequestration in Tropical Forest Stands: Its Control by Plant, Soil and Climatic Factors. Open J. For. 2016, 6, 59–71. [Google Scholar] [CrossRef]
- Hatten, S.S.; Tiunov, A.V.; Scheu, S. Biodiversity and litter decomposition interrestrial ecosystems. Ann. Rev. Ecol. Evo. Syst. 2005, 36, 191–218. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, J.H.C.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Harguindeguy, N.P.; et al. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef]
- Yin, R.; Eisenhauer, N.; Schmidt, A.; Gruss, I.; Purahong, W.; Siebert, J.; Schadler, M. Climate change does not alter land-use effects on soil fauna communities. Appl. Soil. Ecol. 2019, 140, 1–10. [Google Scholar] [CrossRef]
- Yang, X.D.; Zhao, Y.; Warren, M.W.; Chen, J. Mechanical fragmentation enhances the contribution of Collembola to leaf litter decomposition. Eur. J. Soil. Biol. 2012, 53, 23–31. [Google Scholar] [CrossRef]
- Ludwig, S.M.; Alexander, H.D.; Kielland, K.; Mann, P.J.; Natali, S.M.; Ruess, R.W. Fire severity effects on soil carbon and nutrients and microbial processes in a Siberian larch forest. Glob. Chang. Biol. 2018, 24, 5841–5852. [Google Scholar] [CrossRef]
- Stirling, E.; Macdonald, L.M.; Smernik, R.J.; Cavagnaro, T.R. Post fire litters are richer in water soluble carbon and lead to increased microbial activity. Appl. Soil. Ecol. 2019, 136, 101–105. [Google Scholar] [CrossRef]
- Hulugalle, N.R.; Strong, C.; Mcpherson, K.; Nachimuthu, G. Carbon, nitrogen and phosphorus stoichiometric ratios under cotton cropping systems in Australian Vertisols: A meta-analysis of seven experiments. Nutr. Cycl. Agroecosystems 2017, 107, 357–367. [Google Scholar] [CrossRef]
- Schafer, J.L.; Mack, M.C. Nutrient limitation of plant productivity in scrubby flatwoods: Does fire shift nitrogen versus phosphorus limitation? Plant Ecol. 2018, 219, 1063–1079. [Google Scholar] [CrossRef]
- Butler, O.M.; Lewis, T.; Rashti, R.M.; Maunsell, S.C.; Elser, J.J.; Chen, C.R. The stoichiometric legacy of fire regime regulates the roles of micro-organisms and invertebrates in decomposition. Ecology 2019, 100, e02732. [Google Scholar] [CrossRef] [PubMed]
- De, L.J.R.; Dorrepaal, E.; Kardol, P.; Nilsson, M.C.; Teuber, L.M.; Wardle, D.A. Understory plant functional groups and litter species identity are stronger drivers of litter decomposition than warming along a boreal forest post-fire disturbance successional gradient. Soil Biol. Biochem. 2016, 98, 159–170. [Google Scholar] [CrossRef]
- Rodrigo, A.; Arnan, X.; Retana, J. Relevance of soil seed bank and seed rain to immediate seed supply after a large wildfire. Int. J. Wildland Fire 2012, 21, 449–458. [Google Scholar] [CrossRef]
- Wang, W.Q.; Wang, C.; Sardans, J.; Zeng, C.S.; Josep, P.; Tong, C. Plant invasive success associated with higher N-use efficiency and stoichiometric shifts in the soil–plant system in the Minjiang River tidal estuarine wetlands of China. Wetl. Ecol. Manag. 2015, 23, 865–880. [Google Scholar] [CrossRef]
- Osburn, E.D.; Elliottt, K.J.; Knoepp, J.D.; Miniat, C.F.; Barrett, J.E. Soil microbial response to Rhododendron understory removal in southern Appalachian forests: Effects on extracellular enzymes. Soil. Biol. Biochem. 2018, 127, 50–59. [Google Scholar] [CrossRef]
- Jones, G.L.; Tomlinson, M.; Owen, R.; Scullion, J.; Winters, A.; Jenkins, T.; Ratcliffe, J.; Jones, D.J. Shrub establishment favoured and grass dominance reduced in acid heath grassland systems cleared of invasive Rhododendron ponticum. Sci. Rep. 2019, 9, 2239. [Google Scholar] [CrossRef]
- Hernandez, E.; Questad, E.J.; Meyer, W.M.; Suding, K.N. The effects of nitrogen deposition and invasion on litter fuel quality and decomposition in a Stipa pulchra grassland. J. Arid. Environ. 2019, 162, 35–44. [Google Scholar] [CrossRef]
- Duran, J.; Rodriguez, A.; Mendez, J.; Morales, G.; Palacios, J.M.F.; Gallardo, A. Wildfire decrease the local-scale ecosystem spatial variability of Pinus canariensis forests during the first two decades post fire. Int. J. Wildland Fire 2019, 28, 288–294. [Google Scholar] [CrossRef]
- Kurunthachaiam, S.K.; Sajwan, K.S.; Kumar, A.A.; Manian, S. Effects of surface fire on litter decomposition and occurrence of microfungi in a Cymbopogon polyneuros dominated grassland. Arch. Agron. Soil. Sci. 2007, 53, 205–219. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Grootemaat, S.; Verheijen, L.M.; Cornwell, W.K.; Bodegom, P.M.V.; Wal, R.V.D.; Aerts, R. Are litter decomposition and fire linked through plant species traits? New Phytol. 2017, 216, 653–669. [Google Scholar] [CrossRef] [PubMed]
- Throop, H.L.; Salem, M.A.; Whitford, W.G. Fire enhances litter decomposition and reduces vegetation cover influences on decomposition in a dry woodland. Plant Ecol. 2017, 218, 799–811. [Google Scholar] [CrossRef]
- Holden, S.R.; Gutierrez, A.; Treseder, K.K. Changes in soil fungal communities, extracellular enzyme activities, and litter decomposition across a fire chronosequence in Alaskan Boreal Forests. Ecosystems 2013, 16, 34–46. [Google Scholar] [CrossRef]
- Yang, X.F.; Bao, X.L.; Hu, G.Q.; Shao, S.; Zhou, F.; Ye, J.S.; Xie, H.T.; Liang, C. C:N:P stoichiometry characteristics of litter and soil of forests in Great Xing’an Mountains with different fire years. Chin. J. Appl. Ecol. 2016, 27, 1359–1367. [Google Scholar] [CrossRef]
- Wei, M.Y.; Zhong, Y.Q.W.; Zhu, G.Y.; Liu, W.Z.; Shangguan, Z.P. Nutrient limitation of litter decomposition with long-term secondary succession: Evidence from controlled laboratory experiments. J. Soils Sediments 2020, 20, 1858–1868. [Google Scholar] [CrossRef]
- Butler, O.M.; Lewis, T.; Chen, C.R. Fire alters soil labile stoichiometry and litter nutrients in Australian eucalypt forests. Int. J. Wildland Fire 2017, 26, 783–788. [Google Scholar] [CrossRef]
- Santin, C.; Otero, X.L.; Doerr, S.H.; Chafer, C.J. Impact of a moderate/high-severity prescribed eucalypt forest fire on soil phosphorous stocks and partitioning. Sci. Total Environ. 2018, 621, 1103–1114. [Google Scholar] [CrossRef]
- Hume, A.; Chen, H.Y.H.; Taylor, A.R.; Kayahara, G.J.; Man, R.Z. Soil C:N:P dynamics during secondary succession following fire in the boreal forest of central Canada. For. Ecol. Manag. 2016, 369, 1–9. [Google Scholar] [CrossRef]
- Toberman, H.; Chen, C.R.; Lewis, T.; Elser, J.J. High-frequency fire alters C:N:P stoichiometry in forest litter. Glob. Chang. Biol. 2014, 20, 2321–2331. [Google Scholar] [CrossRef]
- Morrison, I.K. Decomposition and element release from confined jack pine needle litter on and in the feathermoss layer. Can. J. For. Res. 2003, 33, 16–22. [Google Scholar] [CrossRef]
- Hilli, S.; Stark, S.; Derome, J. Litter decomposition rates in relation to litter stocks in boreal coniferous forests along climatic and soil fertility gradients. Appl. Soil Ecol. 2010, 46, 200–208. [Google Scholar] [CrossRef]
- Jackson, B.G.; Nilsson, M.C.; Wardle, D.A. The effects of the moss layer on the decomposition of intercepted vascular plant litter across a post-fire boreal forest chronosequence. Plant Soil 2013, 367, 199–214. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Zheng, J.Q.; Xu, Z.H.; Abdullah, K.M.; Zhou, Q.X. Effects of changed litter inputs on soil labile carbon and nitrogen pools in a eucalyptus-dominated forest of southeast Queensland, Australia. J. Soils Sediments 2019, 19, 1661–1671. [Google Scholar] [CrossRef]
- Zhang, R.; Hu, H.Q.; Qu, Z.L.; Hu, T.X. Diurnal variation models for fine fuel moisture content in boreal forests in China. J. For. Res. 2020, 32, 1177–1187. [Google Scholar] [CrossRef]
- Hu, T.X.; Sun, L.; Hu, H.Q.; Guo, F.T. Effects of fire disturbance on soil respiration in the non-growing season in a Larix gmelinii forest in the Daxing’an Mountains, China. PLoS ONE 2017, 12, e0180214. [Google Scholar] [CrossRef]
- Li, S.; Wu, Z.W.; Yu, L.; He, H.S. The temporal and spatial clustering characteristics of forest fire in the Great Xing’an Mountains. Chin. J. Ecol. 2017, 36, 198–204. [Google Scholar] [CrossRef]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Zhang, D.Q.; Hui, D.F.; Luo, Y.Q.; Zhou, G.Y. Rates of litter decomposition in terrestrial ecosystems: Global patterns and controlling factors. J. Plant Ecol. 2008, 1, 85–93. [Google Scholar] [CrossRef]
- Dearden, F.M.; Dehlin, H.; Wardle, D.; Nilsson, M.C. Changes in the ratio of twig to foliage in litterfall with species composition, and consequences for decomposition across a long term chronosequence. Oikos 2006, 115, 453–462. [Google Scholar] [CrossRef]
- Wardle, D.A.; Hornberg, G.; Zackrisson, O.; Kalela-Brundin, M.; Coomes, D.A. Long-term effects of wildfire on ecosystem properties across an island area gradient. Science 2003, 300, 972–975. [Google Scholar] [CrossRef]
- Hernandez, D.L.; Hobbie, S.E. Effects of fire frequency on oak litter decomposition and nitrogen dynamics. Oecologia 2008, 158, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Kay, A.D.; Mankowski, J.; Hobbie, S.E. Long-term burning interacts with herbivory to slow decomposition. Ecology 2008, 89, 1188–1194. [Google Scholar] [CrossRef] [PubMed]
- Bryanin, S.; Kondratova, A.; Abramova, E. Litter decomposition and nutrient dynamics in fire-affected larch forests in the Russian far east. Forests 2020, 11, 882. [Google Scholar] [CrossRef]
- Yan, W.; Shangguan, Z.; Zhong, Y. Responses of mass loss and nutrient release in litter decomposition to ultraviolet radiation. J. Soils Sediments 2021, 21, 698–704. [Google Scholar] [CrossRef]
- Talhelm, A.F.; Smith, A.M.S. Litter moisture adsorption is tied to tissue structure, chemistry, and energy concentration. Ecosphere 2018, 9, 18. [Google Scholar] [CrossRef]
- Musetta-Lambert, J.; Muto, E.; Kreutzweiser, D.; Sibley, P. Wildfire in boreal forest catchments influences leaf litter subsidies and consumer communities in streams: Implications for riparian management strategies. For. Ecol. Manag. 2017, 391, 29–41. [Google Scholar] [CrossRef]
- Köster, K.; Berninger, F.; Heinonsalo, J.; Linden, A.; Köster, E.; Ilvesniemi, H.; Pumpanen, J. The long-term impact of low-intensity surface fires on litter decomposition and enzyme activities in boreal coniferous forests. Int. J. Wildland Fire 2016, 25, 213–223. [Google Scholar] [CrossRef]
- Springett, J.A. Effects of a single hot summer fire on soil fauna and on litter decomposition in jarrah (Eucalyptus marginata) forest in Western Australia. Aust. J. Ecol. 2010, 4, 279–291. [Google Scholar] [CrossRef]
- Jurskis, V.; Turner, J.; Lambert, M.; Bi, H.Q. Fire and N cycling: Getting the perspective right. Appl. Veg. Sci. 2011, 14, 433–434. [Google Scholar] [CrossRef]
- Chen, Y.M.; Liu, Y.; Zhang, J.; Yang, W.Q.; He, R.L.; Deng, C.C. Microclimate exerts greater control over litter decomposition and enzyme activity than litter quality in an alpine forest-tundra ecotone. Sci. Rep. 2018, 8, 14998. [Google Scholar] [CrossRef]
- Gao, L.W.; Zhang, M.Q.; Liu, Y.; Vicente, M.L. Litter cover promotes biocrust decomposition and surface soil functions in sandy ecosystem. Geoderma 2020, 374, 114429. [Google Scholar] [CrossRef]
- Bogorodskaya, A.V.; Ivanova, G.A.; Tarasov, P.A. Post-fire transformation of the microbial complexes in soils of larch forests in the lower Angara River region. Eurasian Soil Sci. 2011, 44, 49–55. [Google Scholar] [CrossRef]
- Ondřej, V.; Kalwij, J.M.; Hédl, R. Effects of simulated historical tree litter raking on the understorey vegetation in a central European forest. Appl. Veg. Sci. 2015, 18, 569–578. [Google Scholar] [CrossRef]
- Weber, M.G. Decomposition, litter fall, and forest floor nutrient dynamics in relation to fire disturbance in eastern Ontario jack pine ecosystems. Can. J. For. Res. 1987, 17, 1496–1506. [Google Scholar] [CrossRef]
- Davies, A.B.; Rensburg, B.J.V.; Eggleton, P.; Parr, C.L. Interactive Effects of Fire disturbance, Rainfall, and Litter Quality on Decomposition in Savannas: Frequent Fire Leads to Contrasting Effects. Ecosystems 2013, 16, 866–880. [Google Scholar] [CrossRef]
- Garcia, P.P.; Prieto, I.; Ourcival, J.M.; Hatten, S.S. Disentangling the litter quality and soil microbial contribution to leaf and fine root litter decomposition responses to reduced rainfall. Ecosystems 2016, 19, 490–503. [Google Scholar] [CrossRef]
- Urgenson, L.S.; Ryan, C.M.; Halpern, C.B.; Bakker, J.D.; Belote, R.T.; Franklin, J.F.; Haugo, R.D.; Nelson, C.R.; Waltz, A.E.M. Erratum to: Visions of restoration in fire-adapted forest landscapes: Lessons from the collaborative forest landscape Restoration Program. Environ. Manag. 2017, 59, 354–355. [Google Scholar] [CrossRef]
- Hoeber, S.; Fransson, P.; Weih, M.; Stefano, M. Leaf litter quality coupled to Salix variety drives litter decomposition more than stand diversity or climate. Plant Soil 2020, 453, 313–328. [Google Scholar] [CrossRef]
- Coleman, T.W.; Rieske, L.K. Arthropod response to prescription burning at the soil-litter interface in oak-pine forests. For. Ecol. Managt. 2006, 233, 52–60. [Google Scholar] [CrossRef]
- Hartshorn, J. A Review of Forest Management Effects on Terrestrial Leaf Litter Inhabiting Arthropods. Forests 2021, 12, 23. [Google Scholar] [CrossRef]
- Gartner, T.B.; Treseder, K.K.; Malcolm, G.M.; Sinsabaugh, R.L. Extracellular enzyme activity in the mycorrhizospheres of a boreal fire chronosequence. Pedobiologia 2012, 55, 121–127. [Google Scholar] [CrossRef]
- Knelman, J.E.; Graham, E.B.; Ferrenberg, S.; Lecoeuvre, A.; Labrado, A.; Darcy, J.L.; Nemergut, D.R.; Schmidt, S.K. Rapid shifts in soil nutrients and decomposition enzyme activity in early succession following forest fire. Forests 2017, 8, 347. [Google Scholar] [CrossRef]
- Auclerc, A.; Le Moine, J.M.; Hatton, P.J.; Bird, J.A.; Nadelhoffer, K.J. Decadal post-fire succession of soil invertebrate communities is dependent on the soil surface properties in a boreal temperate forest. Sci. Total. Environ. 2019, 647, 1058–1068. [Google Scholar] [CrossRef]
- Gui, H.; Hyde, K.; Xu, J.C.; Mortimer, P. Arbuscular mycorrhiza enhance the rate of litter decomposition while inhibiting soil microbial community development. Sci. Rep. 2017, 7, 42184. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.T.; Gutknecht, J.; Nadrowski, K.; Geißler, C.; Kühn, P.; Scholten, T.; Both, S.; Erfmeier, A.; Böhnke, M.; Bruelheide, H.; et al. Relationships between soil microorganisms, plant communities, and soil characteristics in Chinese subtropical forests. Ecosystems 2012, 15, 624–636. [Google Scholar] [CrossRef]
- Zhang, P.; Li, B.; Wu, J.H.; Hu, S.J. Invasive plants differentially affect soil biota through litter and rhizosphere pathways: A meta-analysis. Ecol. Lett. 2019, 22, 200–210. [Google Scholar] [CrossRef]
- Hicks, L.C.; Meir, P.; Nottingham, A.T.; Reay, D.S.; Stott, A.W.; Salinas, N.; Whitaker, J. Carbon and nitrogen inputs differentially affect priming of soil organic matter in tropical lowland and montane soils. Soil Biol. Biochem. 2019, 129, 212–222. [Google Scholar] [CrossRef]
- Mayor, A.G.; Goiran, S.B.; Vallejo, V.R.; Bautista, S. Variation in soil enzyme activity as a function of vegetation amount, type, and spatial structure in fire-prone Mediterranean shrublands. Sci. Total. Environ. 2016, 573, 1209–1216. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, S.Y.; Cheng, Y.; Cai, Z.C.; Muller, C.; Zhang, J.B. Composition of soil recalcitrant C regulates nitrification rates in acidic soils. Geoderma 2019, 337, 965–972. [Google Scholar] [CrossRef]
- Grigal, D.F.; Mccoll, J.G. Litter decomposition following forest fire in Northeastern Minnesota. J. Appl. Ecol. 1977, 14, 531–538. [Google Scholar] [CrossRef]
- Brödlin, D.; Kaiser, K.; Kessler, A.; Hagedorn, F. Drying and rewetting foster phosphorus depletion of forest soils. Soil Biol. Biochem. 2019, 128, 22–34. [Google Scholar] [CrossRef]
- Wang, X.G.; Lu, X.T.; Han, X.G. Responses of nutrient concentrations and stoichiometry of senesced leaves in dominant plants to nitrogen addition and prescribed burning in a temperate steppe. Ecol. Eng. 2014, 70, 154–161. [Google Scholar] [CrossRef]
- Smith, S.W.; Speed, J.D.M.; Bukombe, J.; Hassan, S.N.; Lyamuya, R.D.; Mtweve, P.J.; Sundsdal, A.; Graae, B.J. Litter type and termites regulate root decomposition across contrasting savanna land-uses. Oikos 2019, 128, 596–607. [Google Scholar] [CrossRef]
- Zheng, Z.M.; Mamuti, M.; Liu, H.M.; Shu, Y.Q.; Hu, S.J.; Wang, X.H.; Li, B.B.; Lin, L.; Li, X. Effects of nutrient additions on litter decomposition regulated by phosphorus-induced changes in litter chemistry in a subtropical forest, China. For. Ecol. Manag. 2017, 400, 123–128. [Google Scholar] [CrossRef]
- Jiang, T.H.; Lu, X.Y.; Ma, X.X.; Wang, X.D. Five-year study on the effects of warming and plant litter quality on litter decomposition rate in a Tibetan alpine grassland. Sci. Total Environ. 2021, 750, 142306. [Google Scholar] [CrossRef]
- Xiao, W.Y.; Chen, H.Y.H.; Kumar, P.; Chen, C.; Guan, W.Q. Multiple interactions between tree composition and diversity and microbial diversity underly litter decomposition. Geoderma 2019, 341, 161–171. [Google Scholar] [CrossRef]
- Saura-Mas, S.; Estiarte, M.; Penuelas, J.; Lloret, F. Effects of climate change on leaf litter decomposition across post-fire plant regenerative groups. Environ. Exp. Bot. 2012, 77, 274–282. [Google Scholar] [CrossRef]
- Lin, C.F.; Peng, J.Q.; Hong, H.B.; Yang, Z.J.; Yang, Y.S. Effect of nitrogen and phosphorus availability on forest litter decomposition. Acta Ecol. Sin. 2017, 37, 54–62. [Google Scholar] [CrossRef][Green Version]
- Bengtsson, J.; Janion, C.; Chown, S.L.; Leinaas, H.P. Litter decomposition in fynbos vegetation, South Africa. Soil Biol. Biochem. 2012, 47, 100–105. [Google Scholar] [CrossRef]
- Chacón, N.; Dezzeo, N. Litter decomposition in primary forest and adjacent fire -disturbed forests in the Gran Sabana, southern Venezuela. Biol. Fertil. Soils 2007, 43, 815–821. [Google Scholar] [CrossRef]
- Jabiol, J.; Lecerf, A.; Lamothe, S.; Gessner, M.O.; Chauvet, E. Litter quality modulates effects of dissolved Nitrogen on leaf decomposition by stream microbial communities. Microb. Ecol. 2019, 77, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Taylor, Q.A.; Midgley, M.G. Prescription side effects: Long-term, high-frequency controlled burning enhances nitrogen availability in an Illinois oak-dominated forest. For. Ecol. Manag. 2018, 411, 82–89. [Google Scholar] [CrossRef]
- Zhou, X.; Sun, H.; Pumpanen, J.; Sietio, O.M.; Heinonsalo, J.; Koster, K.; Berninger, F. The impact of wildfire on microbial C:N P stoichiometry and the fungal-to-bacterial ratio in permafrost soil. Biogeochemistry 2019, 142, 1–17. [Google Scholar] [CrossRef]
- Gusewell, S.; Gessner, M.O. N:P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms. Funct. Ecol. 2009, 23, 211–219. [Google Scholar] [CrossRef]
- Ficken, C.; Wright, J.P. Effects of fire frequency on litter decomposition as mediated by changes to litter chemistry and soil environmental conditions. PLoS ONE 2017, 12, e0186292. [Google Scholar] [CrossRef]
- Brennan, K.E.C.; Christie, F.J.; York, A. Global climate change and litter decomposition: More frequent fire slows decomposition and increases the functional importance of invertebrates. Glob. Chang. Biol. 2009, 15, 2958–2971. [Google Scholar] [CrossRef]
- Heim, R.J.; Heim, W.; Darman, G.F.; Thilo, H.; Sergei, M.S.; Norbert, H. Litter removal through fire—A key process for wetland vegetation and ecosystem dynamics. Sci. Total Environ. 2021, 755, 142659. [Google Scholar] [CrossRef]
- Johnston, J.D.; Olszewski, J.H.; Miller, B.; ASchmidt, M.R.; Vernon, M.J.; Ellsworth, L.M. Mechanical thinning without prescribed fire moderates wildfire behavior in an Eastern Oregon, USA ponderosa pine forest. For. Ecol. Manag. 2021, 501, 119674. [Google Scholar] [CrossRef]






| Sample | 3YSF | 9YSF | 28YSF | |||
|---|---|---|---|---|---|---|
| Unburned | Burned | Unburned | Burned | Unburned | Burned | |
| Dominant species | 1 * | 1 *,2 | 1 *,2 | 1 *,2 | 1 *,2,3 | 1 *,2 |
| Mean height(cm) | 26.2 ± 2.0 | 18.1 ± 3.2 | 14.4 ± 2.3 | 13.0 ± 1.5 | 22.0 ± 3.5 | 14.3 ± 1.6 |
| Average tree height (m) | 24.7 ± 1.5 | 20.1 ± 2.2 | 17.0 ± 1.3 | 16.5 ± 0.6 | 22.5 ± 1.1 | 16.4 ± 0.8 |
| Canopy density | 0.8 ± 0.2 | 0.7 ± 0.1 | 0.7 ± 0.1 | 0.7 ± 0.1 | 0.7 ± 0.1 | 0.6 ± 0.1 |
| Age (a) | 70 | 45 | 50 | 50 | 60 | 55 |
| Humus thickness (cm) | 9.2 ± 1.2 | 2.5 ± 0.2 | 10.5 ± 1.0 | 6.2 ± 0.6 | 11.5 ± 2.3 | 6.3 ± 0.5 |
| Soil TOC (g·kg−1) | 54.26 ± 4.40 | 74.20 ± 25.31 | 83.29 ± 26.12 | 99.25 ± 9.69 | 140.88 ± 34.53 | 76.16 ± 32.03 |
| Soil TON (g·kg−1) | 6.94 ± 0.78 | 7.56 ± 0.74 | 4.43 ± 1.40 | 6.75 ± 1.62 | 9.59 ± 1.02 | 5.72 ± 0.18 |
| Soil AP (mg·kg−1) | 40.67 ± 4.55 | 54.35 ± 23.47 | 57.71 ± 6.29 | 13.49 ± 2.29 | 28.66 ± 7.17 | 26.55 ± 16.47 |
| Soil bulk density (g·cm−3) | 0.82 ± 0.01 | 0.58 ± 0.08 | 0.80 ± 0.06 | 0.61 ± 0.16 | 0.65 ± 0.27 | 0.62 ± 0.04 |
| Post-Fire Disturbance Ages | Treat | Fitting Equation | k Value | R Square | Years Until Half Decomposition (a) | Time of 95% Decomposition (a) |
|---|---|---|---|---|---|---|
| 3 | BU | y = 1.0278e−0.405x | 0.405 | 0.7899 | 1.78 | 7.46 |
| BL | y = 1.0204e−0.35x | 0.35 | 0.9136 | 2.04 | 8.62 | |
| UU | y = 1.0302e−0.324x | 0.324 | 0.674 | 2.23 | 9.34 | |
| UL | y = 1.0441e−0.366x | 0.366 | 0.6579 | 2.01 | 8.30 | |
| 9 | BU | y = 1.0156e−0.147x | 0.147 | 0.7582 | 4.82 | 20.48 |
| BL | y = 1.0209e−0.211x | 0.211 | 0.8174 | 3.38 | 14.30 | |
| UU | y = 1.0135e−0.077x | 0.077 | 0.5771 | 9.18 | 39.08 | |
| UL | y = 1.013e−0.101x | 0.101 | 0.674 | 6.99 | 29.79 | |
| 28 | BU | y = 1.0223e−0.163x | 0.163 | 0.6687 | 4.39 | 18.51 |
| BL | y = 1.0342e−0.226x | 0.226 | 0.6014 | 3.22 | 13.40 | |
| UU | y = 1.0391e−0.297x | 0.297 | 0.5343 | 2.46 | 10.22 | |
| UL | y = 1.0222e−0.207x | 0.207 | 0.7124 | 3.45 | 14.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Shi, Z.; Zhao, B.; Bono, G.J.; Sun, L.; Hu, T. The Effects of Fire Disturbance on Litter Decomposition and C:N:P Stoichiometry in a Larix gmelinii Forest Ecosystem of Boreal China. Forests 2022, 13, 1029. https://doi.org/10.3390/f13071029
Li F, Shi Z, Zhao B, Bono GJ, Sun L, Hu T. The Effects of Fire Disturbance on Litter Decomposition and C:N:P Stoichiometry in a Larix gmelinii Forest Ecosystem of Boreal China. Forests. 2022; 13(7):1029. https://doi.org/10.3390/f13071029
Chicago/Turabian StyleLi, Fei, Zhe Shi, Bingqing Zhao, Gong Jinhua Bono, Long Sun, and Tongxin Hu. 2022. "The Effects of Fire Disturbance on Litter Decomposition and C:N:P Stoichiometry in a Larix gmelinii Forest Ecosystem of Boreal China" Forests 13, no. 7: 1029. https://doi.org/10.3390/f13071029
APA StyleLi, F., Shi, Z., Zhao, B., Bono, G. J., Sun, L., & Hu, T. (2022). The Effects of Fire Disturbance on Litter Decomposition and C:N:P Stoichiometry in a Larix gmelinii Forest Ecosystem of Boreal China. Forests, 13(7), 1029. https://doi.org/10.3390/f13071029