
Analysis of Tree Species Suitability for Plantation Forests in Beijing (China) Using an Optimal Random Forest Algorithm



Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Data Preprocessing
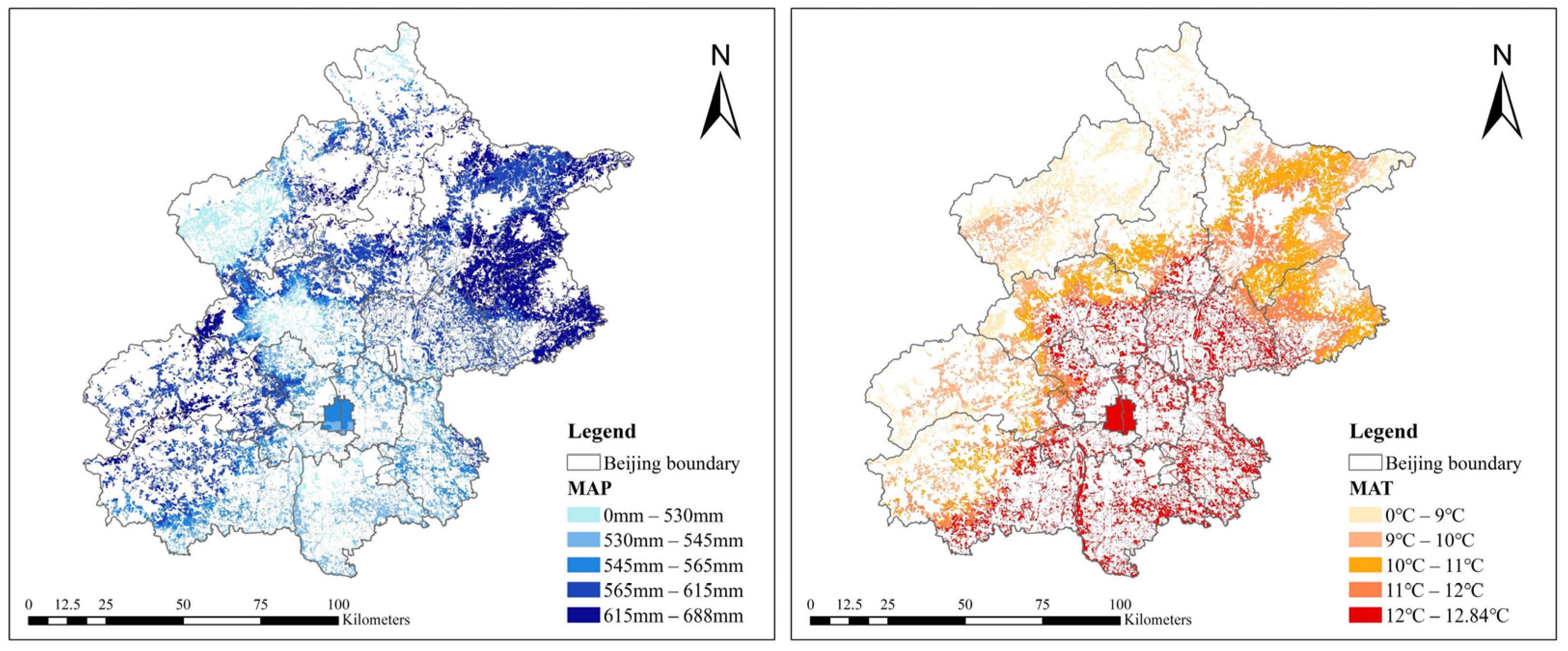
2.2.1. Independent Variables
2.2.2. Dependent Variable—Site Quality Evaluation
2.3. Model
3. Results
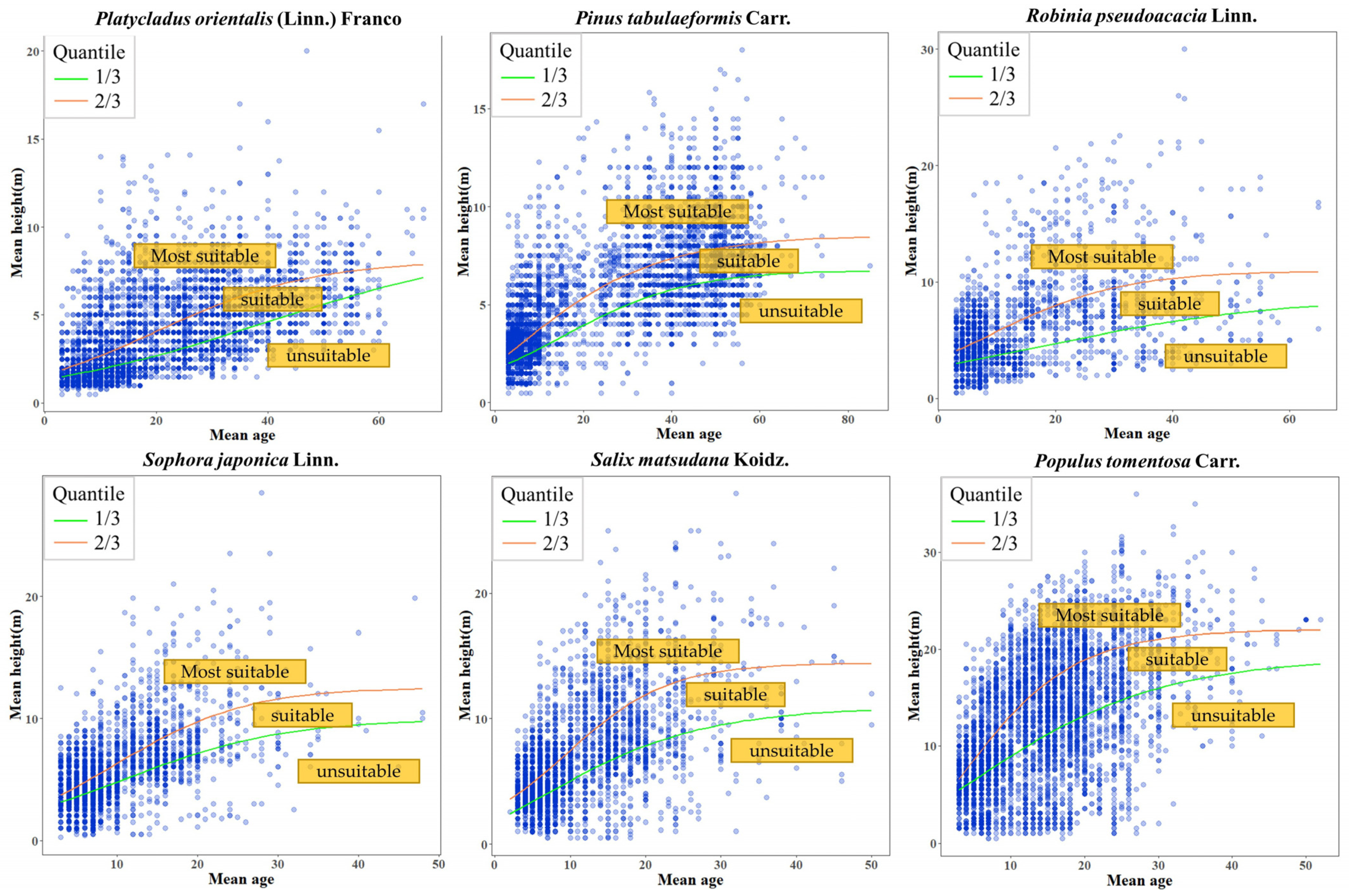
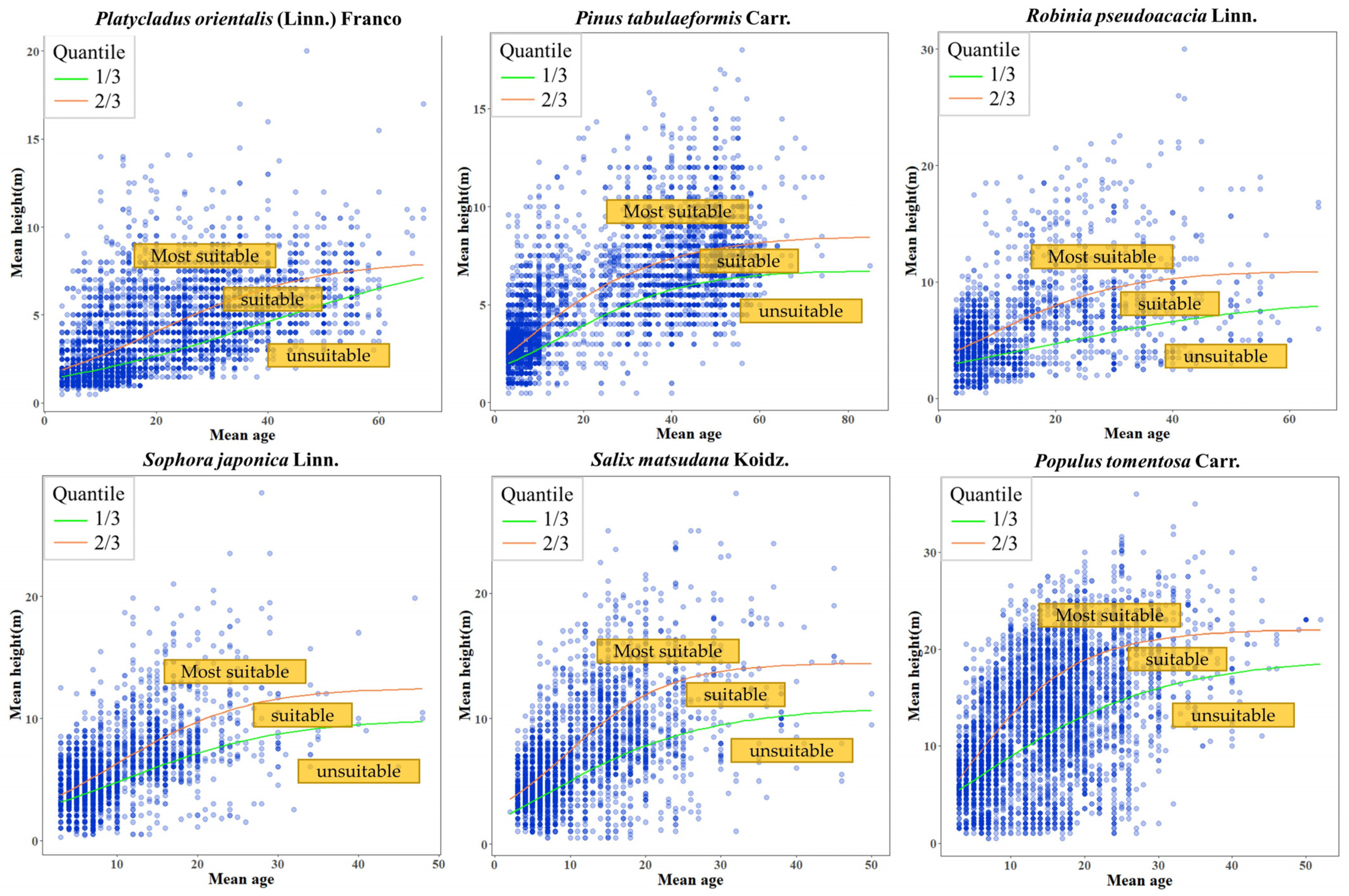
3.1. Site Quality Evaluation
3.2. Model Verification
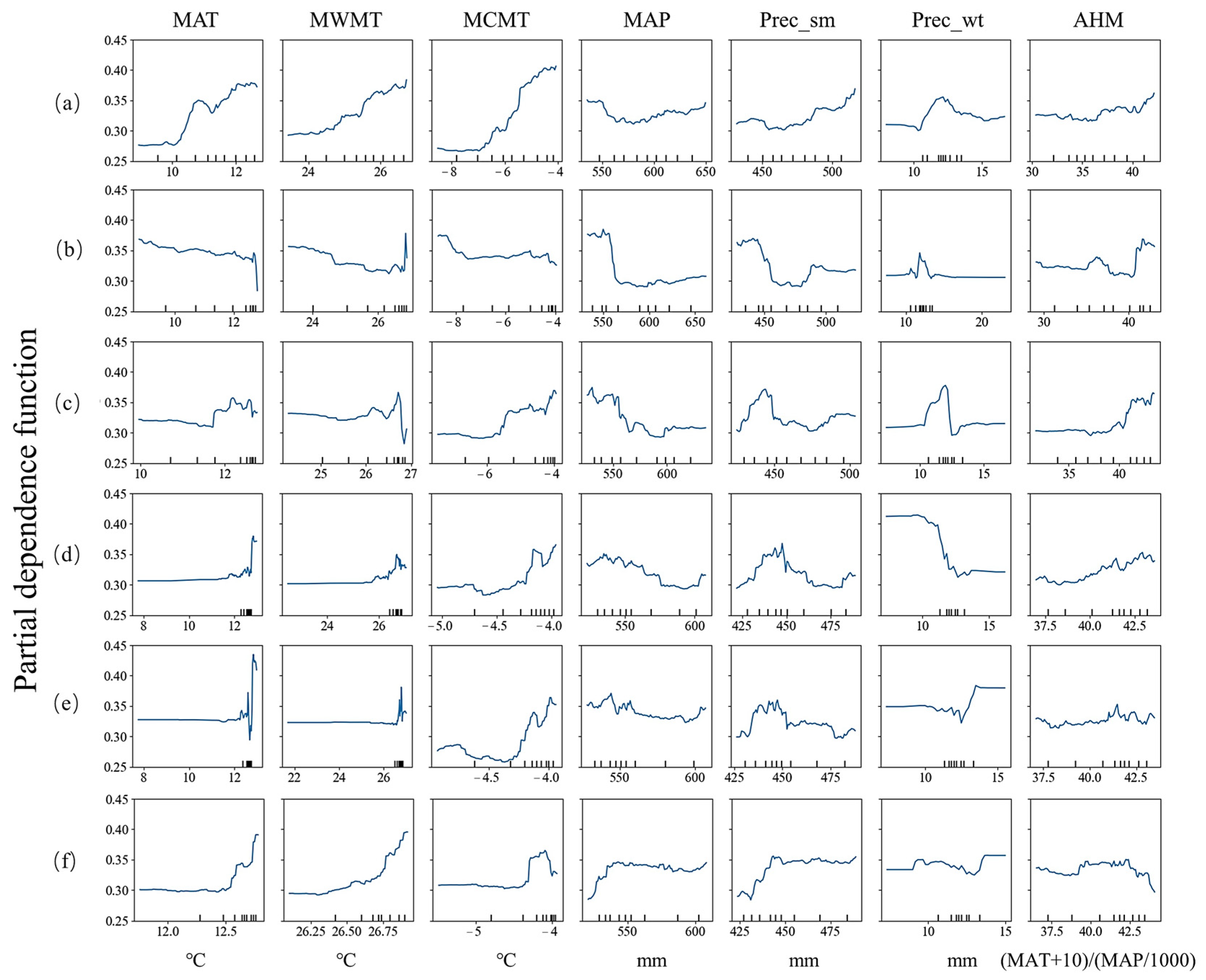
3.3. Relative Importance Analysis and Partial Dependence Plots
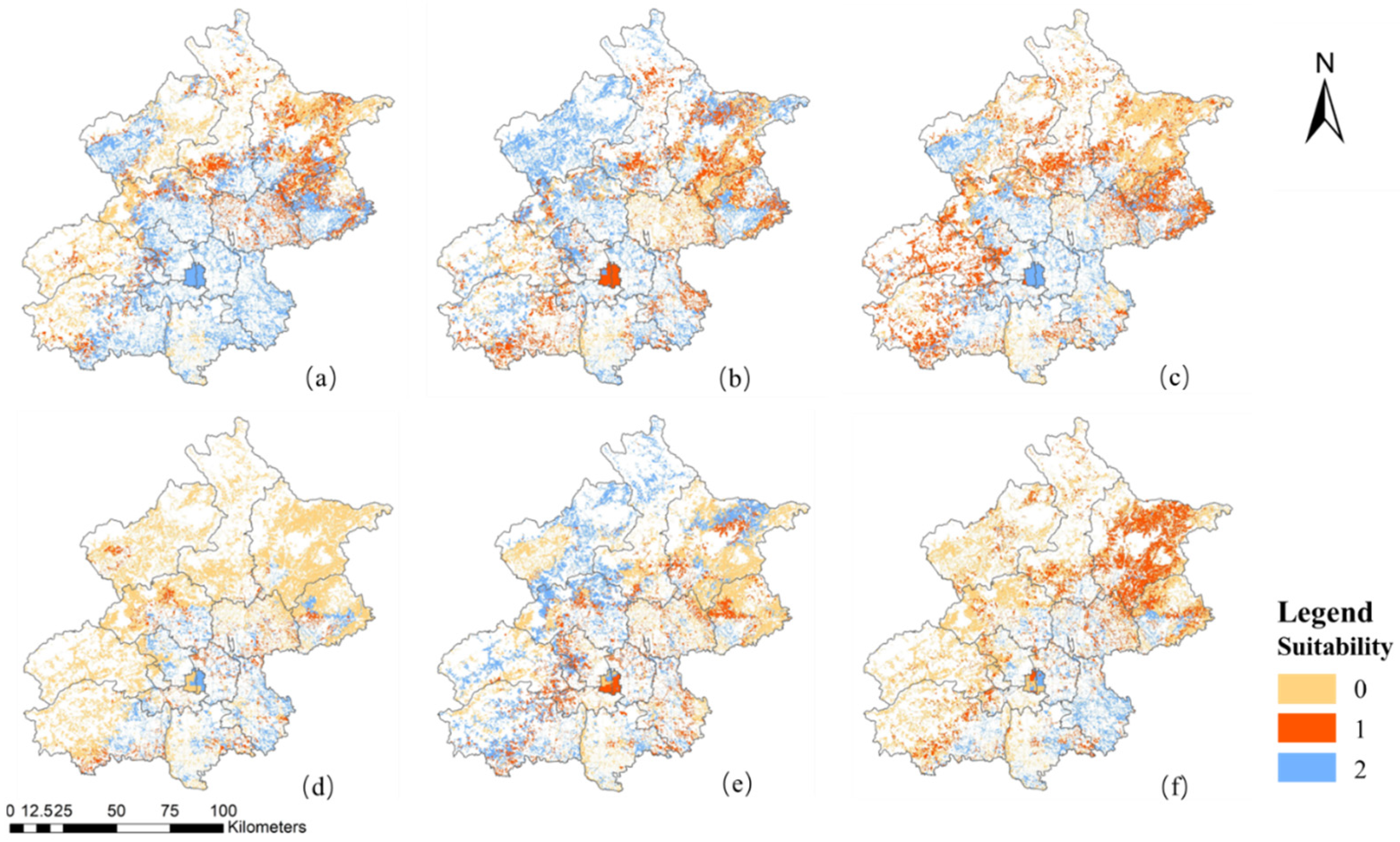
3.4. Spatial Distribution of Tree Species Suitability Zones in Beijing
4. Discussion
4.1. Factors and Their Influence
4.2. Implications for Matching Species with the Site
4.3. Strengths and Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morales-Hidalgo, D.; Oswalt, S.N.; Somanathan, E. Status and trends in global primary forest, protected areas, and areas designated for conservation of biodiversity from the Global Forest Resources Assessment 2015. For. Ecol. Manag. 2015, 352, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Keenan, R.J.; Reams, G.A.; Achard, F.; de Freitas, J.V.; Grainger, A.; Lindquist, E. Dynamics of global forest area: Results from the FAO Global Forest Resources Assessment 2015. For. Ecol. Manag. 2015, 352, 9–20. [Google Scholar] [CrossRef]
- Piao, S.; Huang, M.; Liu, Z.; Wang, X.; Ciais, P.; Canadell, J.G.; Wang, K.; Bastos, A.; Friedlingstein, P.; Houghton, R.A.; et al. Lower land-use emissions responsible for increased net land carbon sink during the slow warming period. Nat. Geosci. 2018, 11, 739–743. [Google Scholar] [CrossRef] [Green Version]
- Aerts, R.; Honnay, O. Forest restoration, biodiversity and ecosystem functioning. BMC Ecol. 2011, 11, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wu, B.; Chen, D.; Qi, Y. Using Machine Learning to Assess Site Suitability for Afforestation with Particular Species. Forests 2019, 10, 739. [Google Scholar] [CrossRef] [Green Version]
- Cuong, N.D.; Volker, M.; Kohl, M. Facilitating objective forest land use decisions by site classification and tree growth modeling: A case study from Vietnam. iForest-Biogeosciences For. 2019, 12, 542–550. [Google Scholar] [CrossRef]
- Mbabazi, J. Ecosystem Goods and Services from Plantation Forests. Int. J. Environ. Stud. 2010, 68, 249–250. [Google Scholar] [CrossRef]
- Barua, S.K.; Lehtonen, P.; Pahkasalo, T. Plantation Vision: Potentials, Challenges and Policy Options for Global Industrial Forest Plantation Development. Int. For. Rev. 2014, 16, 117–127. [Google Scholar] [CrossRef]
- Payn, T.; Carnus, J.M.; Freer-Smith, P.; Kimberley, M.; Kollert, W.; Liu, S.; Orazio, C.; Rodriguez, L.; Silva, L.N.; Wingfield, M.J. Changes in planted forests and future global implications. For. Ecol. Manag. 2015, 352, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Baral, H.; Guariguata, M.R.; Keenan, R.J. A proposed framework for assessing ecosystem goods and services from planted forests. Ecosyst. Serv. 2016, 22, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; Feng, Z.; Song, Y.; Li, M.; Zhang, P. Carbon sequestration potential of forest vegetation in China from 2003 to 2050: Predicting forest vegetation growth based on climate and the environment. J. Clean. Prod. 2020, 252, 119715. [Google Scholar] [CrossRef]
- Schneider, R.; Franceschini, T.; Fortin, M.; Saucier, J.-P. Climate-induced changes in the stem form of 5 North American tree species. For. Ecol. Manag. 2018, 427, 446–455. [Google Scholar] [CrossRef]
- Calama, R.; Conde, M.; de-Dios-García, J.; Madrigal, G.; Vázquez-Piqué, J.; Gordo, F.J.; Pardos, M. Linking climate, annual growth and competition in a Mediterranean forest: Pinus pinea in the Spanish Northern Plateau. Agric. For. Meteorol. 2019, 264, 309–321. [Google Scholar] [CrossRef]
- Zhao, X.; Feng, Z.; Zhou, Y.; Lin, Y. Key Technologies of Forest Resource Examination System Development in China. Engineering 2019, 6, 491–494. [Google Scholar] [CrossRef]
- Eckhart, T.; Pötzelsberger, E.; Koeck, R.; Thom, D.; Lair, G.J.; van Loo, M.; Hasenauer, H. Forest stand productivity derived from site conditions: An assessment of old Douglas-fir stands (Pseudotsuga menziesii (Mirb.) Franco var. menziesii) in Central Europe. Ann. For. Sci. 2019, 76, 19. [Google Scholar] [CrossRef] [Green Version]
- Louw, J.H.; Scholes, M. Forest site classification and evaluation: A South African perspective. For. Ecol. Manag. 2002, 171, 153–168. [Google Scholar] [CrossRef]
- Watt, M.S.; Dash, J.P.; Bhandari, S.; Watt, P. Comparing parametric and non-parametric methods of predicting Site Index for radiata pine using combinations of data derived from environmental surfaces, satellite imagery and airborne laser scanning. For. Ecol. Manag. 2015, 357, 1–9. [Google Scholar] [CrossRef]
- Carmean, W.H. Forest Site Quality Evaluation in The United States. Adv. Agron. 1975, 27, 209–269. [Google Scholar] [CrossRef]
- Schoenholtz, S.H.; Van Miegroet, H.; Burger, J.A. A review of chemical and physical properties as indicators of forest soil quality: Challenges and opportunities. For. Ecol. Manag. 2000, 138, 335–356. [Google Scholar] [CrossRef]
- Fiandino, S.; Plevich, J.; Tarico, J.; Utello, M.; Demaestri, M.; Gyenge, J. Modeling forest site productivity using climate data and topographic imagery in Pinus elliottii plantations of central Argentina. Ann. For. Sci. 2020, 77, 95. [Google Scholar] [CrossRef]
- Barrio-Anta, M.; Castedo-Dorado, F.; Cámara-Obregón, A.; López-Sánchez, C.A. Predicting current and future suitable habitat and productivity for Atlantic populations of maritime pine (Pinus pinaster Aiton) in Spain. Ann. For. Sci. 2020, 77, 41. [Google Scholar] [CrossRef]
- Beaulieu, J.; Raulier, F.; Prégent, G.; Bousquet, J. Predicting site index from climatic, edaphic, and stand structural properties for seven plantation-grown conifer species in Quebec. Can. J. For. Res. 2011, 41, 682–693. [Google Scholar] [CrossRef]
- Curt, T.; Bouchaud, M.; Agrech, G. Predicting site index of Douglas-Fir plantations from ecological variables in the Massif Central area of France. For. Ecol. Manag. 2001, 149, 61–74. [Google Scholar] [CrossRef]
- Sharma, R.P.; Brunner, A.; Eid, T. Site index prediction from site and climate variables for Norway spruce and Scots pine in Norway. Scand. J. For. Res. 2012, 27, 619–636. [Google Scholar] [CrossRef]
- Uzoh, F.C.C.; Oliver, W.W. Individual tree height increment model for managed even-aged stands of ponderosa pine throughout the western United States using linear mixed effects models. For. Ecol. Manag. 2006, 221, 147–154. [Google Scholar] [CrossRef]
- Farrelly, N.; Dhubháin, Á.N.; Nieuwenhuis, M. Sitka spruce site index in response to varying soil moisture and nutrients in three different climate regions in Ireland. For. Ecol. Manag. 2011, 262, 2199–2206. [Google Scholar] [CrossRef]
- Bueis, T.; Bravo, F.; Pando, V.; Turrion, M.B. Relationship between environmental parameters and Pinus sylvestris L. site index in forest plantations in northern Spain acidic plateau. iForest-Biogeosciences For. 2016, 9, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Feng, Z.; Ma, C.; Yang, L. Influencing factors and growth state classification of a natural Metasequoia population. J. For. Res. 2019, 30, 337–345. [Google Scholar] [CrossRef]
- Wang, T.; Wang, G.; Innes, J.L.; Seely, B.; Chen, B. ClimateAP: An application for dynamic local downscaling of historical and future climate data in Asia Pacific. Front. Agric. Sci. Eng. 2017, 4, 448–458. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Duan, A. Approach to Theoretical Growth Equations for Modelling Stands Diameter Structure of Chinese Fir Plantations. Sci. Silvae Sin. 2003, 39, 55–61. [Google Scholar] [CrossRef]
- Koenker, R.; Bassett, G.W. Regression quantiles. Econometrica 1978, 46, 211–244. [Google Scholar] [CrossRef]
- Furno, M.; Vistocco, D. Quantile Regression: Estimation and Simulation; John Wiley & Sons: Hoboken, NJ, USA, 2018; pp. 1–287. [Google Scholar]
- Cade, B.S.; Noon, B.R. A gentle introduction to quantile regression for ecologists. Front. Ecol. Environ. 2003, 1, 412–420. [Google Scholar] [CrossRef]
- Zhang, B.; Sajjad, S.; Chen, K.; Zhou, L.; Zhang, Y.; Yong, K.K.; Sun, Y. Predicting Tree Height-Diameter Relationship from Relative Competition Levels Using Quantile Regression Models for Chinese Fir (Cunninghamia lanceolata) in Fujian Province, China. Forests 2020, 11, 183. [Google Scholar] [CrossRef] [Green Version]
- Weiskittel, A.R.; Crookston, N.L.; Radtke, P.J. Linking climate, gross primary productivity, and site index across forests of the western United States. Can. J. For. Res. 2011, 41, 1710–1721. [Google Scholar] [CrossRef]
- Sun, A.; Lim, E.P. Hierarchical Text Classification and Evaluation. In Proceedings of the 2001 IEEE International Conference on Data Mining, San Jose, CA, USA, 29 November–2 December 2001. [Google Scholar]
- Heung, B.; Bulmer, C.E.; Schmidt, M.G. Predictive soil parent material mapping at a regional-scale: A Random Forest approach. Geoderma 2014, 214–215, 141–154. [Google Scholar] [CrossRef]
- Zhu, Y.; Feng, Z.; Lu, J.; Liu, J. Estimation of Forest Biomass in Beijing (China) Using Multisource Remote Sensing and Forest Inventory Data. Forests 2020, 11, 163. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Uriarte, R.; Bioinformatics, S.A.J.B. Gene selection and classification of microarray data using random forest. BMC Bioinform. 2006, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Bernard, S.; Heutte, L.; Adam, S. Influence of Hyperparameters on Random Forest Accuracy; Springer: Berlin/Heidelberg, Germany, 2009; Volume 5519, pp. 171–180. [Google Scholar]
- Goldberg, D.E. Genetic Algorithm in Search, Optimization, and Machine Learning; Addison Wesley Publishing Company: Boston, MA, USA, 1989. [Google Scholar]
- Bylander, T. Estimating Generalization Error on Two-Class Datasets Using Out-of-Bag Estimates. Mach. Learn. 2002, 48, 287–297. [Google Scholar] [CrossRef]
- Breiman, L. Out-Of-Bag Estimation. 1996. Available online: https://www.stat.berkeley.edu/~breiman/OOBestimation.pdf (accessed on 14 April 2022).
- Jiménez-Valverde, A. Insights into the area under the receiver operating characteristic curve (AUC) as a discrimination measure in species distribution modelling. Glob. Ecol. Biogeogr. 2012, 21, 498–507. [Google Scholar] [CrossRef]
- Ma, W.; Feng, Z.; Cheng, Z.; Chen, S.; Wang, F. Identifying Forest Fire Driving Factors and Related Impacts in China Using Random Forest Algorithm. Forests 2020, 11, 507. [Google Scholar] [CrossRef]
- Friedman, J.H. Greedy function approximation: A gradient boosting machine. Ann. Stat. 2001, 29, 1189–1232. [Google Scholar] [CrossRef]
- Toledo, M.; Poorter, L.; Peña-Claros, M.; Alarcón, A.; Balcázar, J.; Leaño, C.; Licona, J.C.; Llanque, O.; Vroomans, V.; Zuidema, P.; et al. Climate is a stronger driver of tree and forest growth rates than soil and disturbance. J. Ecol. 2011, 99, 254–264. [Google Scholar] [CrossRef]
- Price, D.T.; Cooke, B.J.; Metsaranta, J.M.; Kurz, W.A. If forest dynamics in Canada’s west are driven mainly by competition, why did they change? Half-century evidence says: Climate change. Proc. Natl. Acad. Sci. USA 2015, 112, E4340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jump, A.S.; Mátyás, C.; Peñuelas, J. The altitude-for-latitude disparity in the range retractions of woody species. Trends Ecol. Evol. 2009, 24, 694–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Moreno, J.F.; Mannaerts, C.M.; Jetten, V. Influence of topography on rainfall variability in Santiago Island, Cape Verde. Int. J. Climatol. 2014, 34, 1081–1097. [Google Scholar] [CrossRef]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R.P. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef]
- Babst, F.; Poulter, B.; Trouet, V.; Tan, K.; Neuwirth, B.; Wilson, R.; Carrer, M.; Grabner, M.; Tegel, W.; Levanic, T.; et al. Site- and species-specific responses of forest growth to climate across the European continent. Glob. Ecol. Biogeogr. 2013, 22, 706–717. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, X.; Chhin, S.; Zhang, J.; Duan, A. Disentangling the effects of stand and climatic variables on forest productivity of Chinese fir plantations in subtropical China using a random forest algorithm. Agric. For. Meteorol. 2021, 304–305, 108412. [Google Scholar] [CrossRef]
- Linares, J.C.; Tiscar, P.A. Buffered climate change effects in a Mediterranean pine species: Range limit implications from a tree-ring study. Oecologia 2011, 167, 847–859. [Google Scholar] [CrossRef]
- Sidor, C.G.; Popa, I.; Vlad, R.; Cherubini, P. Different tree-ring responses of Norway spruce to air temperature across an altitudinal gradient in the Eastern Carpathians (Romania). Trees 2015, 29, 985–997. [Google Scholar] [CrossRef]
- Kolář, T.; Čermák, P.; Trnka, M.; Žid, T.; Rybníček, M. Temporal changes in the climate sensitivity of Norway spruce and European beech along an elevation gradient in Central Europe. Agric. For. Meteorol. 2017, 239, 24–33. [Google Scholar] [CrossRef]
- Driscoll, W.W.; Wiles, G.C.; D’Arrigo, R.D.; Wilmking, M. Divergent tree growth response to recent climatic warming, Lake Clark National Park and Preserve, Alaska. Geophys. Res. Lett. 2005, 32, L20703. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Liu, X.; Zhang, Q.; Zeng, X.; Xu, G.; Wu, G.; Wang, W. Species-specific tree growth and intrinsic water-use efficiency of Dahurian larch (Larix gmelinii) and Mongolian pine (Pinus sylvestris var. mongolica) growing in a boreal permafrost region of the Greater Hinggan Mountains, Northeastern China. Agric. For. Meteorol. 2018, 248, 145–155. [Google Scholar] [CrossRef]
- Metzen, D.; Sheridan, G.J.; Benyon, R.G.; Bolstad, P.V.; Griebel, A.; Lane, P.N.J. Spatio-temporal transpiration patterns reflect vegetation structure in complex upland terrain. Sci. Total Environ. 2019, 694, 133551. [Google Scholar] [CrossRef] [PubMed]
- Aerts, R. The advantages of being evergreen. Trends Ecol. Evol. 1995, 10, 402–407. [Google Scholar] [CrossRef]
- Tromp-van Meerveld, H.J.; McDonnell, J.J. On the interrelations between topography, soil depth, soil moisture, transpiration rates and species distribution at the hillslope scale. Adv. Water Resour. 2006, 29, 293–310. [Google Scholar] [CrossRef]
- Burley, S.T.; Harper, K.A.; Lundholm, J.T. Vegetation composition, structure and soil properties across coastal forest–barren ecotones. Plant Ecol. 2010, 211, 279–296. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Species | Sample Size | Elevation (m) | |||
|---|---|---|---|---|---|---|
| Min. | Max. | Mean | Median | |||
| 1 | Platycladus orientalis (Linn.) Franco | 8822 | 0 | 1248 | 252.5 | 219 |
| 2 | Pinus tabulaeformis Carr. | 9689 | 0 | 3501 | 165.7 | 46 |
| 3 | Robinia pseudoacacia Linn. | 3208 | 0 | 1146 | 113.7 | 40 |
| 4 | Sophora japonica Linn. | 4193 | 0 | 824 | 35.3 | 30 |
| 5 | Salix matsudana Koidz. | 4346 | 0 | 980 | 28.6 | 28 |
| 6 | Populus tomentosa Carr. | 5484 | 0 | 750 | 34.9 | 30 |
| Variable Type | Variable Name | Describe | Mean | Median | Min. | Max. |
|---|---|---|---|---|---|---|
| Climatic | MAT | Mean annual temperature (°C) | 11.95 | 12.52 | 5.64 | 12.98 |
| MWMT | Mean warmest month temperature (°C) | 26.11 | 26.62 | 20.58 | 27.1 | |
| MCMT | Mean coldest month temperature (°C) | −4.98 | −4.3 | −12.52 | −3.68 | |
| MAP | Mean annual precipitation (mm) | 575.6 | 567.2 | 472.2 | 764.6 | |
| Prec_sm | Summer precipitation (mm) | 462.4 | 456.2 | 366 | 605.3 | |
| Prec_wt | Winter precipitation (mm) | 12.1 | 12 | 7.3 | 23.7 | |
| AHM | Annual heat:moisture index ((MAT + 10)/(MAP/1000)) | 38.8 | 39.7 | 21.2 | 48.94 | |
| Topographic | Landform | Landform | / | / | / | / |
| Slope-A | Slope aspect | / | / | / | / | |
| Slope-D | Slope degree (°) | 8.06 | 0 | 0 | 60 | |
| Slope-P | Slope position | / | / | / | / | |
| Edaphic | Soil-TP | Soil type | / | / | / | / |
| Soil-TN | Soil thickness (cm) | 47.71 | 60 | 0 | 430 | |
| BRP | Bare rock percentage (%) | 0.57 | 0 | 0 | 90 | |
| HLT | Humus layer thickness (cm) | 0.20 | 0 | 0 | 60 | |
| GWL | Groundwater level (m) | 17.44 | 0 | 0 | 2013 |
| Variable Name | Categories (Number) |
|---|---|
| Slope aspect | North (338°~22°) (1795), Northeast (23°~67°) (1481), East (68°~112°) (1385), Southeast (113°~157°) (1468), South (158°~202°) (1795), Southwest (203°~247°) (1277), West (248°~292°) (1046), Northwest (293°~337°) (1260), No slope direction (24,235) |
| Slope position | Ridge (35), Upslope (1195), Mid (634), Downslope (1570), Valley (164), Flat (24,309), Whole slope (7835) |
| Soil type | Alluvial soil (18,617), Drab soil (13,808), Brown soil (475), Wind-blown soil (2842) |
| Landform | Plain (23,077), Low mountain (12,201), Medium mountain (464) |
| No | Model | Model Expression |
|---|---|---|
| 1 | Logistic (Verhulst, 1838) | h = A/(1 + B × exp(C × t)) |
| 2 | Gompertz (Gompertz, 1825) | h = A × exp(−B × exp(−C × t)) |
| 3 | Richards (Richards, 1959) | h = A × (1 − exp(−C × t))^B |
| 4 | Korf (Korf, 1939) | h = A × exp(−B × t^(−C)) |
| 5 | Mitscherlich (Mitscherlich, 1919) | h = A × (1 − exp(−C × t)) |
| Species | 1/3 Quantile Regression Line | 2/3 Quantile Regression Line | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Model | A | B | C | AIC | Model | A | B | C | AIC | |
| Platycladus orientalis (Linn.) Franco | 1 | 9.07 | 5.77 | −0.04 | 29,267.59 | 1 | 8.275 | 4.135 | −0.069 | 32,881.02 |
| Pinus tabulaeformis Carr. | 1 | 6.808 | 2.967 | −0.070 | 32,963.45 | 2 | 8.628 | 1.464 | 0.056 | 36,639.91 |
| Robinia pseudoacacia Linn. | 1 | 8.719 | 2.206 | −0.048 | 15,158.21 | 1 | 10.955 | 2.191 | −0.089 | 16,228.38 |
| Sophora japonica Linn. | 1 | 9.11 | 3.33 | −0.12 | 17,361.99 | 1 | 12.543 | 3.448 | −0.123 | 18,686.13 |
| Salix matsudana Koidz. | 2 | 10.99 | 1.81 | 0.08 | 20,781.45 | 1 | 14.432 | 4.155 | −0.149 | 18,686.13 |
| Populus tomentosa Carr. | 2 | 19.21 | 1.56 | 0.074 | 34,250.48 | 2 | 22.025 | 1.742 | 0.121 | 34,080.18 |
| Species | n_estimators | max_feature | min_leaf | RF AUC | ORF AUC |
|---|---|---|---|---|---|
| Platycladus orientalis (Linn.) Franco | 600 | 3 | 2 | 0.7920 | 0.8042 |
| Pinus tabulaeformis Carr. | 300 | 2 | 2 | 0.8073 | 0.8257 |
| Robinia pseudoacacia Linn. | 70 | 6 | 3 | 0.8231 | 0.8399 |
| Sophora japonica Linn. | 420 | 2 | 2 | 0.7348 | 0.7626 |
| Salix matsudana Koidz. | 440 | 4 | 2 | 0.7531 | 0.7642 |
| Populus tomentosa Carr. | 510 | 1 | 2 | 0.7424 | 0.7518 |
| Species | Climatic | Topographic | Edaphic |
|---|---|---|---|
| Platycladus orientalis (Linn.) Franco | 73.16% | 13.26% | 13.58% |
| Pinus tabulaeformis Carr. | 81.65% | 7.99% | 10.36% |
| Robinia pseudoacacia Linn. | 72.57% | 16.39% | 11.04% |
| Sophora japonica Linn. | 91.87% | 1.51% | 7.40% |
| Salix matsudana Koidz. | 93.32% | 0.75% | 5.93% |
| Populus tomentosa Carr. | 86.90% | 2.09% | 11.01% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Feng, Z.; Ma, W. Analysis of Tree Species Suitability for Plantation Forests in Beijing (China) Using an Optimal Random Forest Algorithm. Forests 2022, 13, 820. https://doi.org/10.3390/f13060820
Wang Y, Feng Z, Ma W. Analysis of Tree Species Suitability for Plantation Forests in Beijing (China) Using an Optimal Random Forest Algorithm. Forests. 2022; 13(6):820. https://doi.org/10.3390/f13060820
Chicago/Turabian StyleWang, Yuan, Zhongke Feng, and Wenyuan Ma. 2022. "Analysis of Tree Species Suitability for Plantation Forests in Beijing (China) Using an Optimal Random Forest Algorithm" Forests 13, no. 6: 820. https://doi.org/10.3390/f13060820
APA StyleWang, Y., Feng, Z., & Ma, W. (2022). Analysis of Tree Species Suitability for Plantation Forests in Beijing (China) Using an Optimal Random Forest Algorithm. Forests, 13(6), 820. https://doi.org/10.3390/f13060820