
Leaf Anatomical Plasticity of Phyllostachys glauca McClure in Limestone Mountains Was Associated with Both Soil Water and Soil Nutrients

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Species and Study Site
2.2. Habitat Classification
2.3. Experiment Design and Field Sampling
2.4. Experimental Methods
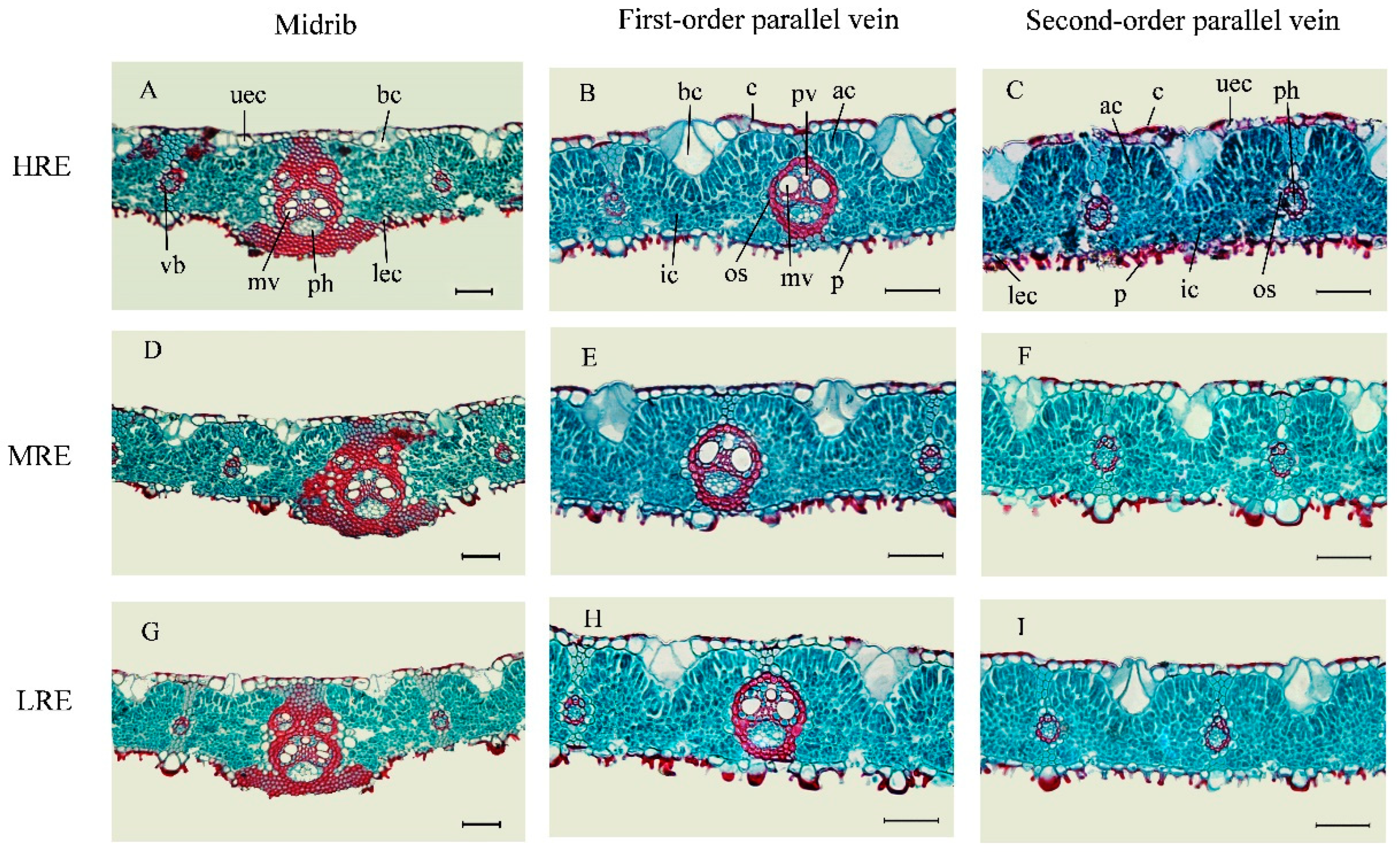
2.4.1. Leaf Anatomical Investigations
2.4.2. Plant Biomass Determination
2.4.3. Soil Water and Nutrient Analysis
2.5. Statistics
3. Results
3.1. Soil Features in Three Habitats
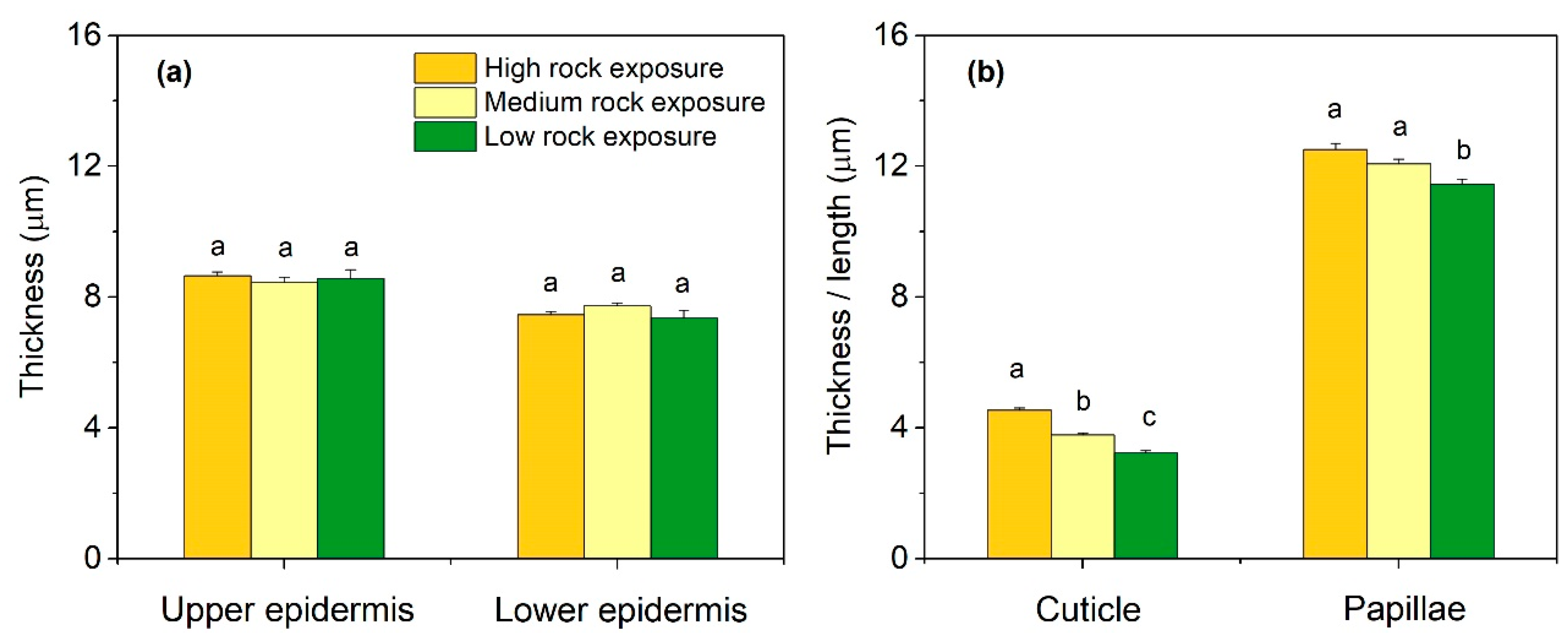
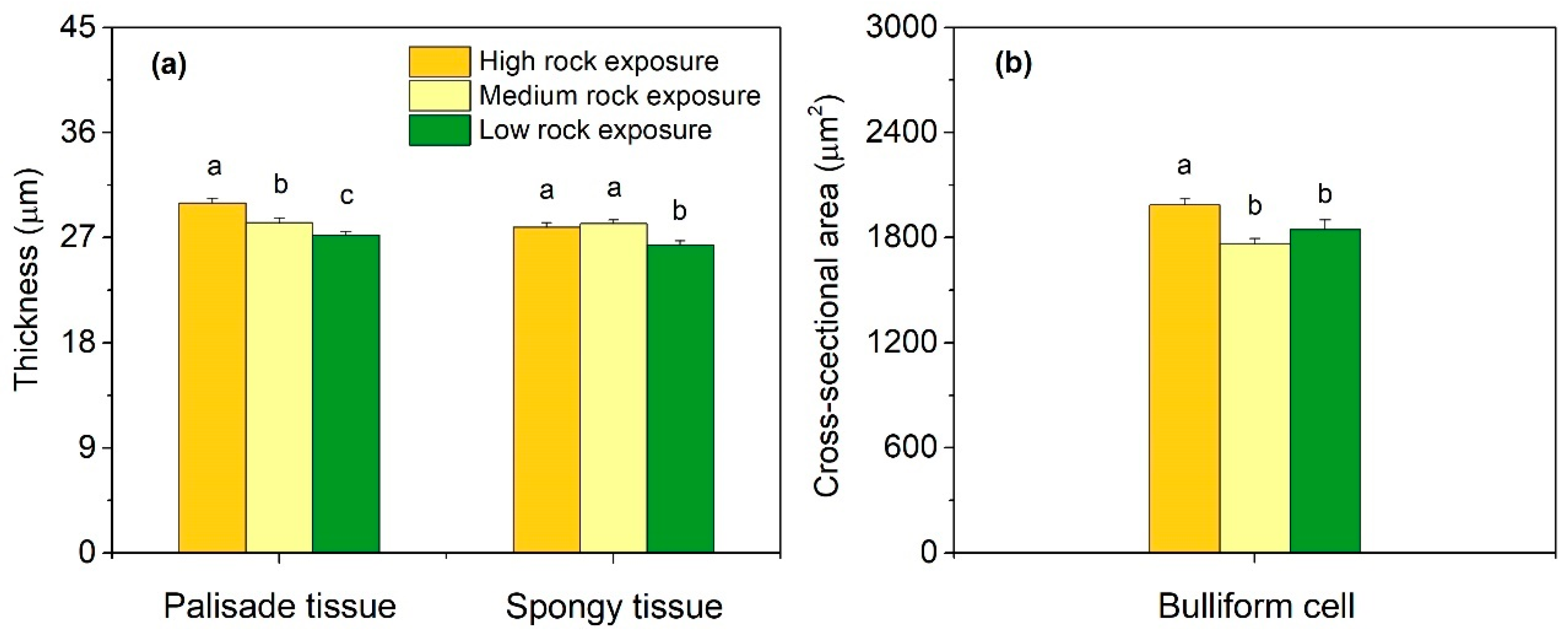
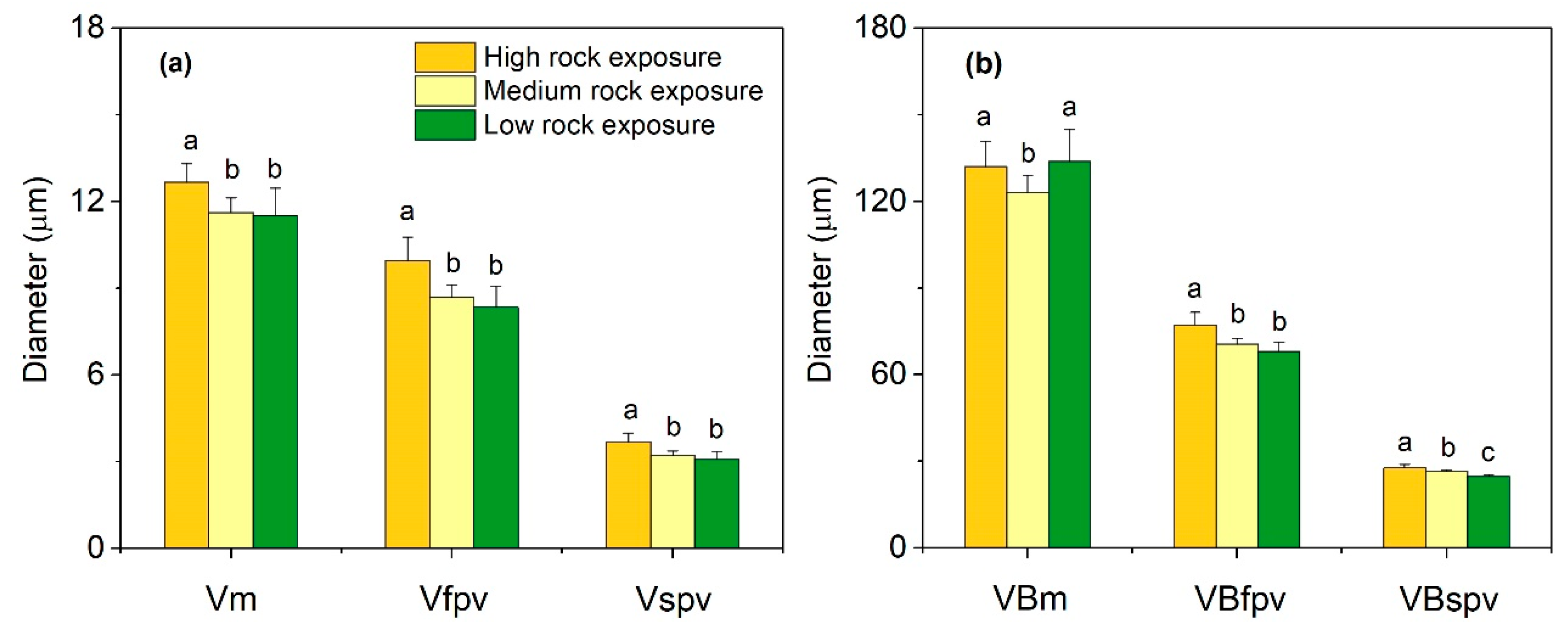
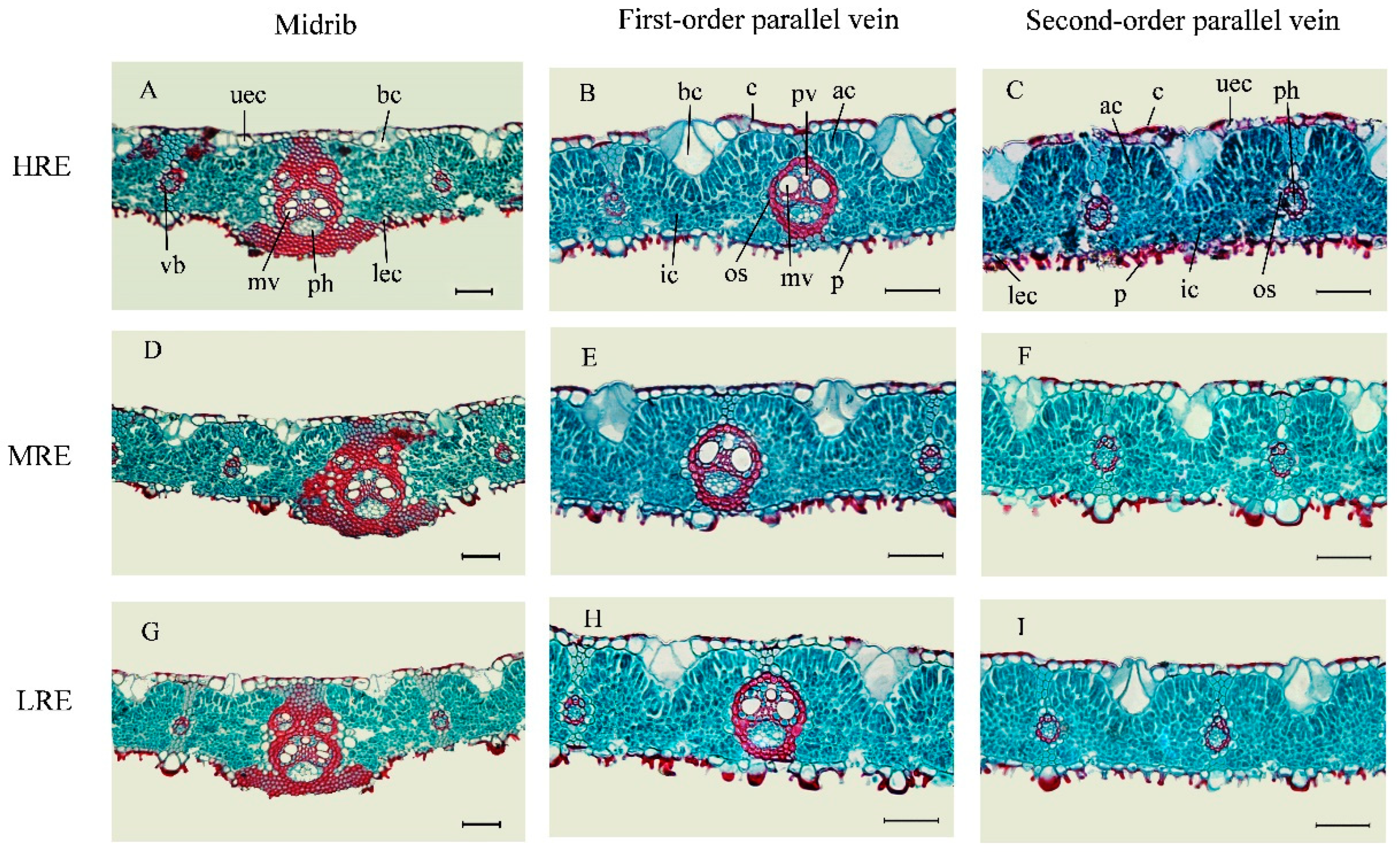
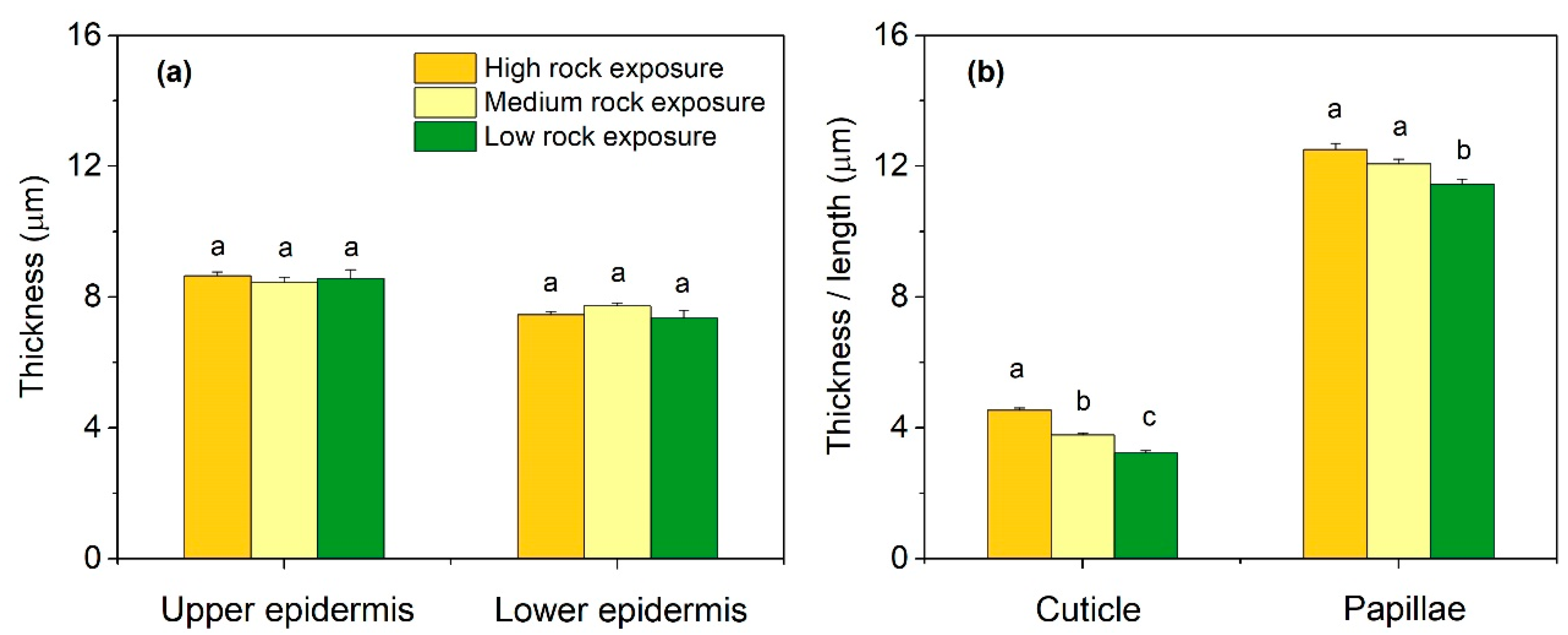
3.2. Leaf Anatomical Features in Three Habitats
3.3. Leaf Anatomical Variations in Three Habitats
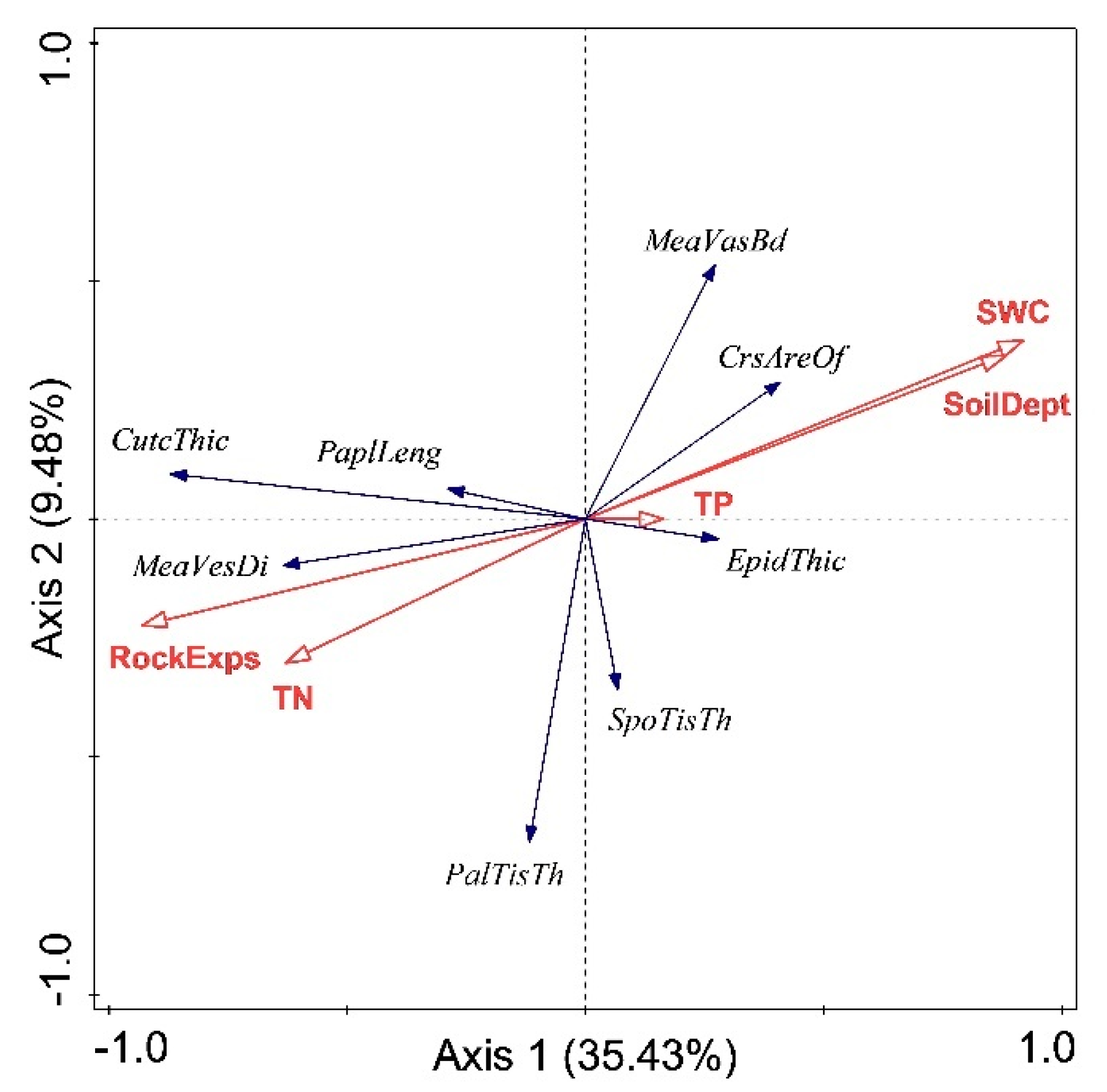
3.4. Relationships between Anatomical Traits and Edaphic Factors
3.5. Relationships between Anatomical Traits and Plant Biomass
4. Discussion
4.1. The Response of Leaf Anatomical Plasticity to Habitat Heterogeneity
4.2. The Effect of Soil Water and Soil Nutrients on Leaf Anatomical Traits
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Global Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Klein, T.; Yakir, D.; Buchmann, N.; Grünzweig, J.M. Towards an advanced assessment of the hydrological vulnerability of forests to climate change-induced drought. New Phytol. 2014, 201, 712–716. [Google Scholar] [CrossRef]
- Findell, K.L.; Berg, A.; Gentine, P.; Krasting, J.P.; Lintner, B.R.; Malyshev, S.; Santanello, J.A.; Shevliakova, E. The impact of anthropogenic land use and land cover change on regional climate extremes. Nat. Commun. 2017, 8, 989. [Google Scholar] [CrossRef]
- Cai, Q.; Ji, C.; Yan, Z.; Jiang, X.; Fang, J. Anatomical responses of leaf and stem of Arabidopsis thaliana to nitrogen and phosphorus addition. J. Plant Res. 2017, 130, 1035–1045. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.; Valladares, F. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Fischer, M. Constraints on the evolution of adaptive phenotypic plasticity in plants. New Phytol. 2005, 166, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Sultan, S.E. Phenotypic plasticity and plant adaptation. Acta Bot. Neerl. 1995, 44, 363–383. [Google Scholar] [CrossRef]
- Dziedek, C.; Fichtner, A.; Calvo, L.; Marcos, E.; Jansen, K.; Kunz, M.; Walmsley, D.; Von Oheimb, G.; Härdtle, W. Phenotypic Plasticity Explains Response Patterns of European Beech (Fagus sylvatica L.) Saplings to Nitrogen Fertilization and Drought Events. Forests 2017, 8, 91. [Google Scholar] [CrossRef] [Green Version]
- Roiloa, S.R.; Retuerto, R. Small-scale heterogeneity in soil quality influences photosynthetic efficiency and habitat selection in a clonal plant. Ann. Bot. 2006, 98, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Bresta, P.; Nikolopoulos, D.; Stavroulaki, V.; Vahamidis, P.; Economou, G.; Karabourniotis, G. How does long-term drought acclimation modify structure-function relationships? A quantitative approach to leaf phenotypic plasticity of barley. Funct. Plant Biol. 2018, 45, 1181–1194. [Google Scholar] [CrossRef]
- Baird, A.S.; Anderegg, L.D.L.; Lacey, M.E.; HilleRisLambers, J.; Van Volkenburgh, E. Comparative leaf growth strategies in response to low-water and low-light availability: Variation in leaf physiology underlies variation in leaf mass per area in Populus tremuloides. Tree Physiol. 2017, 37, 1140–1150. [Google Scholar] [CrossRef] [Green Version]
- Fey, S.B.; Kremer, C.T.; Layden, T.J.; Vasseur, D.A. Resolving the consequences of gradual phenotypic plasticity for populations in variable environments. Ecol. Monogr. 2021, 91, e01478. [Google Scholar] [CrossRef]
- Su, H.; Li, Y.; Lan, Z.; Xu, H.; Liu, W.; Wang, B.; Biswas, D.K.; Jiang, G. Leaf-level plasticity of Salix gordejevii in fixed dunes compared with lowlands in Hunshandake Sandland, North China. J. Plant Res. 2009, 122, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Yu, G.; He, N.; Hou, J. Leaf morphological and anatomical traits from tropical to temperate coniferous forests: Mechanisms and influencing factors. Sci. Rep. 2016, 6, 19703. [Google Scholar] [CrossRef] [Green Version]
- Guan, Z.; Zhang, S.; Guan, K.; Li, S.; Hu, H. Leaf anatomical structures of Paphiopedilum and Cypripedium and their adaptive significance. J. Plant Res. 2011, 124, 289–298. [Google Scholar] [CrossRef]
- Guerfel, M.; Baccouri, O.; Boujnah, D.; Cha?Bi, W.; Zarrouk, M. Impacts of water stress on gas exchange, water relations, chlorophyll content and leaf structure in the two main Tunisian olive (Olea europaea L.) cultivars. Sci. Hortic. 2009, 119, 257–263. [Google Scholar] [CrossRef]
- Haffani, S.; Mezni, M.; Nasri, M.B.; Chaibi, W. Comparative leaf water relations and anatomical responses of three vetch species (Vicia narbonensis L., V. sativa L. and V. villosa Roth.) to cope with water stress. Crop Pasture Sci. 2017, 68, 691–702. [Google Scholar] [CrossRef]
- Façal, B.; Raoudha, A.; Zied, H.; Mohammed, N. Anatomical adaptations of the desert species Stipa lagascae against drought stress. Biologia 2015, 70, 1042–1052. [Google Scholar]
- Wang, Y.; Donovan, L.A.; Temme, A.A. Plasticity and the role of mass-scaling in allocation, morphology, and anatomical trait responses to above- and belowground resource limitation in cultivated sunflower (Helianthus annuus L.). Plant Direct 2020, 4, e00274. [Google Scholar] [CrossRef]
- Dong, X.; Zhang, X. Some observations of the adaptations of sandy shrubs to the arid environment in the Mu Us Sandland: Leaf water relations and anatomic features. J. Arid Environ. 2001, 48, 41–48. [Google Scholar] [CrossRef]
- Bosabalidis, A.M.; Kofidis, G. Comparative effects of drought stress on leaf anatomy of two olive cultivars. Plant Sci. 2002, 163, 375–379. [Google Scholar] [CrossRef]
- Binks, O.; Meir, P.; Rowland, L.; da Costa, A.C.L.; Vasconcelos, S.S.; de Oliveira, A.A.R.; Ferreira, L.; Mencuccini, M. Limited acclimation in leaf anatomy to experimental drought in tropical rainforest trees. Tree Physiol. 2016, 36, 1550–1561. [Google Scholar] [CrossRef] [PubMed]
- Nawazish, S.; Hameed, M.; Naurin, S. Leaf anatomical adaptations of Cenchrus ciliaris L. from the salt range, Pakistan against drought stress. Pak. J. Bot. 2006, 38, 1723–1730. [Google Scholar]
- Burnett, S.E.; Pennisi, S.V.; Thomas, P.A.; Van Iersel, M.W. Controlled drought affects morphology and anatomy of Salvia splendens. J. Am. Soc. Hortic. Sci. 2005, 130, 775–781. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, M.; Schneider, B.; Raveh, E.; Tel-Zur, N. Leaf anatomical characteristics and physiological responses to short-term drought in Ziziphus mauritiana (Lamk.). Sci. Hortic. 2010, 124, 316–322. [Google Scholar] [CrossRef]
- Chartzoulakis, K.; Patakas, A.; Kofidis, G.; Bosabalidis, A.; Nastou, A. Water stress affects leaf anatomy, gas exchange, water relations and growth of two avocado cultivars. Sci. Hortic. 2002, 95, 39–50. [Google Scholar] [CrossRef]
- Grigore, M.N.; Toma, C. Ecological implications of bulliform cells on halophytes, in salt and water stress natural conditions. Studia Universitatis Vasile Goldis Arad, Seria Stiintele Vietii 2011, 21, 785–792. [Google Scholar]
- Zhang, F.; Zhang, K.; Du, C.; Li, J.; Xing, Y.; Yang, L.; Li, Y. Effect of drought stress on anatomical structure and chloroplast ultrastructure in leaves of sugarcane. Sugar Tech 2015, 17, 41–48. [Google Scholar] [CrossRef]
- Dörken, V.M.; Parsons, R.F. Morpho-anatomical studies on the change in the foliage of two imbricate-leaved New Zealand podocarps: Dacrycarpus dacrydioides and Dacrydium cupressinum. Plant Syst. Evol. 2016, 302, 41–54. [Google Scholar] [CrossRef]
- Gessler, A.; Schaub, M.; McDowell, N.G. The role of nutrients in drought-induced tree mortality and recovery. New Phytol. 2017, 214, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Wang, Y.; Hoch, G.; Wang, Z.; Gu, J. Linkage of root morphology to anatomy with increasing nitrogen availability in six temperate tree species. Plant Soil 2018, 425, 189–200. [Google Scholar] [CrossRef]
- Hacke, U.G.; Plavcová, L.; Almeida-Rodriguez, A.; King-Jones, S.; Zhou, W.; Cooke, J.E.K. Influence of nitrogen fertilization on xylem traits and aquaporin expression in stems of hybrid poplar. Tree Physiol. 2010, 30, 1016–1025. [Google Scholar] [CrossRef] [Green Version]
- Plavcová, L.; Hacke, U.G. Phenotypic and developmental plasticity of xylem in hybrid poplar saplings subjected to experimental drought, nitrogen fertilization, and shading. J. Exp. Bot. 2012, 63, 6481–6491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Liu, Y.; Guo, K.; Li, G.; Zheng, Y.; Yu, L.; Yang, R. Comparative ecophysiological responses to drought of two shrub and four tree species from karst habitats of southwestern China. Trees 2011, 25, 537–549. [Google Scholar] [CrossRef]
- Cheng, L.; Liu, Y.; Guo, K.; Fan, D.; Yu, L.; Yang, R. Exploitation of patchy soil water resources by the clonal vine Ficus tikoua in karst habitats of southwestern China. Acta Physiol. Plant. 2011, 33, 93–102. [Google Scholar]
- Geekiyanage, N.; Goodale, U.M.; Cao, K.; Kitajima, K. Plant ecology of tropical and subtropical Karst ecosystems. Biotropica 2019, 51, 626–640. [Google Scholar] [CrossRef]
- Hofmeister, J.; Mihaljevič, M.; Hošek, J.; Sádlo, J. Eutrophication of deciduous forests in the Bohemian Karst (Czech Republic): The role of nitrogen and phosphorus. For. Ecol. Manag. 2002, 169, 213–230. [Google Scholar] [CrossRef]
- Nardini, A.; Battistuzzo, M.; Savi, T. Shoot desiccation and hydraulic failure in temperate woody angiosperms during an extreme summer drought. New Phytol. 2013, 200, 322–329. [Google Scholar] [CrossRef]
- Du, Y.; Pan, G.; Li, L.; Hu, Z.; Wang, X. Leaf N/P ratio and nutrient reuse between dominant species and stands: Predicting phosphorus deficiencies in Karst ecosystems, southwestern China. Environ. Earth Sci. 2011, 64, 299–309. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Guo, K.; Qiao, X.; Zhao, H.; Wang, S.; Zhang, L.; Cai, X. Effects of nitrogen, phosphorus and potassium addition on the productivity of a karst grassland: Plant functional group and community perspectives. Ecol. Eng. 2018, 117, 84–95. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, J.; Pan, F.; Li, D.; Chen, H.; Wang, K. Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in southwest China. Plant Soil 2015, 391, 77–91. [Google Scholar] [CrossRef]
- Liu, C.; Liu, Y.; Guo, K.; Wang, S.; Yang, Y. Concentrations and resorption patterns of 13 nutrients in different plant functional types in the karst region of south-western China. Ann. Bot. 2014, 113, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Liang, K.; Fan, Y.; Bu, W.; Wang, L.; Shi, J. Phenotypic plasticity of a dominant bamboo species (Phyllostachys glauca) in limestone mountain in northwest of Jiangxi province. Acta Agric. Univ. Jiangxiensis 2017, 39, 1178–1186. (In Chinese) [Google Scholar]
- Shi, J.; Mao, S.; Wang, L.; Ye, X.; Wu, J.; Wang, G.; Chen, F.; Yang, Q. Clonal integration driven by source-sink relationships is constrained by rhizome branching architecture in a running bamboo species (Phyllostachys glauca): A 15N assessment in the field. For. Ecol. Manag. 2021, 481, 118754. [Google Scholar] [CrossRef]
- Chen, S.; Li, D.; Zhu, G.; Wu, Z.; Lu, S.; Liu, L.; Wang, Z.; Sun, B.; Zhu, Z.; Xia, N.; et al. Poaceae (Gramineae). In Flora of China; Wu, C.-Y., Raven, P.H., Hong, D.-Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St Louis, MI, USA, 2006; Volume 22, pp. 1–653. [Google Scholar]
- Xiao, J. Bamboo Forest Management in China; Science Press: Beijing, China, 2010. (In Chinese) [Google Scholar]
- Kang, M.; Tao, J.; Wang, J.; Ren, C.; Qi, Q.; Xiang, Q.-Y.; Huang, H. Adaptive and nonadaptive genome size evolution in Karst endemic flora of China. New Phytol. 2014, 202, 1371–1381. [Google Scholar] [CrossRef]
- Lawless, P.J.; Baskin, J.M.; Baskin, C.C. Scale-dependent classification of xeric limestone prairies: Annual or perennial grasslands? Ann. Mo. Bot. Gard. 2006, 93, 455–464. [Google Scholar] [CrossRef]
- Zhou, Y.; He, X.; Xie, Y.; Wang, M.; Wu, M.; Wu, D. Type Classification for Vegetation Restoration of Karst Mountains in Bijie. Sci. Silvae Sin. 2008, 44, 123–128. (In Chinese) [Google Scholar]
- Gao, H. Study on site classification system of steep cultivated land for quitting in Guizhou Province. Res. Soil Water Conserv. 2003, 10, 76–79. (In Chinese) [Google Scholar]
- Price, A.H.; Young, E.M.; Tomos, A.D. Quantitative trait loci associated with stomatal conductance, leaf rolling and heading date mapped in upland rice (Oryza sativa). New Phytol. 1997, 137, 83–91. [Google Scholar] [CrossRef]
- Balsamo, R.A.; Willigen, C.V.; Bauer, A.M.; Farrant, J. Drought tolerance of selected eragrostis species correlates with leaf tensile properties. Ann. Bot. 2006, 97, 985–991. [Google Scholar] [CrossRef] [Green Version]
- Bresta, P.; Nikolopoulos, D.; Economou, G.; Vahamidis, P.; Lyra, D.; Karamanos, A.; Karabourniotis, G. Modification of water entry (xylem vessels) and water exit (stomata) orchestrates long term drought acclimation of wheat leaves. Plant Soil 2011, 347, 179–193. [Google Scholar] [CrossRef]
- Fan, D.; Jie, S.; Liu, C.; Zhang, X.; Xu, X.; Zhang, S.; Xie, Z. The trade-off between safety and efficiency in hydraulic architecture in 31 woody species in a karst area. Tree Physiol. 2011, 31, 865–877. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Chen, H.; Nie, Y.; Wang, K. Dynamic variations in profile soil water on karst hillslopes in Southwest China. Catena 2019, 172, 655–663. [Google Scholar] [CrossRef]
- Tang, B.; Yin, C.; Yang, H.; Sun, Y.; Liu, Q. The coupling effects of water deficit and nitrogen supply on photosynthesis, WUE, and stable isotope composition in Picea asperata. Acta Physiol. Plant. 2017, 39, 148. [Google Scholar] [CrossRef]
- Lenka, S.; Singh, A.K.; Lenka, N.K. Water and nitrogen interaction on soil profile water extraction and ET in maize–wheat cropping system. Agric. Water Manag. 2009, 96, 195–207. [Google Scholar] [CrossRef]
- Sun, L.; Qi, Y.; Dong, Y.; He, Y.; Peng, Q.; Liu, X.; Jia, J.; Guo, S.; Cao, C. Interactions of water and nitrogen addition on soil microbial community composition and functional diversity depending on the inter-annual precipitation in a Chinese steppe. J. Integr. Agric. 2015, 14, 788–799. [Google Scholar] [CrossRef]
- Lenka, S.; Singh, A.K.; Lenka, N.K. Soil water and nitrogen interaction effect on residual soil nitrate and crop nitrogen recovery under maize–wheat cropping system in the semi-arid region of northern India. Agric. Ecosyst. Environ. 2013, 179, 108–115. [Google Scholar] [CrossRef]
- Aulakh, M.S.; Malhi, S.S. Interactions of nitrogen with other nutrients and water: Effect on crop yield and quality, nutrient use efficiency, carbon sequestration, and environmental pollution. Adv. Agron. 2005, 86, 341–409. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitats | Rock Exposure (%) | Soil Depth (m) | Water Content (%) | Total Nitrogen (mg·g−1) | Total Phosphorus (mg·g−1) |
|---|---|---|---|---|---|
| High rock exposure (HRE) | 71.20 ± 9.66 a | 0.18 ± 0.03 c | 9.27 ± 0.56 c | 3.22 ± 0.41 a | 0.38 ± 0.05 a |
| Medium rock exposure (MRE) | 42.53 ± 3.83 b | 0.37 ± 0.03 b | 14.32 ± 1.31 b | 2.36 ± 0.31 b | 0.29 ± 0.02 b |
| Low rock exposure (LRE) | 4.83 ± 0.73 c | 0.75 ± 0.04 a | 23.55 ± 1.28 a | 1.47 ± 0.19 c | 0.38 ± 0.05 a |
| Anatomical Traits | r | p |
|---|---|---|
| Cuticle thickness | 0.588 | 0.096 |
| Upper epidermal thickness | 0.226 | 0.559 |
| Lower epidermal thickness | 0.562 | 0.116 |
| Papillae length | 0.332 | 0.382 |
| Palisade tissue thickness | 0.601 | 0.087 |
| Spongy tissue thickness | 0.414 | 0.268 |
| Cross-sectional area of bulliform cell | 0.660 | 0.053 |
| Vessel diameter of midrib | 0.309 | 0.419 |
| Vessel diameter of first-order parallel vein | 0.761 | 0.017 |
| Vessel diameter of second-order parallel vein | 0.762 | 0.017 |
| Vascular bundle diameter of midrib | 0.001 | 0.998 |
| Vascular bundle diameter of first-order parallel vein | 0.842 | 0.004 |
| Vascular bundle diameter of second-order parallel vein | 0.900 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Fan, Y.; Yu, F.; Shen, Z.; Song, Q.; Zhang, Z.; Hu, S.; Chen, Y.; Shi, J. Leaf Anatomical Plasticity of Phyllostachys glauca McClure in Limestone Mountains Was Associated with Both Soil Water and Soil Nutrients. Forests 2022, 13, 493. https://doi.org/10.3390/f13040493
Wu H, Fan Y, Yu F, Shen Z, Song Q, Zhang Z, Hu S, Chen Y, Shi J. Leaf Anatomical Plasticity of Phyllostachys glauca McClure in Limestone Mountains Was Associated with Both Soil Water and Soil Nutrients. Forests. 2022; 13(4):493. https://doi.org/10.3390/f13040493
Chicago/Turabian StyleWu, Hongyan, Yan Fan, Fen Yu, Zhan Shen, Qingni Song, Zhenye Zhang, Shuzhen Hu, Yongzhen Chen, and Jianmin Shi. 2022. "Leaf Anatomical Plasticity of Phyllostachys glauca McClure in Limestone Mountains Was Associated with Both Soil Water and Soil Nutrients" Forests 13, no. 4: 493. https://doi.org/10.3390/f13040493
APA StyleWu, H., Fan, Y., Yu, F., Shen, Z., Song, Q., Zhang, Z., Hu, S., Chen, Y., & Shi, J. (2022). Leaf Anatomical Plasticity of Phyllostachys glauca McClure in Limestone Mountains Was Associated with Both Soil Water and Soil Nutrients. Forests, 13(4), 493. https://doi.org/10.3390/f13040493