Abstract
Natural regeneration in forest management, which relies on artificial planting, is considered a desirable alternative to reforestation. However, there are large uncertainties regarding the natural regeneration processes, such as seed production, seed dispersal, and seedling establishment. Among these processes, seed dispersal by wind must be modeled accurately to minimize the risks of natural regeneration. This study aimed to (1) review the main mechanisms of seed dispersal models, their characteristics, and their applications and (2) suggest prospects for seed dispersal models to increase the predictability of natural regeneration. With improving computing and observation systems, the modeling technique for seed dispersal by wind has continued to progress steadily from a simple empirical model to the Eulerian-Lagrangian model. Mechanistic modeling approaches with a dispersal kernel have been widely used and have attempted to be directly incorporated into spatial models. Despite the rapid development of various wind-dispersal models, only a few studies have considered their application in natural regeneration. We identified the potential attributes of seed dispersal modeling that cause high uncertainties and poor simulation results in natural regeneration scenarios: topography, pre-processing of wind data, and various inherent complexities in seed dispersal processes. We suggest that seed dispersal models can be further improved by incorporating (1) seed abscission mechanisms by wind, (2) spatiotemporally complex wind environments, (3) collisions with the canopy or ground during seed flight, and (4) secondary dispersal, long-distance dispersal, and seed predation. Interdisciplinary research linking climatology, biophysics, and forestry would help improve the prediction of seed dispersal and its impact on natural regeneration.
1. Introduction
Renewing a forest stand through natural regeneration has various economic, environmental, and ecological advantages over artificial regeneration [1,2,3]. However, it also has difficulties and risks in practice because natural regeneration is largely dependent on highly uncertain interactions between biological and environmental factors [4,5]. Specifically, seed dispersal determines the distribution of seeds and trees during the process of natural regeneration and is influenced by both biological (e.g., wing loading of seeds, seed yield, and distance between seeds and the ground) and environmental factors (e.g., wind environment, topography, and climate), which increase the overall uncertainty.
Therefore, seed dispersal models can be useful for simulating seed dispersal and its impact on natural regeneration. Because of the highly stochastic nature of seed dispersal, it is important to better understand the mechanisms of different seed dispersal models, identify the sources of uncertainties, and validate model simulations by comparing them with observations. In this review, we identify the key processes and uncertainties in seed dispersal and natural regeneration. We further investigate the mechanisms of seed dispersal models and the challenges in predicting seed dispersal in natural regeneration scenarios. Finally, we discuss the future directions for developing seed dispersal models for the best practice of natural regeneration.
2. Natural Regeneration and Seed Dispersal
Natural regeneration is defined as the development of a new stand from seeds or sprouts sourced from pre-harvesting or adjacent stands after a portion of the canopy is harvested by clear-cutting, partial-cutting, selective-cutting, or seed-tree methods [6]. The key principle of natural regeneration is to imitate natural disturbance regimes with various intensities, such as fires, windthrows, and flooding, which create niches with favorable environmental conditions (e.g., light, nutrients, and lack of competitors) for newly established seedlings or sprouts [7,8]. Natural regeneration is expected to promote vigorous belowground development in seedlings [5] and lower costs by reducing the need for the nurseries, planting, and seeding procedures required for artificial regeneration [2,9].
Natural regeneration processes can be divided into three stages: seed production, seed dispersal, and seedling establishment (Figure 1) [4,6,7]. These are mainly affected by the microclimate (e.g., wind, temperature, rainfall, and light) and stand structure [4,5]. In the seed production phase, seeds are produced following the flowering, pollination, and the maturation of fertilized seeds in the strobilus or ovary. Seed production results from the allocation of primary production and is influenced by the microclimate and physiology of flowering and fruiting.
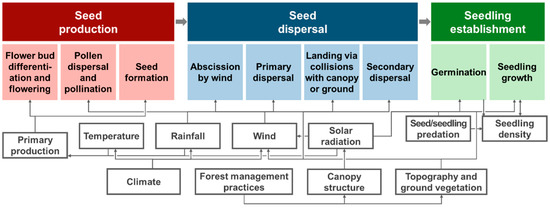
Figure 1.
Natural regeneration processes from seed production to seedling establishment of tall forest trees depend on wind-dispersal of seeds. Microclimate (e.g., temperature, rainfall, wind, and solar radiation) and stand structure (e.g., canopy, topography, and ground vegetation) interactively regulate the regeneration processes. Human-induced transformations due to climate change and forest management practices could alter microclimate and stand structure, respectively.
During the seed dispersal phase, the seed is carried by the wind because of its winged or pappus assemblages [10]. The morphological unit of dispersal (i.e., diaspores) differs between species. Gymnosperm seeds from genera such as Pinus, Abies, and Picea have wings and are released directly into the atmosphere. In contrast, the dispersal unit of angiosperms such as those from the genera Acer, Fraxinus, and Alnus is a winged fruit, generally called a samara [11], which carries the seeds. Additionally, for angiosperms with capsule-type fruits such as Populus, Salix, and Catalpa, seeds are released from dehisced fruits and dispersed with the aid of a pappus [12]. Morphologically, the dispersal of angiosperms should be distinguished from that of winged gymnosperm seeds. Nevertheless, we refer to the release of all these morphologically different bodies as seed dispersal in practice. They may move by parachuting, gliding, rocking, or rotating (i.e., spinning) depending on the species [10] and reach a steady state of falling at a certain terminal velocity [13]. The movement of seeds is regulated by various aerodynamic forces, such as gravity, drag, and lift, owing to the interactions between the aerodynamic structures of the seed and air [10,14]. Finally, the seed lands on the ground directly or after colliding with parts of the tree’s crown during its descent. Seeds that arrive on the ground may be transported further by wind, gravity, or water flow greater than the frictional force of the ground. This is called secondary dispersal [4,15,16] and is distinct from primary dispersal following the initial release of seeds from the plant into the air. Wind, the primary factor controlling seed dispersal, is regulated by the climate, topography, and stand structure. Animals also aid seed dispersal, and animal-mediated seed dispersal is an effective and significant means for plant migration in various ecosystems [17,18,19]. Nevertheless, most silviculture species in temperate forests (e.g., pine, larch, spruce, fir, and ash) have winged seeds that have evolved to rely on seed dispersal by wind. During the seedling establishment phase, seeds break dormancy, if any, and germinate depending on environmental conditions, such as light quantity, light quality, temperature, and water availability, and develop into seedlings.
The results of natural regeneration are highly uncertain because they depend on processes, such as seed production, dispersal, and germination, which are influenced by the interaction between biological and environmental factors. Natural regeneration can fail when seeds disperse to locations and conditions that are not feasible for germination and growth [4,5,8,20]. The success of natural regeneration practices requires site-specific knowledge and experience to determine the optimal size, design, and timing of harvesting [4,21,22] to ensure that environmental conditions are feasible for regeneration processes. Otherwise, a regenerating stand may develop undesirably in terms of species composition, density, or growing stock. Natural regeneration has been widely applied in North America and Europe, where modern silviculture skills have been used for over a century, but not in countries where these skills have not yet been widely adopted.
Natural regeneration should be predictable to provide a viable alternative and replacement for some areas now managed with artificial regeneration. However, there are no models that can simulate the entire process of natural regeneration from seed production to seed dispersal and seedling establishment. Few studies have reported empirical or geospatial models for seedling establishment [4,21,22]; however, the studies using a mechanical approach for seed dispersal in the context of natural regeneration are scarce.
Modeling seed dispersal, which mainly regulates seedling establishment, could effectively minimize the uncertainty of natural regeneration. Seed dispersal models can support science-based programs by optimizing the design of natural regeneration, such as (1) displacement, size, and shape of harvesting patches or (2) displacement and the number of remaining seed trees, based on predicting spatial patterns and the effective range of seed dispersal from site information (e.g., stand structure, micro-climate, properties of seeds of a particular tree species, and other factors). In the following section, we review seed dispersal models in more detail.
3. Key Mechanisms of Seed Dispersal Models
Among the seed dispersal processes, wind-dispersal is the most studied and modeled [23]. Therefore, several empirical and mechanistic models have been developed to simulate windborne seed dispersal. Additionally, highly complex models have been developed more recently due to improvements in observation technology and computing performance that enabled more advanced physical calculations (Table 1). Here, we review the key mechanisms of seed dispersal models by wind and their application to natural regeneration practices in forest management.

Table 1.
A brief description of major seed dispersal models.
3.1. Simple Empirical and Mechanistic Dispersal Models
The most widely used empirical models to describe seed dispersal are the inverse power law model [24] and the negative exponential model [26,45]. Despite their simplicity, they effectively describe the asymptotic distribution of seeds (as well as spores or pollen) along the course between the release and landing points. The inverse power law (Equation (1)) and negative exponential (Equation (2)) models are described as follows:
where s is the distance from the source; y is the probability density associated with dispersal; and a and b are constants.
The negative exponential model can be converted into a log-linear model through log transformation. The advantage of this transformation is that the y value can be estimated when s approaches 0. Therefore, the log-linear model is preferred over the negative exponential model, although both models are similarly accurate [24,46].
With the development of mathematical theories, empirical models for trees have been developed using log-normal, 2Dt (bivariate Student’s t distribution), and two-parameter Weibull functions [25,47,48]. These functions are better descriptors of dispersal curves than negative exponential models. In particular, 2Dt can better illustrate long-distance dispersal (LDD) than classic empirical models. Skarpaas et al. [49] confirmed that the 2Dt model performed realistic and flexible fitting for the dispersal of leafless hawk’s beard (Crepis praemorsa) in both field release experiments and trap monitoring. Mechanistic models have also been applied to simulate seed dispersal by animals and humans [50,51]. However, practitioners still disagree on the most appropriate functional form of the dispersal curve [25,48,52,53].
The major advantage of the empirical models is their simplicity. They require only two parameters to be used, which are easy to obtain from widely available in-situ measurements. Empirical models are effective in approximating the range of seed dispersal. However, they reproduce observations with measured parameters in a specific environment rather than imitating physical mechanisms. Thus, applying the empirical models to different environments where parameter measurements are limited is challenging. In this context, mechanistic models have better universality than empirical models because they imitate physical mechanisms.
Empirical modeling makes it easier to calibrate dispersal kernels than mechanistic modeling. Mechanistic models include a realistic representation of processes such as seed abscission and turbulence [54]. Several mechanistic dispersal models are based on a simple ballistic equation that predicts the distance from the point of particle release to the deposition location [55,56] (Equation (3)).
where xd is the deposition location, h is the release height of the particle, u is the horizontal wind velocity at which the particle was released, and vt is the particle’s terminal velocity.
However, these early mechanical models could not explain the variation in dispersal with wind speed and vertical and horizontal directions [55,56,57]. Moreover, it is difficult to quantify the LDD of seeds based on the average dispersal distance derived from a simple mechanistic dispersal model [55].
Similar to empirical models, mechanistic wind-dispersal models have improved and expanded. Mechanistic wind-dispersal models have been used to identify the variation in dispersal with the interaction between external factors and plants [44,57,58]. Using mechanistic model calculations, Wright et al. [44] evaluated the spatial pattern of dispersed seeds for 16 factorial combinations of four traits for two contrasting tropical tree species. They showed that the spatial pattern of dispersed seeds was affected by a combination of seed terminal velocity, seed release height, seed release in response to sensible heat flux, and seasonal differences in wind direction.
Few studies have used empirical or mechanistic seed dispersal models for trees [49,59,60,61,62]. For example, negative exponential regression using two dispersal kernels was used to estimate the relative seed density of birch clear-cut within a specific distance from the trees in Sweden [61]. Axer et al. [59] applied a log-normal model to predict oak regeneration density in a natural regeneration plot.
3.2. Eulerian and Lagrangian Modeling Approach
Based on the theory of atmospheric diffusion, the Eulerian modeling approach can describe the random motion of particles around the mean eddy. In principle, particle motion can be characterized by an uncorrelated, homogeneous random motion (diffusion) around deterministic drift (advection), and is therefore expressed as an advection-diffusion equation [63,64]. According to the gradient diffusion theory, the mass-conservation equation, which is the key equation of Eulerian modeling, is expressed as
where Kjj is the eddy diffusivity, is the wind velocity, and S is the source or sink distribution at location x = (x, y, z) at time t.
The Gaussian plume model is a representative Eulerian model developed based on Equation (4) and is a simplified model of the effects of gravity, deposition, and spatiotemporal fluctuations of particle motion (Equation (5)). This model was designed to increase the model accuracy of seed dispersal by the wind to reflect the random motion of seeds according to the airflow [65,66]. Applying the Gaussian plume model, assuming the flow of ux and uy velocities in the x-and y-directions (Kxx, Kyy, and Kzz are constants), the average concentration of the particles in the airflow can be explained as follows (Equation (5)):
A common problem with seed dispersal models based on the Eulerian modeling approach is that adding more variables to the equation for improvement is challenging [31,37]. For example, the canopy affects actual seed dispersal, but mathematically solving an equation that reflects this effect is either impossible or requires overgeneralization. Therefore, in general, the effect of canopy structure on seed dispersal was not considered in the Eulerian modeling approach [31,34,67]. While Eulerian methods focus on population-level patterns, Lagrangian approaches consider the movements of individual seeds. The simplest method for obtaining a straight-line trajectory of airborne particles was devised to calculate the landing point based on fixed horizontal and vertical wind speeds [29,68]. This trajectory simulation was performed in the following steps: (1) dividing time into short individual steps, (2) considering specific airflow conditions at each time step, and (3) synthesizing the estimation of particle dispersal at each time step [32,69]. In many previous studies, the Lagrangian stochastic (LS) turbulence model has been proposed to represent transient airflow more realistically [35,68,69,70]. This model simulates the airflow by estimating the acceleration of the air compartments under atmospheric conditions [35,69], and simulate seed dispersal more accurately than other types of wind-dispersal models [32,68].
Over the last two decades, the LS turbulence model has been widely used to develop a wind-dispersal model for seeds or pollen by (1) considering wind flow in three dimensions and local changes in ground surface roughness [69]; (2) describing particle dispersal from forest canopies [70], fields, and grasslands [35,68]; and (3) reflecting the change in the trajectory of particles as they descend from one air stream to another due to gravity [35,71].
The seed acceleration can be calculated from the drag forces and integrated along the trajectory. This principle can be realized by coupling Eulerian flow statistics to the Lagrangian description of seed motion, resulting in the Coupled Eulerian model (CELC) [70,72]. The dispersal kernel of the CELC, which describes seed dispersal, is the probability density function (PDF). The CELC has been used to analyze seed dispersal patterns, such as LDD and secondary dispersal, and to understand ecological phenomena, such as migration and succession [40,41,42,43,44,73,74]. However, CELC or Eulerian and Lagrangian modeling approaches have not yet been used to inform natural regeneration.
3.3. Long-Distance Dispersal
By accumulating empirical data and developing modeling techniques, models can be used to predict the average seed dispersal distance. However, the tail of the dispersal curve in the model results tends to be underestimated [17,51,75,76,77]. For instance, Nathan et al. [70] modeled the seed dispersal of five tree species and showed that 1–5% of fallen seeds traveled an extremely long distance (~10 km), which was much farther than the commonly conceived distance (<100 m). This is called LDD and is an important ecological event (resource use, species coexistence, and large-scale metapopulation dynamics) as well as in the context of evolution (gene flow, genetic structure, and species diversity) [78,79,80,81,82]. Seed dispersal should be distinguished between short-distance dispersal and LDD modeling because their main mechanisms are different [39]. Short-distance dispersal is affected by fall height, horizontal wind speed, and terminal velocity, whereas LDD dispersal is mainly influenced by gusts and updrafts [39]. Over the past two decades, one of the primary research topics on seed dispersal models has been improved LDD methodology. This is because of the practical requirements of the associated research fields, and a scientific objective to overcome the inherent difficulties and complexity of LDD simulations.
Nathan et al. [83] classified studies related to LDD into the following four types based on the methodologies followed. (1) The biogeographic approach. Through observation of LDD, the phenomenon is identified. It also contributes to data construction for the advancement of dispersal algorithms. (2) The migration/redistribution approach. The pattern, rate, and extent of animal and plant reproduction are quantified by tracking the migration/redistribution of an individual or population. This is typically divided into two sub-groups: Eulerian and Lagrangian modeling. (3) The genetic approach. The genetic marker methodology that can distinguish the genetic composition and characteristics of plants and animals is used to observe, identify, and verify LDD. (4) The model approach. A mathematical modeling methodology is used to describe and implement LDD.
Although insight into and understanding of LDD have been greatly improved as each methodology has been advanced and linked, human activities cause unpredictable LDD of seeds, making overall interpretation and prediction more difficult [83].
3.4. Challenges in Seed Dispersal Prediction for Natural Regeneration Using the WINDISPER
As a case study, we tested a mechanistic model to simulate seed dispersal in a naturally regenerating Japanese larch (Larix kaempferi) stand in South Korea to evaluate model performance and address potential improvements for the effective use of the model in the context of natural regeneration (see details of materials, methods, and results in Appendix A). In short, the WINDISPER model [29] poorly simulated seed dispersal compared with the observed seed dispersal data from seed traps (Figure 2 and Figure A1). The coefficient of determination (R2) for the simple linear regressions between the spatial patterns of predicted and observed seed dispersal was up to 0.357, but generally low (<0. 1), or the relationship was occasionally insignificant, depending on the natural regeneration patch and wind speed threshold (Table A2).
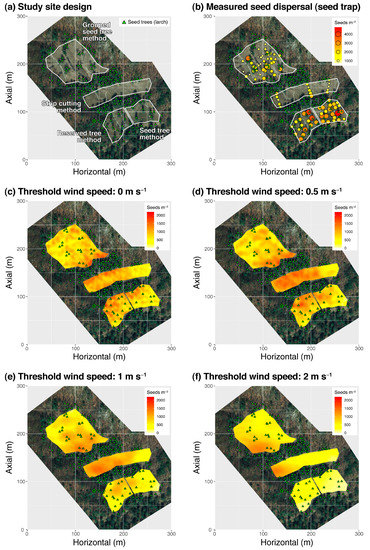
Figure 2.
Distributions of natural regeneration patches and seed trees (a), measured seed dispersal using seed traps (b) and predicted seed dispersal using wind flow dataset filtered by different wind speed thresholds (c–f). Third, various inherent complexities contribute to errors in seed dispersal predictions.
Several challenges in predicting seed dispersal for natural regeneration design using the WINDISPER remain. First, the effects of topography on seed dispersal were not included in the algorithms of current seed dispersal models. Forests in South Korea generally develop on mountainous, complex terrains with relatively small sedimentary flatlands. The Korean Peninsula consists mainly of land with slopes from 10–25° (44%) or above 25° (20%), indicating a much steeper topography than other East Asian regions, with Taiwan being the exception [84]. Depending on the slope direction and steepness, seed flight may be longer or shorter than the estimated flight distance on flat terrain. However, most previous seed dispersal models have disregarded flight distance extension or reduction by slope, resulting in serious errors in seed dispersal prediction in steep terrains. Recently, a few modeling studies have considered the effects of terrain on seed dispersal [74,85]. Trakhtenbrot et al. [74] developed a seed dispersal model combining Eulerian wind statistics and Lagrangian trajectory algorithms to consider topographic effects on the wind environment around the canopy and the landing location of the seed. They predicted the different seed dispersal patterns between the hillcrest and bottom of a mountainous landscape.
The second challenge relates to how pre-treatment wind environment data produced different outcomes of seed dispersal simulations using the WINDISPER model. There is no concrete definition of “seed dispersal-effective wind flow.” Although the presence of a wind speed threshold at which seeds begin to abscise from a seed tree and flight might be assumed, a portion of seeds can fall to the ground vertically under calm wind conditions, which may be lower than the assumed wind speed threshold. Wind flows could be heterogeneous between the shapes of natural regeneration patches, depending on horizontal distances from shelterbelts or seed trees, and vertical heights from the ground or canopy. Although the WINDISPER model presents algorithms that model a vertical profile of wind speed from the wind speed at a certain height, the corrected wind speed may not be sufficient to represent the complexity of wind flow in the space of natural regeneration patches. In summary, these uncertainties in wind flow data are a barrier to accurate simulation of seed dispersal.
Third, various inherent complexities contribute to errors in seed dispersal predictions. The height of the seed trees and the seed yield are not homogeneous. Seed flight ability, mainly represented by terminal velocity, could have intraspecific variations [86]. The WINDISPER model addresses this issue by stochastically generating the height of seed trees, seed production, and terminal velocity of seeds from their mean and standard deviation. However, this model cannot reproduce the fine-scale spatial patterns of actual seed dispersal in natural regeneration patches. Moreover, unpredictable random events affecting flowing seeds, such as changes in wind flow, sudden turbulence, or collisions with remnant trees, make it difficult to predict the seed dispersal trajectory accurately.
4. Prospects for Seed Dispersal Models Applicable to Natural Regeneration
We discovered four complexities resulting in unpredictable patterns of seed dispersal in a naturally regenerated forest, especially on mountainous terrain (Figure 3). The following section explores the challenges and future prospects for developing seed dispersal modeling to a level sufficient to support the field of natural regeneration.
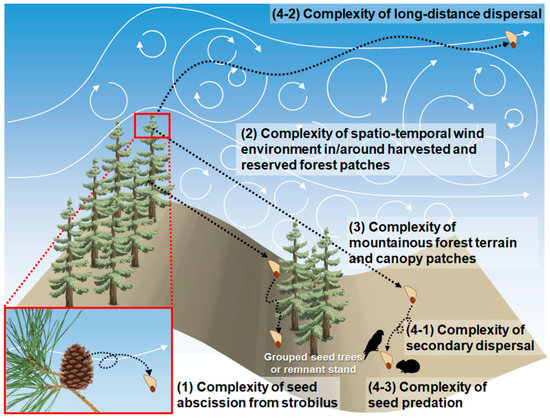
Figure 3.
Challenges in seed dispersal modeling for natural regeneration estimation, especially in managed forest stands on mountainous terrain.
4.1. Seed Abscission Mechanisms by Wind
Current seed dispersal models are primarily based on the unrealistic assumption that seeds are released evenly or randomly over a period set by the user. However, seed abscission from the strobilus or twig could be affected by various factors such as wind force, the openness of the strobilus, and the development of the abscission layer between the twig and samara. To make this assumption more realistic, Pazos et al. [87] hypothesized two mechanisms that determine the wind speed threshold for seed abscission: (1) the maximum deflection angle at which seeds can escape throughout the acute angle of gaps in the strobilus or dehisced fruit, and (2) the material fatigue of seed attachment tissue through cumulative stress by the wind.
If seed abscission is initiated at wind speeds above a certain level, the effective wind environment involved in seed dispersal is much more intensive than the ambient wind environment. For example, the mean wind speed and direction differed depending on the wind speed threshold adopted in the case study (Table A1). Consequently, more seeds can be dispersed over longer distances than the general seed dispersal models predict [88]. Therefore, the stochasticity of seed abscission by wind speed must be developed and applied to an advanced seed dispersal model to support natural regeneration practices. It is necessary to secure experimental and field data to investigate the effects of wind speed on seed abscission [54,89]. Solar radiation and atmospheric temperature also affect seed dispersal, but they are not directly related to wind. They are usually ignored in seed dispersal models, even though they are correlated with the seed dispersal period and mass [90,91].
In addition, the microscale wind environment near a strobilus is complex [92]. Moreover, the stochastics of seed abscission may be heterogeneous, depending on the species-specific morphological structure of the strobilus. The scales of the strobilus open and close when they are dry and wet, respectively [93,94]; therefore, the degree of scale opening, which responds to the lapse of time after rain and air humidity, affects the probability of seeds being exposed to and released by the wind. A mechanistic understanding and experiments on seed abscission timing are required [95].
Most seed dispersal models have been developed to explain the movement of seeds carried by airflow or falling patterns of seeds by distance from seed trees. However, some seeds could land below the trees due to gravity or morphological characteristics. Attempts to distinguish this phenomenon in the modeling approach from general dispersal by wind have not been made. Similarly, research on the effects of rain or raindrops on seed dispersal, such as wet fall by raindrops or reduction in dispersal distance owing to high humidity, is insufficient [35].
4.2. Primary Dispersal by Spatio-Temporally Complex Wind Environment
Silviculture practices for natural regeneration create a land mosaic of harvesting patches for newly regenerated seedlings, and remnant patches for seeded trees, resulting in a heterogeneous canopy structure. Various eddies generate extremely complicated three-dimensional (3D) distributions of longitudinal, latitudinal, and vertical wind speeds in managed stands. For example, remnant patches distort wind flow, resulting in unpredictable eddies [96,97,98,99].
Therefore, the wind environment of a natural regeneration stand is complex and depend on the interior or exterior of the harvesting patch, distance from the harvesting patch boundary or shelterbelt, height from the ground, and direction of the slope. Its characteristics also depend on the outside and distance from the edge of the forest. In addition to the spatial complexity of the wind environment, temporal complexity is also expected to be significant. The wind environment changes with seasonal changes in the prevailing wind, weather phenomena such as typhoons, and the leaf area index (LAI) of the forest canopy according to leaf unfolding and senescence [100,101].
Seed dispersal prediction using models is highly sensitive to the wind environment [29,39] because wind determines the seeds’ flight. The spatiotemporal complexity of the wind environment in a natural regeneration stand is a major source of uncertainty. For example, in the case study presented herein, wind environment input data for the WINDISPER model were obtained from the automatic weather station (AWS) installed on the surface of the harvested patch (SC). However, the data would only represent superficial characteristics and not the overall characteristics of the wind in the natural regeneration stand. Additional observational data from a nearby meteorological station would be of no value in this case. Large-eddy simulation (LES) can delineate the complex wind environment in heterogeneous canopy structures [36]; however, the computational requirement for LES operation is too high to be efficiently deployed in silviculture. Therefore, the spatiotemporal complexity of wind environments in the context of natural regeneration should be investigated. Multiple approaches would secure representative data suitable for model operation, combining the sensor network for continuously detecting high-frequency temporal patterns of the wind environment [102], UAV monitoring for 3D patterns of wind environments with high resolution [103,104], and regional-scale time-series analysis using a mountainous meteorology observation system [105].
4.3. Termination of Seed Flight via Collisions with Canopy or Ground
The seed flight is completed when it collides with the ground. The wind and slope directions on mountainous terrain determine this moment. The flight time and distance can be shortened or extended several times by considering the slope effect. Therefore, an algorithm in which the slope affects the horizontal flight distance of seeds must be developed using correction factors (Box 1, Table 2). Furthermore, developing a real-time simulation from the initiation of seed flight from the seeding tree to its landing on the ground and producing a 3D trajectory of seed dispersal would be more progressive. Visualization of 3D seed trajectory using a real-time simulator would be effective in enhancing field foresters’ understanding of seed dispersal and communication with stakeholders.
Table 2.
Correction factors for applying a seed dispersal distance which is predicted on flat terrain into that on up or down slope.
Box 1. A simple correction of seed dispersal distance for sloped terrain
The extended or reduced distance of seed dispersal caused by slope can be calculated using the following function (Equation (6)):
where dc indicates the corrected distance of seed dispersal for sloped terrain, df indicates the distance originally predicted for seed dispersal on flat terrain, and x indicates the slope angle in degrees.
According to Equation (6), compared to the seeds on flat terrain, the seeds on a downslope of 20° would fly 57% further; on the other hand, the seeds on an upslope of 20° would travel 27% less. When on a down-slope of 40°, seed dispersal distance can be extended approximately six times that of the flat terrain. This exponential increase suggests that the estimation of seed dispersal is very sensitive to the variation of seed dispersal direction and slope direction/inclination. Therefore, care should be taken, especially when a seed dispersal model is applied to a region of more complicated topography (e.g., mountains). The issue can be overcome in the short term by applying correction factors (Table 2) using Equation (6) to the predicted seed dispersal distance on the flat terrain. For example, if the seed dispersal distance is predicted to be 20 m on flat terrain, the expected dispersal distance on a downslope of 25° is 37.4 m (20 m multiplied by 1.87).
In general, the displacement design of seeded trees (number, location, cluster type, etc.) is closely associated with the success of natural regeneration. What is often overlooked in this work is that seed trees are not only the source of seeds but also an obstacle to seed dispersal by the wind. For example, there is a possibility that a seed released from the seeding trees may collide with a leaf or branch of a tree in a shelterbelt or remnant patch during flight, stopping it and sending it to the ground. Therefore, the seed dispersal model for natural regeneration requires an algorithm to quantify the effect of surrounding trees blocking seed dispersal [106].
However, the effects of blocked seed dispersal between trees have been neglected in previous studies. First, most studies have focused on the interpretation and simulation of the seed dispersal phenomenon; thus, the link between the model and forest management is not considered a high priority. Second, the empirical data to reflect this phenomenon are insufficient [107]. Recently, with the development of mechanical models and models applying large-eddy simulation (LES), models that can reflect airflow changes according to the canopy’s density and structure have been developed. However, falling owing to collisions with stems has not yet been well reflected in the model [36,39,108]. Therefore, in further modeling of the seed dispersal process, it is necessary to predict the probability of colliding with the physical structures of the stand through wind tunnel experiments [109,110], field observations, and simulations.
4.4. Secondary Dispersal, LDD, and Consumption by Herbivores
Seed dispersal is controlled by primary wind-dispersal at the local scale and by additive mechanisms such as secondary dispersal [111,112] and LDD [70,78,113]. Even after a seed lands on the ground and completes its flight, it can be exposed to various forces such as gravity, wind, water flow, and even animal predation, which transports it to unexpected places. For example, long-distance secondary dispersal downhill and overland by floating in rivers was reported for the tree-of-heaven (Alianthus altissima), a well-known wind-dispersing angiosperm species [15]. Secondary dispersal of wind-dispersed seeds by animals can occur; however, this is less likely to occur than animal-dispersed seeds [114,115]. Several studies using mechanistic modeling [112], field experiments [116,117], and wind tunnel experiments [16,118] have been conducted to elucidate the processes and patterns of secondary dispersal by wind. For example, Zhu et al. [16] reported a trade-off between primary and secondary dispersal, particularly of a smoother surface. A seed with a lower primary dispersal ability and faster terminal velocity has greater secondary dispersal ability because the greater vertical/horizontal ratio allows more opportunity for exposure to ground wind flow than the frictional force of the ground [16].
Based on the integration of currently available but insufficient knowledge, secondary dispersal is expected to significantly affect the spatial distribution of seeds and seedlings in the context of natural regeneration. Secondary dispersal by animals may not be sufficient for silvicultural species that generally produce wind-dispersed and small-sized seeds. However, the scarped ground surfaces of naturally regenerating patches have low roughness. They are exposed to wind and water flows, resulting in feasible conditions for secondary dispersal by wind or water. Experimental studies and field observations of the secondary dispersal of major silvicultural species in naturally regenerating patches are required.
Although LDD is an important mechanism for plant migration and colonization, it may be less meaningful in the context of natural regeneration. While a minority of seeds make the migration of species, metapopulations, or genes possible at the landscape, regional, and even intercontinental scales [70,78,113], most seeds correspond to the peak of the distribution curve and only move within tens of meters. These determine the fate of natural regeneration. In other words, interest in natural regeneration focuses on the local dispersal of seeds by standard mechanisms. LDD prediction may not be an urgent challenge; nevertheless, it is not necessary to exclude LDD prediction from seed dispersal models for natural regeneration.
Lastly, the consumption of seeds by herbivores during the pre-and post-dispersal stages can substantially slow natural regeneration [119,120,121]. Pre- and post-dispersal seeds are affected by tree species, seed production, the number and type of herbivores and predators, site characteristics, and climate conditions [119,120,121,122,123,124]. However, the impact of seed consumption by herbivores has been neglected in most cases because of its strong dependency on site-specific environments and limited empirical data.
5. Conclusions
In this study, we reviewed the literature on representative wind-dispersal models for seeds. A case study was also presented, which compared the observed spatial patterns of seed dispersal with the results of the WINDISPER model. Finally, based on these results, prospects for developing a seed dispersal model to support natural regeneration on mountainous terrains were suggested. Seed dispersal modeling has rarely been applied to natural regeneration practices despite advances in seed dispersal modeling along with improved analytical methodologies, observation equipment, simulation techniques, and computer systems. We believe that developing a seed dispersal model designed to support the planning and evaluation of natural regeneration programs will motivate the scientific community in this field to contribute to reducing uncertainties in the neglected aspects of seed dispersal. In the long run, efforts to improve seed dispersal models will help reduce the risk of natural regeneration and make it a reliable forest management practice.
For example, in South Korea, a nationwide afforestation program half a century ago led to the rehabilitation of formerly devastated mountainous lands covering approximately two-thirds of the national land cover [125,126,127,128]. Most afforestation and reforestation practices (over 200 km2 year−1) rely heavily on artificial regeneration, costing 10 billion KRW every year [129]. In 2021, the South Korean Forest Service announced a 3 billion tree-planting plan concluding in 2050 to support national carbon neutrality under the Paris Agreement, with planted trees being supplied by nurseries. Natural regeneration is not elected as a major practice in this plan but is allowed to be tested for experimental purposes [130,131,132]. Some of the early attempts at natural regeneration were unsuccessful because of the failure to control natural processes, such as seed production, dispersal, and germination [131,133]. Despite the need to expand the use of natural regeneration in terms of the economic compatibility of South Korean forestry [129], the uncertainty about the success of natural regeneration made decision-makers turn their back on it, favoring artificial regeneration instead [134]. However, advances in seed dispersal modeling will contribute to minimizing the uncertainty and risks in natural regeneration and support the practice thereof.
In addition, the development of wind-dispersal models will contribute greatly to advancing interdisciplinary research in climatology, biophysics, and forestry, such as (1) upgrading knowledge of the forest wind environment and wind-related forest disturbances, (2) estimating seed dispersal patterns and future species distribution by forest management and climate change, and (3) satisfying the societal needs for ecologically friendly forest management and net-zero carbon objectives.
Author Contributions
Conceptualization, T.K.Y., M.K. and H.S.K.; methodology, T.K.Y., H.S.K. and S.C.; software, K.Y., H.-S.K. and T.K.Y.; validation, T.K.Y.; formal analysis, S.L. (Seonghun Lee) and S.L. (Songhee Lee); investigation, J.C.; resources, J.C.; data curation, T.K.Y.; writing—original draft preparation, T.K.Y. and M.K.; writing—review and editing, T.K.Y. and M.K.; visualization, T.K.Y.; supervision, T.K.Y.; project administration, S.C. and H.S.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by a research fund from the Forest Technology and Management Research Center of the National Institute of Forest Science, Republic of Korea (SC0400-2021-01-2021, SC0400-2021-01-2022).
Data Availability Statement
The data are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A. Case of Seed Dispersal Simulation by the WINDISPER
To test the applicability of the currently available seed dispersal model, seed dispersal in a naturally regenerated stand of Japanese larch in South Korea was simulated using the WINDISPER model [29]. Japanese larch, a major conifer species in South Korean forestry, was introduced from Japan to Korea in the early 1900s. During the last five years (2016–2020), Japanese larch was planted in 17% of the afforestation and reforestation stands (234 km2 year−1) in South Korea [135].
Appendix A.1. Study Site
The study site is located in central Korea (37°54′47″ N, 127°20′3″ E, elevation: 360 m). Four different harvesting methods (i.e., seed tree (ST), grouped seed tree (GST), reserved seed tree (RST), and strip clear-cut (SC)) was used in a Japanese larch plantation in September 2016 [136]. The plantation was originally established in the 1970s, and natural regeneration is practiced. The mean diameter at breast height and height of the pre-harvesting stand were 37.2 cm (31.6–41.1 cm) and 27.4 m (26.6–28.3 m), respectively [136]. After harvesting, stand density decreased from 165 to 20 trees ha−1 for ST, 20 trees ha−1 for GST, 40 trees ha−1 for RST, and 0 trees ha−1 for SC (Figure A1a,c). Two shelterbelts consisting of the remaining larch trees were placed above and below the SC (Figure A1a). Soil scarification and scraping were performed after harvest to create feasible conditions for seed germination. The organic horizon, including the forest floor and humus layer, harvest residues, and removable rocks, were removed using a forestry excavator (EC55C, Volvo) assembled with a wood grapple and bucket. Mesh fences (height: 1.6 m) were installed along the boundaries of regeneration patches to prevent the invasion of wild animals.
Appendix A.2. Seed Dispersal Monitoring
A total of 102 seed traps were installed to monitor seed dispersal (Figure A1b). The seed traps were evenly distributed in each patch using a square lattice design (e.g., 5 × 3 for ST and RST, 3 × 3 for GST, and 3 × 5 for SC). In addition, one and three sets of 12 seed traps each were installed in ST and GST, respectively, to observe the distance-dependence pattern of seed dispersal by distance from a selected seed tree. In each set, 12 seed traps (4 sides × 3 distances) were installed 0, 10, and 20 m from the four sides of a selected seed tree (“radial design”). The seed traps under the square lattice and radial designs were integrated into the geospatial analysis. Each seed trap covered 0.25 m2 and was installed 1 m above the ground.
Seeds in the traps were retrieved monthly from October 2016–April 2017 and the numbers were recorded. Monthly monitoring might be limited to observing fine-scale temporal patterns of seed dispersal; nevertheless, this was the best monitoring interval for our limited workforce. The proportion of monthly seed inflow to total seed inflow during the seed-fall period was 5.3% in September, 15.3% in October, 16.7% in February, and 10.1% in March and April [31]. The germination rate from seed quality testing was over 40% for seeds trapped before November 2016 and decreased from 27.8% in December 2016 to 12.8% in February 2017.
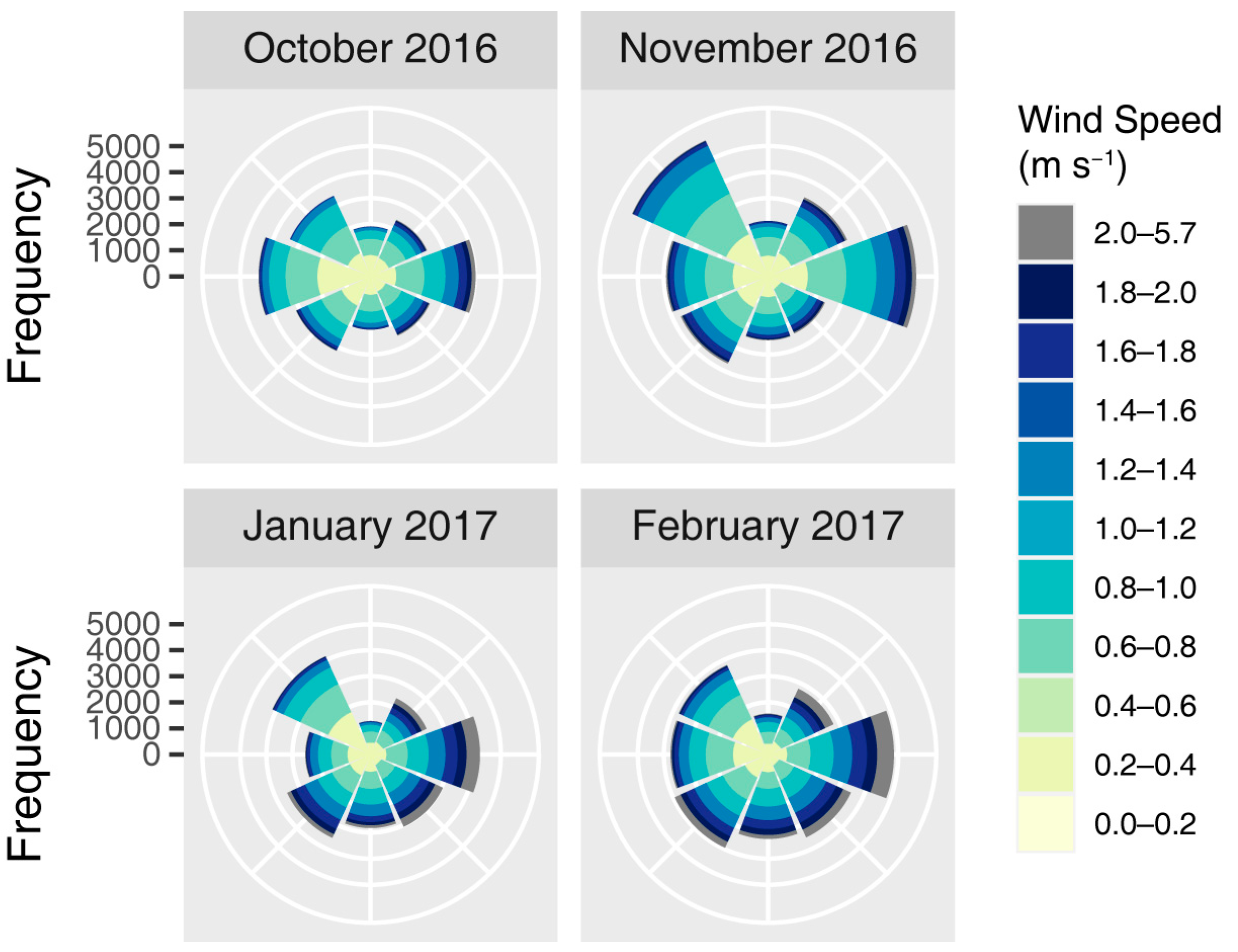
The coordinates of all seed trees and traps were recorded in the field using a global navigation satellite system (GNSS) (Trimble R2, Trimble, CA, USA). The climate of the stand, including wind direction and speed, was recorded every 5 min by an AWS located at the center of the SC. The wind direction patterns for the seed dispersal season of 2016-2017 were in line with the typical wind environment in this region, in which northwest wind was predominant (Figure A2).

Figure A1.
Placement of natural regeneration patches and seed trees (a), placement of seed traps (b), and photographs of the study site taken in May 2020 (c).
Figure A1.
Placement of natural regeneration patches and seed trees (a), placement of seed traps (b), and photographs of the study site taken in May 2020 (c).

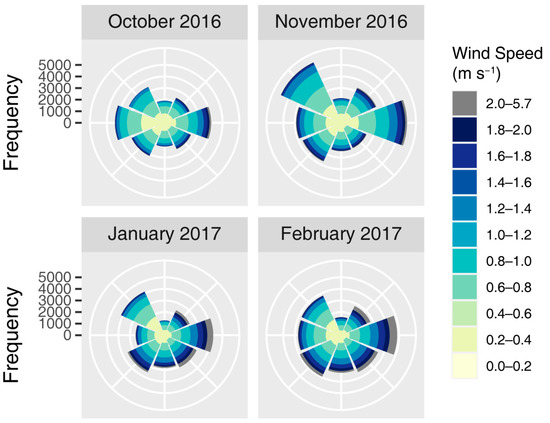
Figure A2.
Monthly wind rose plots in the seed dispersal season of 2016–2017. Data in December 2016 was unavailable because of the AWS failure.
Figure A2.
Monthly wind rose plots in the seed dispersal season of 2016–2017. Data in December 2016 was unavailable because of the AWS failure.
Appendix A.3. Modeling Procedures
The WINDISPER model is a mechanistic model based on a ballistic algorithm for simulating the trajectory of seed flight caused by wind [39]. The input variables for model operation include the arrangement and height of seed trees, the terminal velocity of seeds, density and vertical distribution of seeds in the crown, wind direction and speed. Finally, the model output is a number representing the number of dispersed seeds in each grid with a predefined size (e.g., 1 m × 1 m). The WINDISPER model, scripted in C language, is open source software [29].
Key input parameters were prepared from various field surveys and experiments for model operation. The terminal velocity of the seed was 0.98 m s−1 (Lee et al. unpublished), determined using video camera recordings [137]. The arrangement of the seed trees followed the coordinates of the seed trees obtained from the field survey (Figure A1a). To determine the number and vertical distribution of seeds in a seed tree, three harvested trees were analyzed in September 2016. The number of cones in the upper, middle, and lower crowns, total number of cones, and number of seeds per cone were measured. The mean bole height of the seed trees was 10.1 m. The mean and standard deviation of the portion height were set to 0.70 ± 0.087. Seed production in 2016 was estimated to be 690,339 seeds tree−1. Based on the ratio of the number of seeds collected from seed traps to the estimate of total seed production, it was assumed that 29% of the seeds in cones were abscised and dispersed [133]. Assuming that the number of abscised seeds was 210,000 seeds tree−1, the crown diameter was 6 m, and the seed dispersal period was 180 d, the seed abscission rate was estimated to be 39 seeds m−2 d−1.
The seeds were not uniformly abscised at the different wind speeds. A wind speed threshold for seed abscission in Japanese larch can be assumed; however, an accurate value of this figure remains unknown. Because the relationship between wind speed and seed abscission is unavailable, we simulated seed dispersal based on four scenarios that assumed different wind speed thresholds for seed abscission (0, 0.5, 1, and 2 m s−1). In each scenario, wind speeds above the threshold were extracted from the entire dataset for the seed dispersal period (October 2016–February 2017). Observations above the wind speed threshold at 0, 0.5, 1, and 2 m s−1 represented 64.6%, 45.1%, 15.4%, and 2.1% of the entire dataset, respectively (Table A1). The wind speeds in the extracted dataset were assigned to eight wind directions (N, NE, E, SE, S, SW, W, and NW) and were natural log-transformed to satisfy the required structure for the wind input data of the WINDISPER model.
The wind directions were evenly distributed when the wind speed above 0 m s−1 was analyzed. The proportions of the wind direction ranged from 6.6% (N) to 18.7% (E) (Table A1); however, the proportion of winds from the east (90°) notably increased to 40.6% at 2 m s−1 of the wind speed threshold (Table A1). The dominance of breezes and gales faster than 2 m s−1 from east to west was probably due to the topography of the studied stand, in which the valley developed towards the west. The shape of the SC, in which the AWS was placed, may also have influenced this, as it elongated from east to west.
Table A1.
Wind flow characteristics by applying different wind speed thresholds (0, 0.5, 1, and 2 m s−1) to a naturally regenerating Japanese larch site during the seed dispersion period (from October 2016–February 2017).
Table A1.
Wind flow characteristics by applying different wind speed thresholds (0, 0.5, 1, and 2 m s−1) to a naturally regenerating Japanese larch site during the seed dispersion period (from October 2016–February 2017).
| Wind Direction (°) | 0 m s−1 | 0.5 m s−1 | 1 m s−1 | 2 m s−1 | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | % | Mean | % | Mean | % | Mean | % | |
| 0 | 0.60 | 6.6 | 0.86 | 5.8 | 1.48 | 3.0 | 2.51 | 0.7 |
| 45 | 0.78 | 10.6 | 1.07 | 11.1 | 1.71 | 12.6 | 2.69 | 16.7 |
| 90 | 0.86 | 18.7 | 1.13 | 20.8 | 1.74 | 27.1 | 2.68 | 40.6 |
| 135 | 0.88 | 10.5 | 1.12 | 12.0 | 1.69 | 15.6 | 2.64 | 18.9 |
| 180 | 0.72 | 9.6 | 1.00 | 9.7 | 1.57 | 9.5 | 2.55 | 6.5 |
| 225 | 0.72 | 13.2 | 1.04 | 12.8 | 1.61 | 14.5 | 2.60 | 10.2 |
| 270 | 0.61 | 13.8 | 0.93 | 11.8 | 1.53 | 8.5 | 2.53 | 4.6 |
| 315 | 0.64 | 16.9 | 0.89 | 15.9 | 1.45 | 9.1 | 2.44 | 1.7 |
| Total * | 0.72 | 64.6 | 1.02 | 45.1 | 1.64 | 15.4 | 2.65 | 2.1 |
* Percentages in the total column indicate the proportion of frequency over the wind speed threshold for all observations.
The other input parameters unavailable at our study site were assigned default values of the WINDISPER model. The source codes of the WINDISPER model were compiled and run in Visual Studio 2019 (Microsoft) to simulate seed dispersal from early autumn (September) to late winter (February). We modified the random number generator to allow the generation of stochastic distributions of wind speed, the terminal velocity of seeds, vertical distribution of seeds, etc., from their mean and standard deviation defined by the user, as this functionality was not provided in the source codes of the WINDISPER.
The output data of the wind-dispersal simulation were analyzed using R [138]. The relationships between the observed (i.e., seed trap-based) and simulated seed dispersal were tested using simple linear regressions with combinations of the four wind speed thresholds and natural regeneration methods (ST, GST, RST, SC, and global). To visualize the spatial patterns of seed dispersal, the ggplot2 package in R [139] and QGIS [140] was used.
Appendix A.4. Results of Seed Dispersal Simulation Using the WINDISPER
Overall, in the simulations adopting a lower wind speed threshold (0 and 0.5 m s−1), seed dispersal mainly occurred where seed trees were densely distributed (e.g., the west side of SC and the downside of the GST) (see Figure 2c,d). In contrast, simulations adopting a larger wind speed threshold (1 and 2 m s−1) estimated widely scattered seed dispersal around the natural regeneration patches (see Figure 2e,f). The estimated spatial patterns of seed dispersal may differ depending on the wind speed threshold adopted. The strong sensitivity of predicted seed dispersal to wind speed is not surprising, given that seed dispersal is mechanistically dependent on wind flow. The stronger the wind and taller the tree, the longer and further the seeds will fly.
In comparison with the measured seed dispersal (i.e., the number of seeds trapped in seed traps), none of the simulations satisfactorily reproduced the spatial patterns of measured seed dispersal (Table A2). A statistically significant linear relationship between predicted seed dispersal using 0 and 2 m s−1 and measured seed dispersal was observed in the dataset of all natural regeneration patches (i.e., global); however, the coefficient of determination was insignificant (R2 = 0.035 and 0.038, respectively). In addition, linear regression was tested separately in each natural regeneration patch to account for the differences in stand structure and wind flow between the natural regeneration methods (Table A2). Nevertheless, small improvements in the model reliability were observed. For example, the observed seed dispersals were best fitted by seed dispersal predicted using a wind speed threshold of 1 m s−1 (R2 = 0.321) for the SC patch and 2 m s−1 (R2 = 0.357) for the RST. The observed seed dispersal patterns of ST and GST were not predicted by simulations using any wind speed threshold.
Table A2.
Simple linear regressions between spatial patterns of predicted and observed seed dispersal by applying different wind speeds threshold (0, 0.5, 1, 2 m s−1) and by natural regeneration patches. A dash indicates a statistically insignificant R2 (p > 0.10).
Table A2.
Simple linear regressions between spatial patterns of predicted and observed seed dispersal by applying different wind speeds threshold (0, 0.5, 1, 2 m s−1) and by natural regeneration patches. A dash indicates a statistically insignificant R2 (p > 0.10).
| 0 m s−1 | 0.5 m s−1 | 1 m s−1 | 2 m s−1 | |||||
|---|---|---|---|---|---|---|---|---|
| R2 | p-Value | R2 | p-Value | R2 | p-Value | R2 | p-Value | |
| Seed tree method | – | 0.793 | – | 0.515 | – | 0.615 | – | 0.981 |
| Grouped seed tree | 0.091 | 0.044 | 0.067 | 0.086 | 0.077 | 0.064 | – | 0.109 |
| Reserved seed tree | – | 0.246 | – | 0.436 | – | 0.567 | 0.357 | 0.019 |
| Strip cutting | – | 0.478 | – | 0.147 | 0.321 | 0.028 | – | 0.525 |
| Global | 0.035 | 0.059 | – | 0.102 | – | 0.636 | 0.038 | 0.050 |
References
- Boydak, M. Silvicultural characteristics and natural regeneration of Pinus brutia Ten.—A review. Plant Ecol. 2004, 171, 153–163. [Google Scholar] [CrossRef]
- Crouzeilles, R.; Beyer, H.L.; Monteiro, L.M.; Feltran-Barbieri, R.; Pessôa, A.C.; Barros, F.S.; Lindenmayer, D.B.; Lino, E.D.; Grelle, C.E.; Chazdon, R.L. Achieving cost-effective landscape-scale forest restoration through targeted natural regeneration. Conserv. Lett. 2020, 13, e12709. [Google Scholar] [CrossRef]
- Crouzeilles, R.; Ferreira, M.S.; Chazdon, R.L.; Lindenmayer, D.B.; Sansevero, J.B.; Monteiro, L.; Iribarrem, A.; Latawiec, A.E.; Strassburg, B.B. Ecological restoration success is higher for natural regeneration than for active restoration in tropical forests. Sci. Adv. 2017, 3, e1701345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gärtner, S.M.; Lieffers, V.J.; Macdonald, S.E. Ecology and management of natural regeneration of white spruce in the boreal forest. Environ. Rev. 2011, 19, 461–478. [Google Scholar] [CrossRef]
- Kohler, M.; Pyttel, P.; Kuehne, C.; Modrow, T.; Bauhus, J. On the knowns and unknowns of natural regeneration of silviculturally managed sessile oak (Quercus petraea (Matt.) Liebl.) forests—A literature review. Ann. For. Sci. 2020, 77, 101. [Google Scholar] [CrossRef]
- Goerlich, D.L.; Nyland, R.D. Natural regeneration of eastern hemlock: A review. In Symposium on Sustainable Management of Hemlock Ecosystems in Eastern North America. Gen. Tech. Rep. NE-267; McManus Katherine, A., Shields Kathleen, S., Souto Dennis, R., Eds.; US Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Newtown Square, PA, USA, 2000; pp. 14–22. [Google Scholar]
- Ashton, M.S.; Kelty, M.J. Chater 5. Ecology of regerneration. In The Practice of Silviculture: Applied Forest Ecology; John Wiley & Sons Ltd.: West Sussex, UK, 2018; pp. 80–116. [Google Scholar]
- Dey, D.C.; Knapp, B.O.; Battaglia, M.A.; Deal, R.L.; Hart, J.L.; O’Hara, K.L.; Schweitzer, C.J.; Schuler, T.M. Barriers to natural regeneration in temperate forests across the USA. New For. 2019, 50, 11–40. [Google Scholar] [CrossRef]
- Kaliszewski, A. Cost analysis of artificial and natural oak regeneration in selected forest districts. For. Res. Pap. 2018, 78, 315–321. [Google Scholar] [CrossRef]
- Minami, S.; Azuma, A. Various flying modes of wind-dispersal seeds. J. Theor. Biol. 2003, 225, 1–14. [Google Scholar] [CrossRef]
- Tan, K.; DONG, S.-P.; Lu, T.; ZHANG, Y.-J.; XU, S.-T.; REN, M.-X. Diversity and evolution of samara in angiosperm. Chin. J. Plant Ecol. 2018, 42, 806. [Google Scholar] [CrossRef] [Green Version]
- Landhäusser, S.M.; Pinno, B.D.; Mock, K.E. Tamm Review: Seedling-based ecology, management, and restoration in aspen (Populus tremuloides). For. Ecol. Manag. 2019, 432, 231–245. [Google Scholar] [CrossRef]
- Zakaria, M.Y.; Dos Santos, C.R.; Dayhoum, A.; Marques, F.; Hajj, M.R. Modeling and prediction of aerodynamic characteristics of free fall rotating wing based on experiments. In Proceedings of the International Conference on Aerospace Sciences and Aviation Technology, Cairo, Egypt, 9–11 April 2019; pp. 1–15. [Google Scholar]
- Lentink, D.; Dickson, W.B.; Van Leeuwen, J.L.; Dickinson, M.H. Leading-edge vortices elevate lift of autorotating plant seeds. Science 2009, 324, 1438–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaproth, M.A.; McGraw, J.B. Seed viability and dispersal of the wind-dispersed invasive Ailanthus altissima in aqueous environments. For. Sci. 2008, 54, 490–496. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, M.; Xin, Z.; Liu, Z.; Schurr, F.M. A trade-off between primary and secondary seed dispersal by wind. Plant Ecol. 2019, 220, 541–552. [Google Scholar] [CrossRef]
- Carlo, T.A.; García, D.; Martínez, D.; Gleditsch, J.M.; Morales, J.M. Where do seeds go when they go far? Distance and directionality of avian seed dispersal in heterogeneous landscapes. Ecology 2013, 94, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Montoya, D.; Zavala, M.A.; Rodríguez, M.A.; Purves, D.W. Animal versus wind dispersal and the robustness of tree species to deforestation. Science 2008, 320, 1502–1504. [Google Scholar] [CrossRef] [Green Version]
- Thomson, F.J.; Moles, A.T.; Auld, T.D.; Kingsford, R.T. Seed dispersal distance is more strongly correlated with plant height than with seed mass. J. Ecol. 2011, 99, 1299–1307. [Google Scholar] [CrossRef]
- Harmer, R. Natural regeneration of broadleaved trees in Britain: II Seed production and predation. Forestry 1994, 67, 275–286. [Google Scholar] [CrossRef]
- Barna, M. The effects of cutting regimes on natural regeneration in submountain beech forests: Species diversity and abundance. J. For. Sci. 2008, 54, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Stoehr, M.U. Seed production of western larch in seed-tree systems in the southern interior of British Columbia. For. Ecol. Manag. 2000, 130, 7–15. [Google Scholar] [CrossRef]
- Kuparinen, A. Mechanistic models for wind dispersal. Trends Plant Sci. 2006, 11, 296–301. [Google Scholar] [CrossRef]
- Gregory, P. Interpreting plant disease dispersal gradients. Annu. Rev. Phytopathol. 1968, 6, 189–212. [Google Scholar] [CrossRef]
- Clark, J.S.; Silman, M.; Kern, R.; Macklin, E.; HilleRisLambers, J. Seed dispersal near and far: Patterns across temperate and tropical forests. Ecology 1999, 80, 1475–1494. [Google Scholar] [CrossRef]
- Frampton, V.L. The spread of virus diseases of the yellows type under field conditions. Phytopathology 1942, 32, 799–808. [Google Scholar]
- Katul, G.; Porporato, A.; Nathan, R.; Siqueira, M.; Soons, M.; Poggi, D.; Horn, H.; Levin, S. Mechanistic analytical models for long-distance seed dispersal by wind. Am. Nat. 2005, 166, 368–381. [Google Scholar] [CrossRef] [PubMed]
- Portnoy, S.; Willson, M.F. Seed dispersal curves: Behavior of the tail of the distribution. Evol. Ecol. 1993, 7, 25–44. [Google Scholar] [CrossRef]
- Nathan, R.; Safriel, U.N.; Noy-Meir, I. Field validation and sensitivity analysis of a mechanistic model for tree seed dispersal by wind. Ecology 2001, 82, 374–388. [Google Scholar] [CrossRef]
- Kruse, S.; Gerdes, A.; Kath, N.J.; Herzschuh, U. Implementing spatially explicit wind-driven seed and pollen dispersal in the individual-based larch simulation model: LAVESI-WIND 1.0. Geosci. Model Dev. 2018, 11, 4451–4467. [Google Scholar] [CrossRef] [Green Version]
- Okubo, A.; Levin, S.A. A theoretical framework for data analysis of wind dispersal of seeds and pollen. Ecology 1989, 70, 329–338. [Google Scholar] [CrossRef]
- Tackenberg, O. Modeling long-distance dispersal of plant diaspores by wind. Ecol. Monogr. 2003, 73, 173–189. [Google Scholar] [CrossRef]
- Horn, S.; Raabe, A.; Will, H.; Tackenberg, O. TurbSeed—a model for wind dispersal of seeds in turbulent currents based on publicly available climate data. Ecol. Model. 2012, 237, 1–10. [Google Scholar] [CrossRef]
- Loos, C.; Seppelt, R.; Meier-Bethke, S.; Schiemann, J.; Richter, O. Spatially explicit modelling of transgenic maize pollen dispersal and cross-pollination. J. Theor. Biol. 2003, 225, 241–255. [Google Scholar] [CrossRef]
- Aylor, D.E.; Flesch, T.K. Estimating spore release rates using a Lagrangian stochastic simulation model. J. Appl. Meteorol. Climatol. 2001, 40, 1196–1208. [Google Scholar] [CrossRef]
- Bohrer, G.; Katul, G.G.; Walko, R.L.; Avissar, R. Exploring the effects of microscale structural heterogeneity of forest canopies using large-eddy simulations. Bound.-Layer Meteorol. 2009, 132, 351–382. [Google Scholar] [CrossRef]
- Di-Giovanni, F.; Beckett, P. On the mathematical modeling of pollen dispersal and deposition. J. Appl. Meteorol. Climatol. 1990, 29, 1352–1357. [Google Scholar] [CrossRef] [Green Version]
- Maurer, K.D.; Bohrer, G.; Medvigy, D.; Wright, S.J. The timing of abscission affects dispersal distance in a wind-dispersed tropical tree. Funct. Ecol. 2013, 27, 208–218. [Google Scholar] [CrossRef]
- Nathan, R.; Katul, G.G.; Bohrer, G.; Kuparinen, A.; Soons, M.B.; Thompson, S.E.; Trakhtenbrot, A.; Horn, H.S. Mechanistic models of seed dispersal by wind. Theor. Ecol. 2011, 4, 113–132. [Google Scholar] [CrossRef]
- Thompson, S.E.; Assouline, S.; Chen, L.; Trahktenbrot, A.; Svoray, T.; Katul, G.G. Secondary dispersal driven by overland flow in drylands: Review and mechanistic model development. Mov. Ecol. 2014, 2, 7. [Google Scholar] [CrossRef] [Green Version]
- Treep, J.; de Jager, M.; Kuiper, L.S.; Duman, T.; Katul, G.G.; Soons, M.B. Costs and benefits of non-random seed release for long-distance dispersal in wind-dispersed plant species. Oikos 2018, 127, 1330–1343. [Google Scholar] [CrossRef] [Green Version]
- Treep, J.; de Jager, M.; Bartumeus, F.; Soons, M.B. Seed dispersal as a search strategy: Dynamic and fragmented landscapes select for multi-scale movement strategies in plants. Mov. Ecol. 2021, 9, 4. [Google Scholar] [CrossRef]
- Williams, C.G.; LaDeau, S.L.; Oren, R.; Katul, G.G. Modeling seed dispersal distances: Implications for transgenic Pinus taeda. Ecol. Appl. 2006, 16, 117–124. [Google Scholar] [CrossRef]
- Wright, S.J.; Trakhtenbrot, A.; Bohrer, G.; Detto, M.; Katul, G.G.; Horvitz, N.; Muller-Landau, H.C.; Jones, F.A.; Nathan, R. Understanding strategies for seed dispersal by wind under contrasting atmospheric conditions. PNAS 2008, 105, 19084–19089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiyosawa, S.; Shiyomi, M. A theoretical evaluation of the effect of mixing resistant variety with susceptible variety for controlling plant diseases. JJP 1972, 38, 41–51. [Google Scholar] [CrossRef]
- McCartney, H.; Bainbridge, A. Deposition gradients near to a point source in a barley crop. J. Phytopathol. 1984, 109, 219–236. [Google Scholar] [CrossRef]
- Ribbens, E.; Silander, J.A., Jr.; Pacala, S.W. Seedling recruitment in forests: Calibrating models to predict patterns of tree seedling dispersion. Ecology 1994, 75, 1794–1806. [Google Scholar] [CrossRef]
- Stoyon, D.; Wagner, S. Estimating the fruit dispersion of anemochorous trees. Ecol. Model. 2001, 145, 35–47. [Google Scholar] [CrossRef]
- Skarpaas, O.; Stabbetorp, O.E.; Rønning, I.; Svennungsen, T.O. How far can a hawk’s beard fly? Measuring and modelling the dispersal of Crepis praemorsa. J. Ecol. 2004, 92, 747–757. [Google Scholar] [CrossRef]
- Morales, J.M.; López, T.M. Mechanistic models of seed dispersal by animals. Oikos 2022. [Google Scholar] [CrossRef]
- Wichmann, M.C.; Alexander, M.J.; Soons, M.B.; Galsworthy, S.; Dunne, L.; Gould, R.; Fairfax, M.; Niggemann, M.; Hails, R.S.; Bullock, J.M. Human-mediated dispersal of seeds over long distances. Proc. R. Soc. B: Biol. Sci. 2009, 276, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Greene, D.F.; Canham, C.D.; Coates, K.D.; Lepage, P.T. An evaluation of alternative dispersal functions for trees. J. Ecol. 2004, 92, 758–766. [Google Scholar] [CrossRef]
- LePage, P.T.; Canham, C.D.; Coates, K.D.; Bartemucci, P. Seed abundance versus substrate limitation of seedling recruitment in northern temperate forests of British Columbia. Can. J. For. Res. 2000, 30, 415–427. [Google Scholar] [CrossRef]
- Soons, M.B.; Bullock, J.M. Non-random seed abscission, long-distance wind dispersal and plant migration rates. J. Ecol. 2008, 96, 581–590. [Google Scholar] [CrossRef]
- Greene, D.; Johnson, E. A model of wind dispersal of winged or plumed seeds. Ecology 1989, 70, 339–347. [Google Scholar] [CrossRef]
- Levin, S.A.; Muller-Landau, H.C.; Nathan, R.; Chave, J. The ecology and evolution of seed dispersal: A theoretical perspective. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 575–604. [Google Scholar] [CrossRef]
- Nathan, R.; Getz, W.M.; Revilla, E.; Holyoak, M.; Kadmon, R.; Saltz, D.; Smouse, P.E. A movement ecology paradigm for unifying organismal movement research. Proc. Natl. Acad. Sci. USA 2008, 105, 19052–19059. [Google Scholar] [CrossRef] [Green Version]
- Nuttle, T.; Haefner, J.W. Seed dispersal in heterogeneous environments: Bridging the gap between mechanistic dispersal and forest dynamics models. Am. Nat. 2005, 165, 336–349. [Google Scholar] [CrossRef] [PubMed]
- Axer, M.; Schlicht, R.; Wagner, S. Modelling potential density of natural regeneration of European oak species (Quercus robur L., Quercus petraea (Matt.) Liebl.) depending on the distance to the potential seed source: Methodological approach for modelling dispersal from inventory data at forest enterprise level. For. Ecol. Manag. 2021, 482, 118802. [Google Scholar] [CrossRef]
- de Andrés, E.G.; Camarero, J.J.; Martínez, I.; Coll, L. Uncoupled spatiotemporal patterns of seed dispersal and regeneration in Pyrenean silver fir populations. For. Ecol. Manag. 2014, 319, 18–28. [Google Scholar] [CrossRef]
- Holmström, E.; Karlsson, M.; Nilsson, U. Modeling birch seed supply and seedling establishment during forest regeneration. Ecol. Model. 2017, 352, 31–39. [Google Scholar] [CrossRef]
- Manso, R.; Pardos, M.; Keyes, C.R.; Calama, R. Modelling the spatio-temporal pattern of primary dispersal in stone pine (Pinus pinea L.) stands in the Northern Plateau (Spain). Ecol. Model. 2012, 226, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Katul, G.G.; Albertson, J.D. Modeling CO2 sources, sinks, and fluxes within a forest canopy. J. Geophys. Res. Atmos. 1999, 104, 6081–6091. [Google Scholar] [CrossRef] [Green Version]
- Stockie, J.M. The mathematics of atmospheric dispersion modeling. Siam Rev. 2011, 53, 349–372. [Google Scholar] [CrossRef]
- Di-Giovanni, F.; Beckett, P.; Flenley, J. Modelling of dispersion and deposition of tree pollen within a forest canopy. Grana 1989, 28, 129–139. [Google Scholar] [CrossRef]
- McCartney, H.; Lacey, M.E. Wind dispersal of pollen from crops of oilseed rape (Brassica napus L.). J. Aerosol. Sci. 1991, 22, 467–477. [Google Scholar] [CrossRef]
- Andersen, M. Mechanistic models for the seed shadows of wind-dispersed plants. Am. Nat. 1991, 137, 476–497. [Google Scholar] [CrossRef]
- Soons, M.B.; Heil, G.W.; Nathan, R.; Katul, G.G. Determinants of long-distance seed dispersal by wind in grasslands. Ecology 2004, 85, 3056–3068. [Google Scholar] [CrossRef] [Green Version]
- Jarosz, N.; Loubet, B.; Huber, L. Modelling airborne concentration and deposition rate of maize pollen. Atmos. Environ. 2004, 38, 5555–5566. [Google Scholar] [CrossRef] [Green Version]
- Nathan, R.; Katul, G.G.; Horn, H.S.; Thomas, S.M.; Oren, R.; Avissar, R.; Pacala, S.W.; Levin, S.A. Mechanisms of long-distance dispersal of seeds by wind. Nature 2002, 418, 409–413. [Google Scholar] [CrossRef]
- Boehm, M.T.; Aylor, D.E. Lagrangian stochastic modeling of heavy particle transport in the convective boundary layer. Atmos. Environ. 2005, 39, 4841–4850. [Google Scholar] [CrossRef]
- Poggi, D.; Katul, G.; Albertson, J. Scalar dispersion within a model canopy: Measurements and three-dimensional Lagrangian models. Adv. Water Resour. 2006, 29, 326–335. [Google Scholar] [CrossRef]
- Nathan, R.; Katul, G.G. Foliage shedding in deciduous forests lifts up long-distance seed dispersal by wind. Proc. Natl. Acad. Sci. USA 2005, 102, 8251–8256. [Google Scholar] [CrossRef] [Green Version]
- Trakhtenbrot, A.; Katul, G.; Nathan, R. Mechanistic modeling of seed dispersal by wind over hilly terrain. Ecol. Model. 2014, 274, 29–40. [Google Scholar] [CrossRef]
- Bullock, J.M.; Clarke, R.T. Long distance seed dispersal by wind: Measuring and modelling the tail of the curve. Oecologia 2000, 124, 506–521. [Google Scholar] [CrossRef] [PubMed]
- Dauer, J.T.; Mortensen, D.A.; Vangessel, M.J. Temporal and spatial dynamics of long-distance Conyza canadensis seed dispersal. J. Appl. Ecol. 2007, 44, 105–114. [Google Scholar] [CrossRef]
- Levey, D.J.; Tewksbury, J.J.; Bolker, B.M. Modelling long-distance seed dispersal in heterogeneous landscapes. J. Ecol. 2008, 96, 599–608. [Google Scholar] [CrossRef] [Green Version]
- Cain, M.L.; Milligan, B.G.; Strand, A.E. Long-distance seed dispersal in plant populations. Am. J. Bot. 2000, 87, 1217–1227. [Google Scholar] [CrossRef] [Green Version]
- Kot, M.; Lewis, M.A.; van den Driessche, P. Dispersal data and the spread of invading organisms. Ecology 1996, 77, 2027–2042. [Google Scholar] [CrossRef]
- Ouborg, N.; Piquot, Y.; Van Groenendael, J. Population genetics, molecular markers and the study of dispersal in plants. J. Ecol. 1999, 87, 551–568. [Google Scholar] [CrossRef]
- Turchin, P. Quantitative Analysis of Movement: Measuring and Modeling Population Redistribution in Animals and Plants; Sinauer Associates: Sunderland, MA, USA, 1998. [Google Scholar]
- Webster, M.S.; Marra, P.P.; Haig, S.M.; Bensch, S.; Holmes, R.T. Links between worlds: Unraveling migratory connectivity. Trends Ecol. Evol. 2002, 17, 76–83. [Google Scholar] [CrossRef]
- Nathan, R.; Perry, G.; Cronin, J.T.; Strand, A.E.; Cain, M.L. Methods for estimating long-distance dispersal. Oikos 2003, 103, 261–273. [Google Scholar] [CrossRef] [Green Version]
- National Geographic Information Institute. The National Atlas of Korea II.; National Geographic Information Institute, Gyeonggi–do 16517, Korea. Available online: http://nationalatlas.ngii.go.kr/ (accessed on 20 August 2021).
- Katul, G.; Poggi, D. The effects of gentle topographic variation on dispersal kernels of inertial particles. Geophys. Res. Lett. 2012, 39. [Google Scholar] [CrossRef] [Green Version]
- Wyse, S.V.; Hulme, P.E.; Holland, E.P. Partitioning intraspecific variation in seed dispersal potential using a low-cost method for rapid estimation of samara terminal velocity. Methods Ecol. Evol. 2019, 10, 1298–1307. [Google Scholar] [CrossRef]
- Pazos, G.E.; Greene, D.F.; Katul, G.; Bertiller, M.B.; Soons, M.B. Seed dispersal by wind: Towards a conceptual framework of seed abscission and its contribution to long-distance dispersal. J. Ecol. 2013, 101, 889–904. [Google Scholar] [CrossRef]
- Schippers, P.; Jongejans, E. Release thresholds strongly determine the range of seed dispersal by wind. Ecol. Model. 2005, 185, 93–103. [Google Scholar] [CrossRef]
- Darling, E.; Samis, K.E.; Eckert, C.G. Increased seed dispersal potential towards geographic range limits in a Pacific coast dune plant. New Phytol. 2008, 178, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, B.; González, E. Pulses of seed release in riparian Salicaceae coincide with high atmospheric temperature. River Res. Appl. 2019, 35, 1590–1596. [Google Scholar] [CrossRef]
- Murray, B.R.; Brown, A.; Dickman, C.; Crowther, M. Geographical gradients in seed mass in relation to climate. J. Biogeogr. 2004, 31, 379–388. [Google Scholar] [CrossRef]
- Niklas, K.J. The aerodynamics of wind pollination. Bot. Rev. 1985, 51, 328–386. [Google Scholar] [CrossRef]
- Dawson, C.; Vincent, J.F.; Rocca, A.-M. How pine cones open. Nature 1997, 390, 668. [Google Scholar] [CrossRef]
- Song, K.; Chang, S.S.; Lee, S.J. How the pine seeds attach to/detach from the pine cone scale? Front. Life Sci. 2017, 10, 38–47. [Google Scholar] [CrossRef]
- Greene, D.F. The role of abscission in long-distance seed dispersal by the wind. Ecology 2005, 86, 3105–3110. [Google Scholar] [CrossRef]
- Endalew, A.M.; Hertog, M.; Delele, M.; Baetens, K.; Persoons, T.; Baelmans, M.; Ramon, H.; Nicolaï, B.; Verboven, P. CFD modelling and wind tunnel validation of airflow through plant canopies using 3D canopy architecture. Int. J. Heat Fluid Flow 2009, 30, 356–368. [Google Scholar] [CrossRef]
- Lee, J.P.; Lee, S.J. PIV analysis on the shelter effect of a bank of real fir trees. J. Wind Eng. Ind. Aerodyn. 2012, 110, 40–49. [Google Scholar] [CrossRef]
- Clark, T.L.; Mitchell, S.J. Three-dimensional simulations of air flow and momentum transfer in partially harvested forests. Bound. Layer Meteorol. 2007, 125, 505–524. [Google Scholar] [CrossRef]
- Novak, M.D.; Warland, J.S.; Orchansky, A.L.; Ketler, R.; Green, S. Wind tunnel and field measurements of turbulent flow in forests. Part I: Uniformly thinned stands. Bound. Layer Meteorol. 2000, 95, 457–495. [Google Scholar] [CrossRef]
- Daikoku, K.; Hattori, S.; Deguchi, A.; Fujita, Y.; Park, H.; Matsumoto, K. Impact of wind direction on diurnal and seasonal changes in wind profiles. J. For. Res. 2007, 12, 452–466. [Google Scholar] [CrossRef]
- Ha, T.; Lee, I.-b.; Kwon, K.-s.; Lee, S.-J. Development of a micro-scale CFD model to predict wind environment on mountainous terrain. Comput. Electron. Agric. 2018, 149, 110–120. [Google Scholar] [CrossRef]
- Domínguez-Brito, A.C.; Cabrera-Gámez, J.; Viera-Pérez, M.; Rodríguez-Barrera, E.; Hernández-Calvento, L. A DIY low-cost wireless wind data acquisition system used to study an arid coastal foredune. Sensors 2020, 20, 1064. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, K.; Inoue, M.; Shimura, T.; Iguchi, M. In Situ, Rotor-Based Drone measurement of wind vector and aerosol concentration in volcanic areas. Atmosphere 2021, 12, 376. [Google Scholar] [CrossRef]
- Thielicke, W.; Hübert, W.; Müller, U.; Eggert, M.; Wilhelm, P. Towards accurate and practical drone-based wind measurements with an ultrasonic anemometer. Atmos. Meas. Tech. 2021, 14, 1303–1318. [Google Scholar] [CrossRef]
- Yoon, S.; Jang, K.; Won, M. The spatial distribution characteristics of Automatic Weather Stations in the mountainous area over South Korea. Korean J. Agric. For. Meteorol. 2018, 20, 117–126. [Google Scholar] [CrossRef]
- Pounden, E.; Greene, D.; Quesada, M.; Contreras Sánchez, J. The effect of collisions with vegetation elements on the dispersal of winged and plumed seeds. J. Ecol. 2008, 96, 591–598. [Google Scholar] [CrossRef]
- Bohrer, G.; Katul, G.G.; Nathan, R.; Walko, R.L.; Avissar, R. Effects of canopy heterogeneity, seed abscission and inertia on wind-driven dispersal kernels of tree seeds. J. Ecol. 2008, 96, 569–580. [Google Scholar] [CrossRef]
- Esmail, S.; Agrawal, P.; Aly, S. A novel analytical approach for advection diffusion equation for radionuclide release from an area source. Nucl. Eng. Technol. 2020, 52, 819–826. [Google Scholar] [CrossRef]
- Jeong, S.H.; Lee, S.H. Effects of windbreak Forest according to tree species and planting methods based on wind tunnel experiments. For. Sci. Technol. 2020, 16, 188–194. [Google Scholar] [CrossRef]
- Vollsinger, S.; Mitchell, S.J.; Byrne, K.E.; Novak, M.D.; Rudnicki, M. Wind tunnel measurements of crown streamlining and drag relationships for several hardwood species. Can. J. For. Res. 2005, 35, 1238–1249. [Google Scholar] [CrossRef]
- der Weduwen, D.; Ruxton, G.D. Secondary dispersal mechanisms of winged seeds: A review. Biol. Rev. 2019, 94, 1830–1838. [Google Scholar] [CrossRef] [PubMed]
- Schurr, F.M.; Bond, W.J.; Midgley, G.F.; Higgins, S.I. A mechanistic model for secondary seed dispersal by wind and its experimental validation. J. Ecol. 2005, 93, 1017–1028. [Google Scholar] [CrossRef]
- Higgins, S.I.; Nathan, R.; Cain, M.L. Are long-distance dispersal events in plants usually caused by nonstandard means of dispersal? Ecology 2003, 84, 1945–1956. [Google Scholar] [CrossRef]
- Fornara, D.; Dalling, J.W. Post-dispersal removal of seeds of pioneer species from five Panamanian forests. J. Trop. Ecol. 2005, 21, 79–84. [Google Scholar] [CrossRef]
- Kim, G.-T.; Kim, H.-J. Secondary dispersion of several broadleaved tree seeds by wildlife in Mt. Jungwang, Pyeongchang-gun, Korea. Korean J. Environ. Ecol. 2013, 27, 64–70. [Google Scholar]
- Vander Wall, S.B.; Joyner, J.W. Secondary dispersal by the wind of winged pine seeds across the ground surface. Am. Midl. Nat. 1998, 139, 365–373. [Google Scholar] [CrossRef]
- Von der Lippe, M.; Bullock, J.M.; Kowarik, I.; Knopp, T.; Wichmann, M. Human-mediated dispersal of seeds by the airflow of vehicles. PLoS ONE 2013, 8, e52733. [Google Scholar] [CrossRef]
- Liang, W.; Liu, Z.; Liu, M.; Qin, X.; Baskin, C.C.; Baskin, J.M.; Xin, Z.; Wang, Z.; Su, Z.; Zhou, Q. Wing loading, not terminal velocity, is the best parameter to predict capacity of diaspores for secondary wind dispersal. J. Exp. Bot. 2020, 71, 4298–4307. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Madrigal, J.; Candel-Pérez, D.; Jiménez, E.; Moya, D.; de las Heras, J.; Guijarro, M.; Vega, J.A.; Fernández, C.; Hernando, C. Effects of prescribed burning, vegetation treatment and seed predation on natural regeneration of Spanish black pine (Pinus nigra Arn. ssp. salzmannii) in pure and mixed forest stands. For. Ecol. Manag. 2016, 378, 24–30. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Silva-Santos, P.; Fonseca, T.; López-Serrano, F.R.; Tiscar, P.A.; Martínez-García, E.; Andrés, M.; Del Cerro, A. Modelling Spanish black pine postdispersal seed predation in Central-eastern Spain. For. Syst. 2010, 19, 393–403. [Google Scholar] [CrossRef] [Green Version]
- Worthy, F.R.; Law, R.; Hulme, P.E. Modelling the quantitative effects of pre-and post-dispersal seed predation in Pinus sylvestris L. J. Ecol. 2006, 94, 1201–1213. [Google Scholar] [CrossRef]
- Castro, J.; Gómez, J.M.; García, D.; Zamora, R.; Hódar, J.A. Seed predation and dispersal in relict Scots pine forests in southern Spain. Plant Ecol. 1999, 145, 115–123. [Google Scholar] [CrossRef]
- Manso, R.; Pardos, M.; Calama, R. Climatic factors control rodent seed predation in Pinus pinea L. stands in Central Spain. Ann. For. Sci. 2014, 71, 873–883. [Google Scholar] [CrossRef] [Green Version]
- Ruano, I.; Del Peso, C.; Bravo, F. Post-dispersal predation of Pinus pinaster Aiton seeds: Key factors and effects on belowground seed bank. Eur. J. For. Res. 2015, 134, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.S.; Joo, R.W.; Kim, Y.-S. Forest transition in South Korea: Reality, path and drivers. Land Use Policy 2012, 29, 198–207. [Google Scholar] [CrossRef]
- Kim, M.; Kraxner, F.; Son, Y.; Jeon, S.W.; Shvidenko, A.; Schepaschenko, D.; Ham, B.-Y.; Lim, C.-H.; Song, C.; Hong, M. Quantifying impacts of national-scale afforestation on carbon budgets in South Korea from 1961 to 2014. Forests 2019, 10, 579. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Yoon, T.; Han, S.; Kim, S.; Yi, M.; Park, G.; Kim, C.; Son, Y.; Kim, R.; Son, Y. Estimating the carbon dynamics of South Korean forests from 1954 to 2012. Biogeosciences 2014, 11, 4637–4650. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Lee, J.Y.; Song, M. Scientific activities responsible for successful forest greening in Korea. For. Sci. Technol. 2017, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- You, J.W.; Won, H.K.; Han, H.; Kim, H.S. A comparative analysis of management efficiency between natural regeneration and plantation. Korean J. For. Econ. 2020, 27, 85–91. [Google Scholar] [CrossRef]
- Bae, S.W. Structure and tending method for naturally regenerated young Pinus densiflora Sieb. et Zucc. stands. J. Korean Soc. For. Sci. 1994, 83, 50–62. [Google Scholar]
- Kim, J.H.; Yang, H.M.; Jin, G.Z. The pattern of natural regeneration by three different silvicultural systems in a natural deciduous forest. J. Korean For. Soc. 1999, 88, 169–178. [Google Scholar]
- Na, S.J.; Woo, K.S.; Kim, C.S.; Yoon, J.H.; Lee, H.H.; Lee, D.H. Above-ground growth characteristics between naturally regenerated and planted stands of Pinus densiflora for. erecta Uyeki in Gangwon province. J. Korean Soc. For. Sci. 2010, 99, 323–330. [Google Scholar]
- Chung, J.; Kim, H.S.; Chung, S. Seed fructification and seed dispersal characteristics by harvest types in Natural Regeneration Sites of Larix kaempferi. J. Korean Soc. For. Sci. 2021, 110, 295–307. [Google Scholar] [CrossRef]
- Kim, M.; Kraxner, F.; Forsell, N.; Song, C.; Lee, W.-K. Enhancing the provisioning of ecosystem services in South Korea under climate change: The benefits and pitfalls of current forest management strategies. Reg. Environ. Chang. 2021, 21, 6. [Google Scholar] [CrossRef]
- Korea Forest Service. Statistical Yearbook of Forestry; Korea Forest Service: Daejon, Korea, 2021. (In Korean) [Google Scholar]
- Chung, J.; Kim, H. Occurrence and growth characteristics of natural seedlings by harvest type in a Larix kaempferi Forest. J. Korean For. Soc. 2020, 109, 400–412. [Google Scholar] [CrossRef]
- Liu, M.; Xin, Z.; Su, Z.; Zhao, Y.; Li, X.; Liu, Z.; Cony, M.A.; Liang, W.; Qin, X.; Qian, J. A video camera recording method for measuring terminal velocity of seed dispersal by wind. J. For. Res. 2021, 32, 81–90. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing (R Version 4.0. 3, R Foundation for Statistical Computing, Vienna, Austria, 2020). 2021. Available online: https://www.R-project.org/ (accessed on 17 September 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online: http://qgis.osgeo.org (accessed on 24 November 2021).
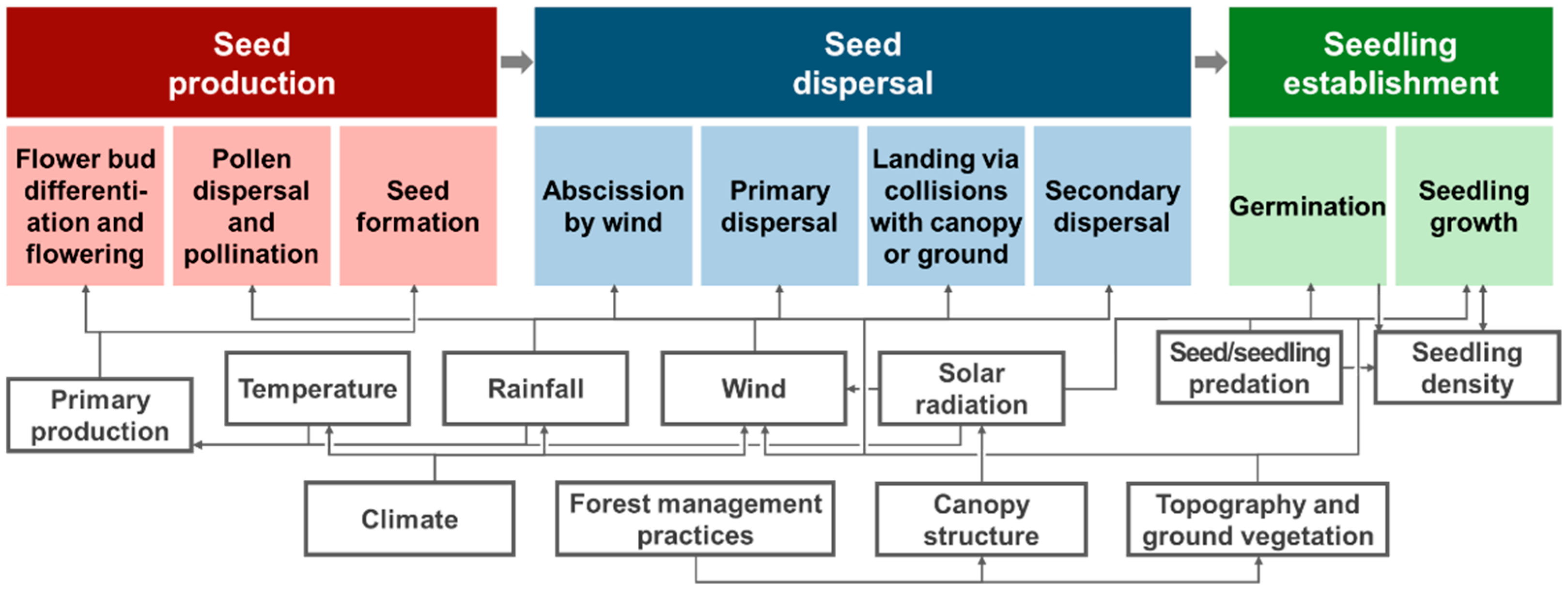
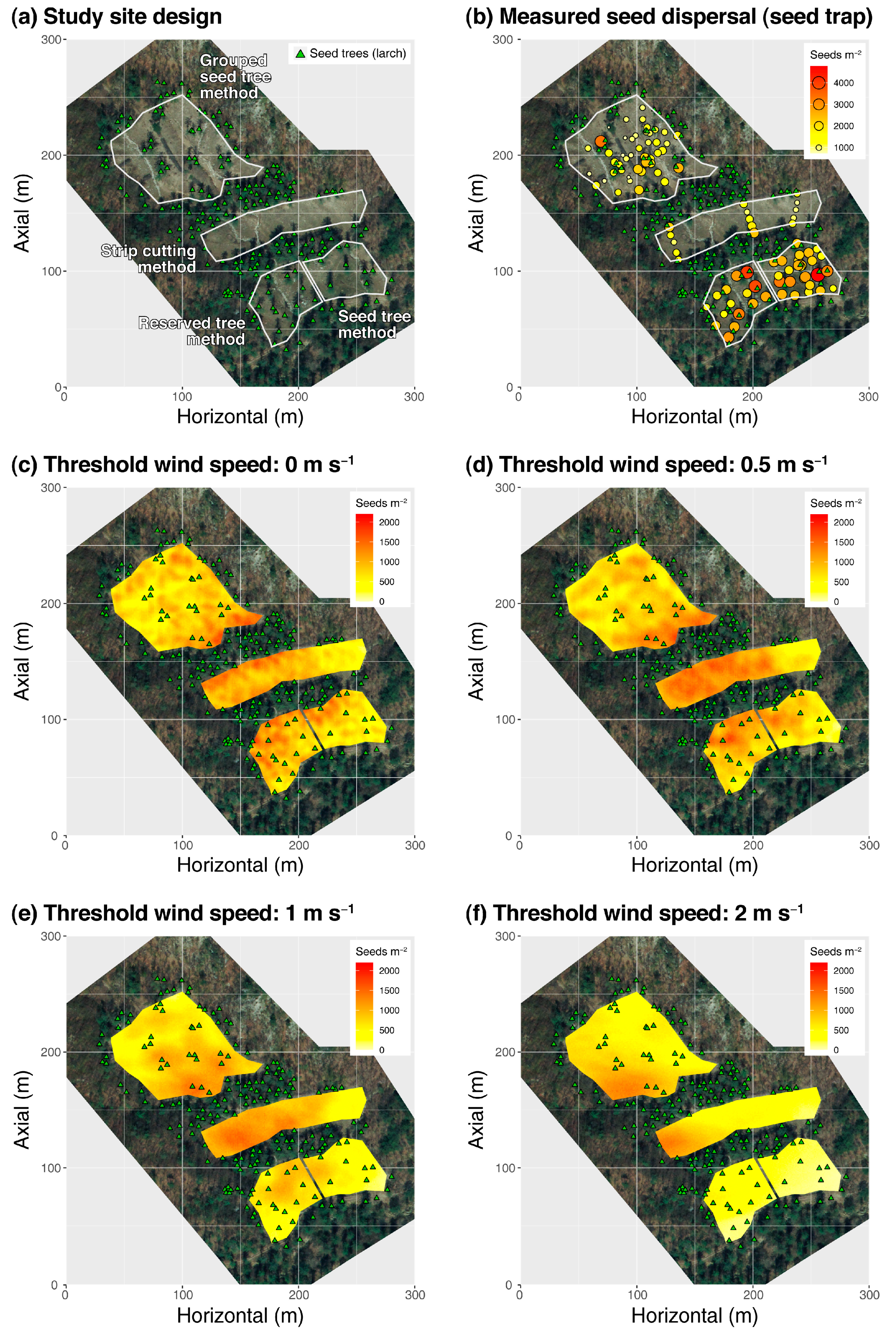
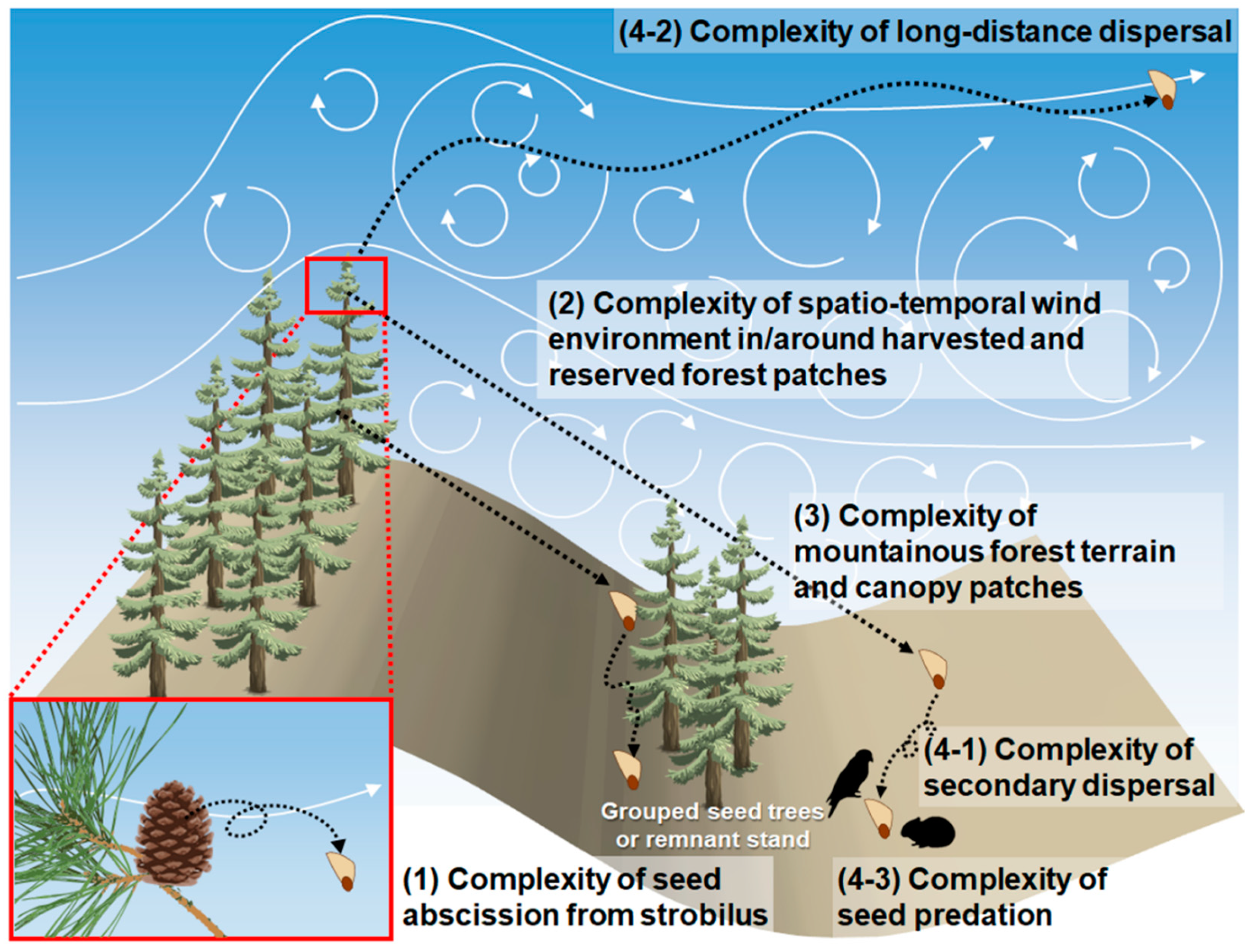
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).