
Competitive Hierarchy Processes Support Mixed Species Regeneration in Strip-Cuts in the British Columbia Interior



Abstract
:1. Introduction
2. Methods
2.1. Study Area and Species
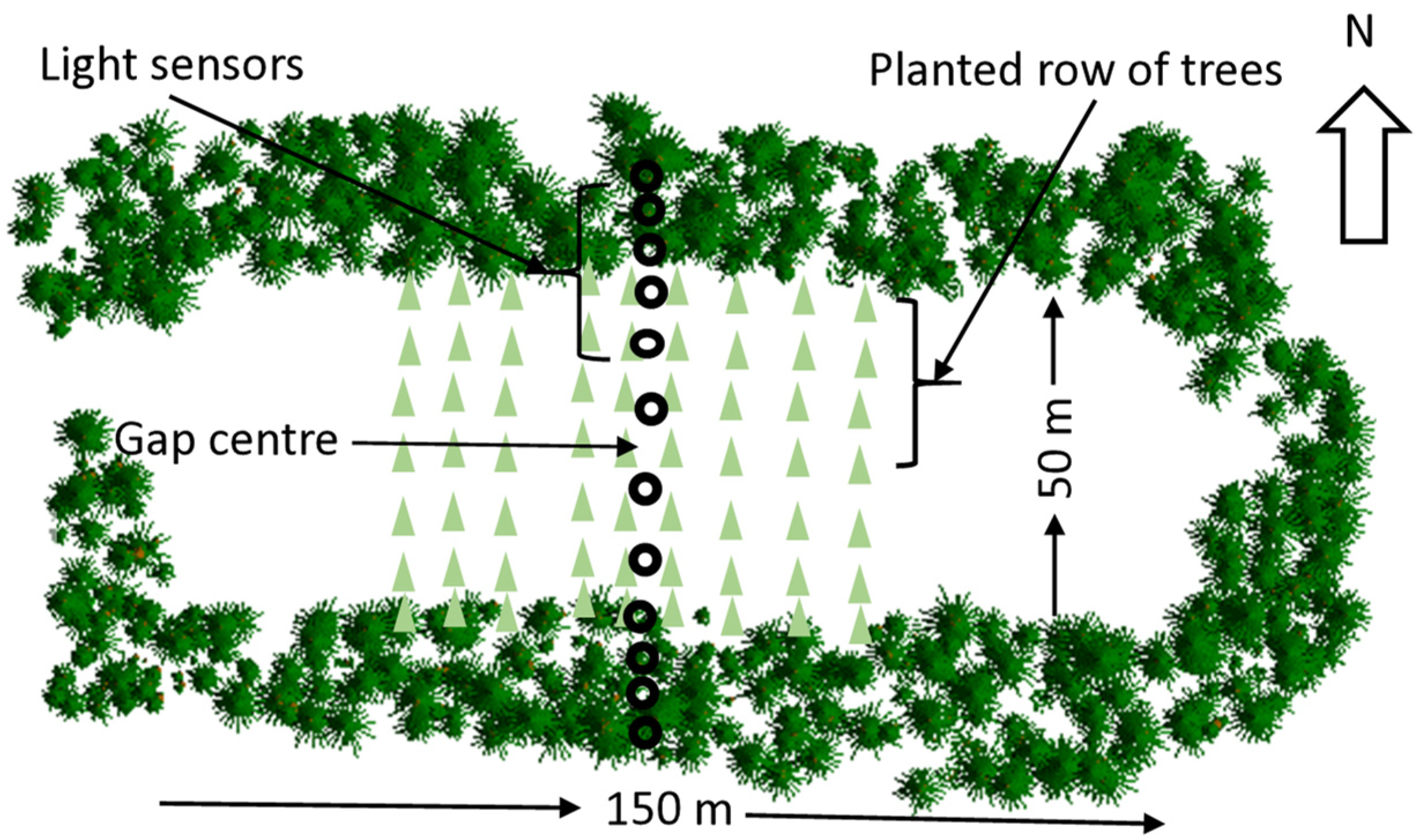
2.2. Experimental Design
2.3. Light Measurements
2.4. Tree Measurements
2.5. Data Analysis
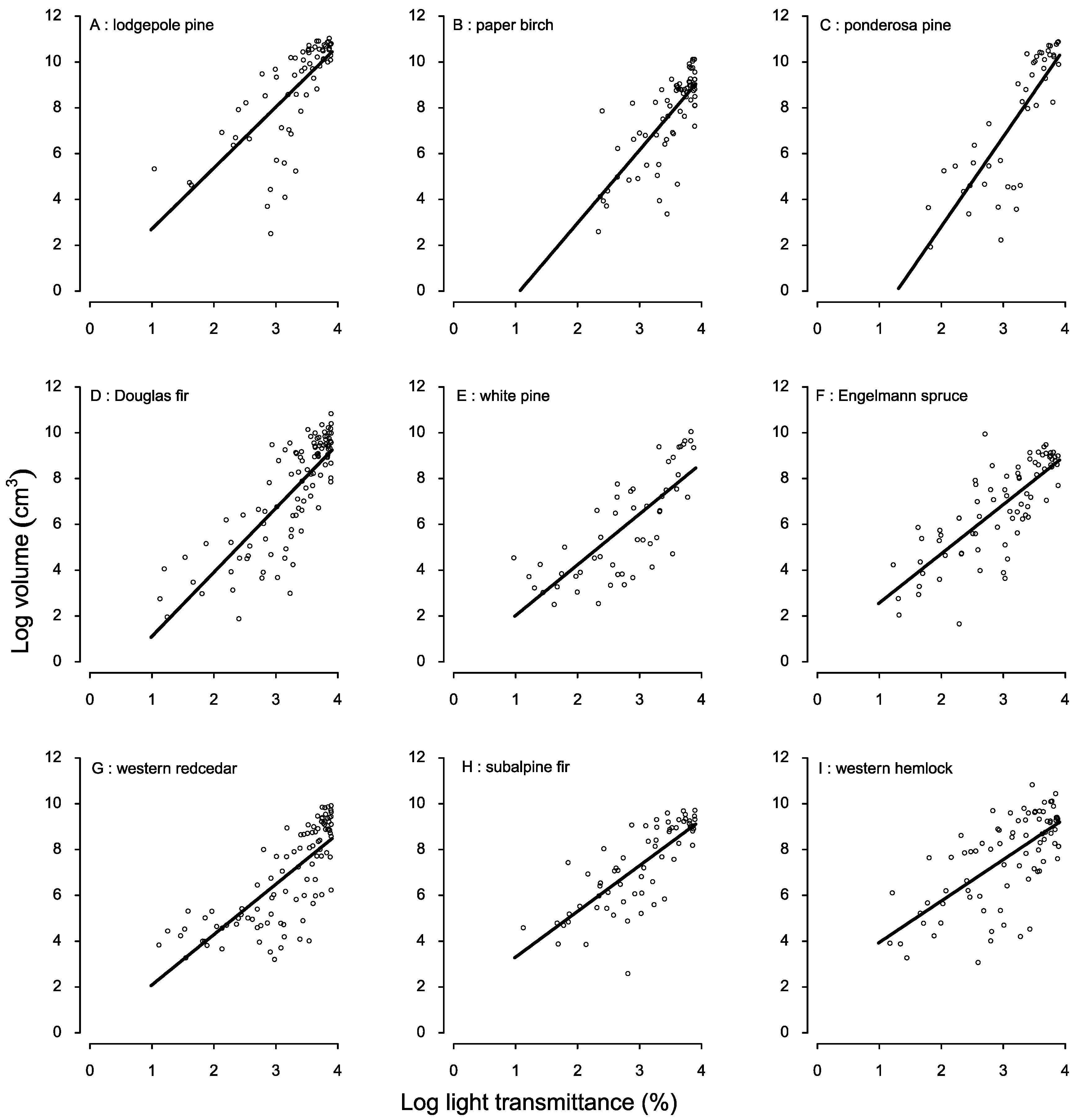
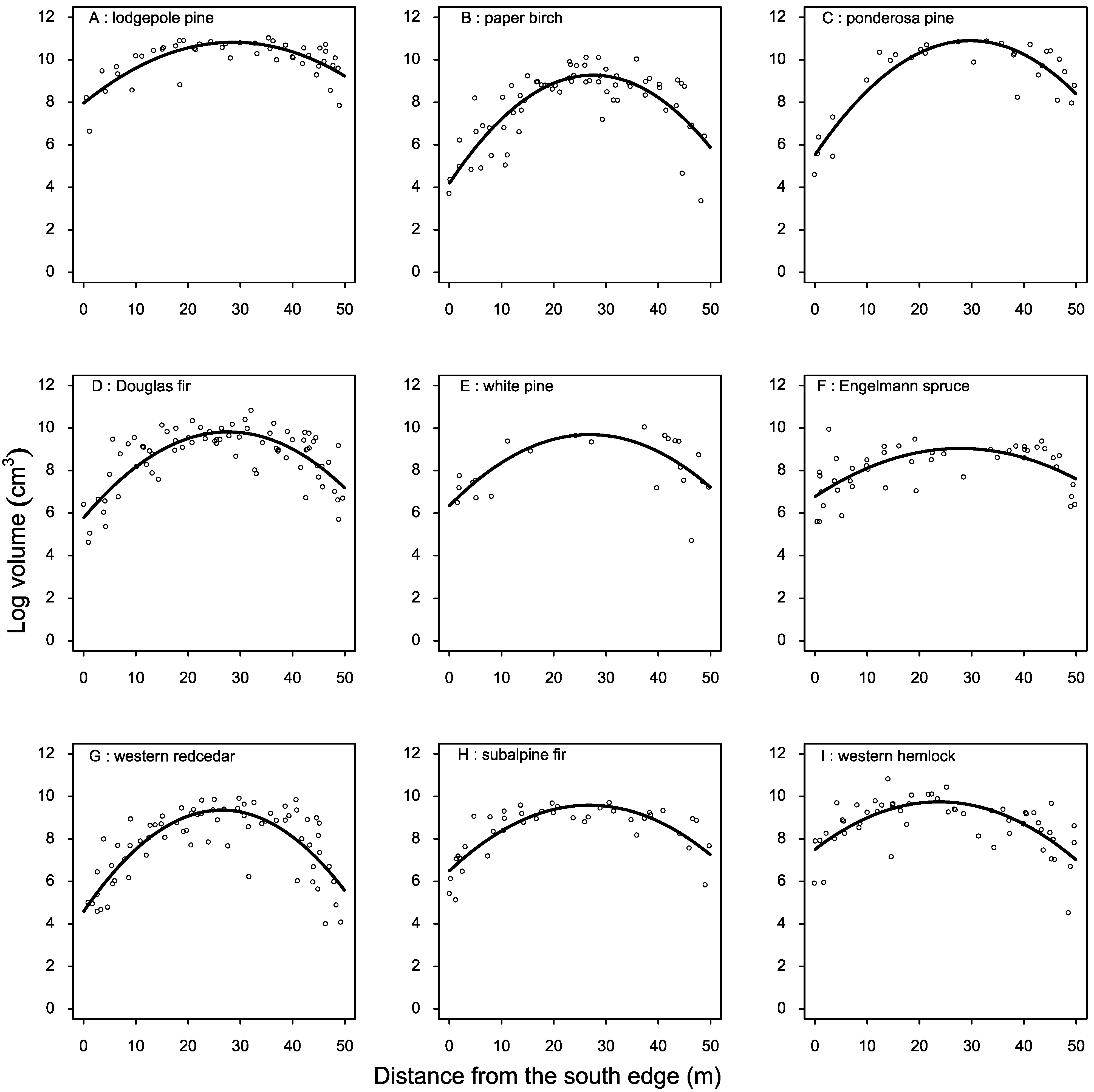
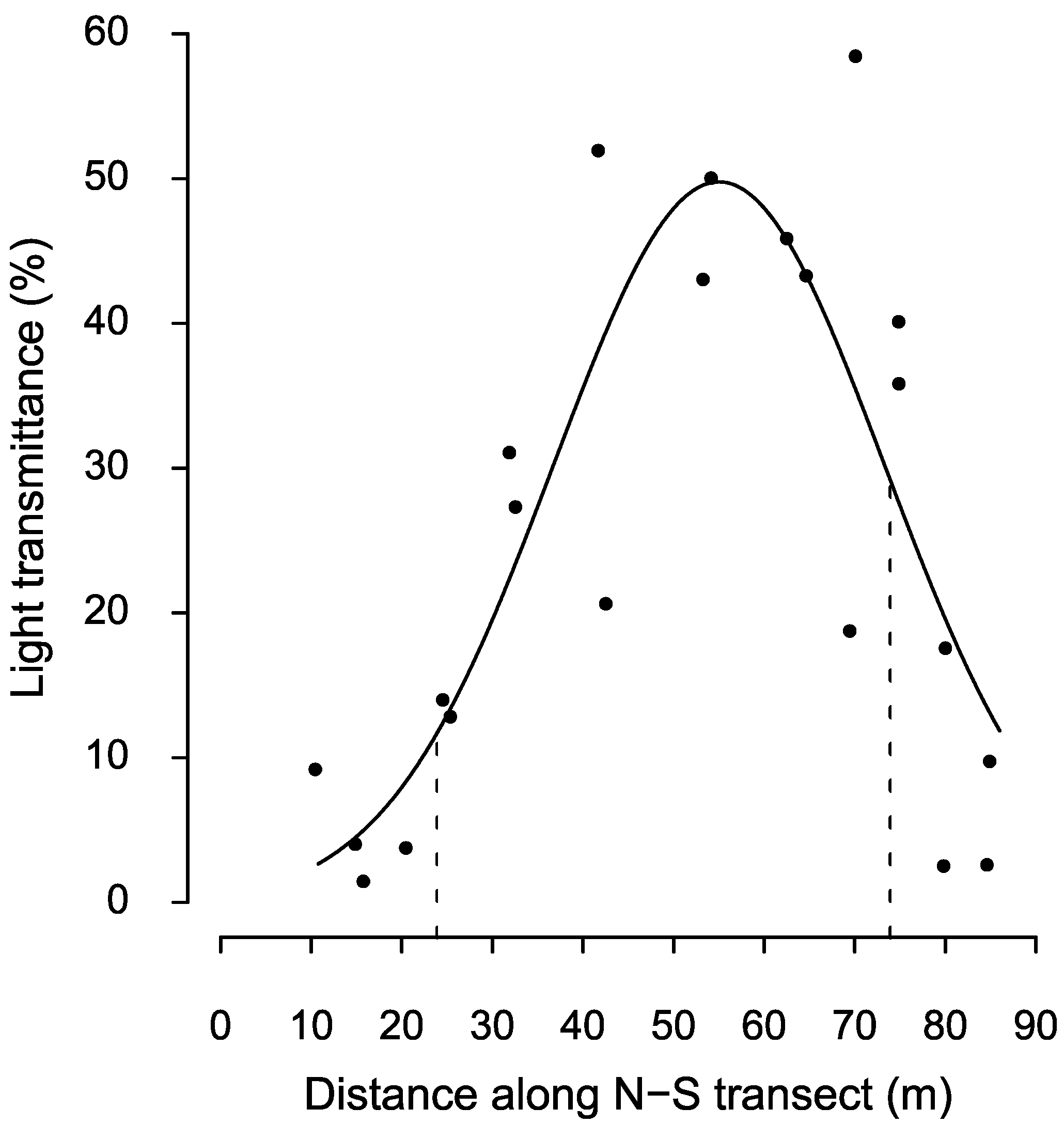
3. Results
3.1. Species-Specific Light-Growth Response
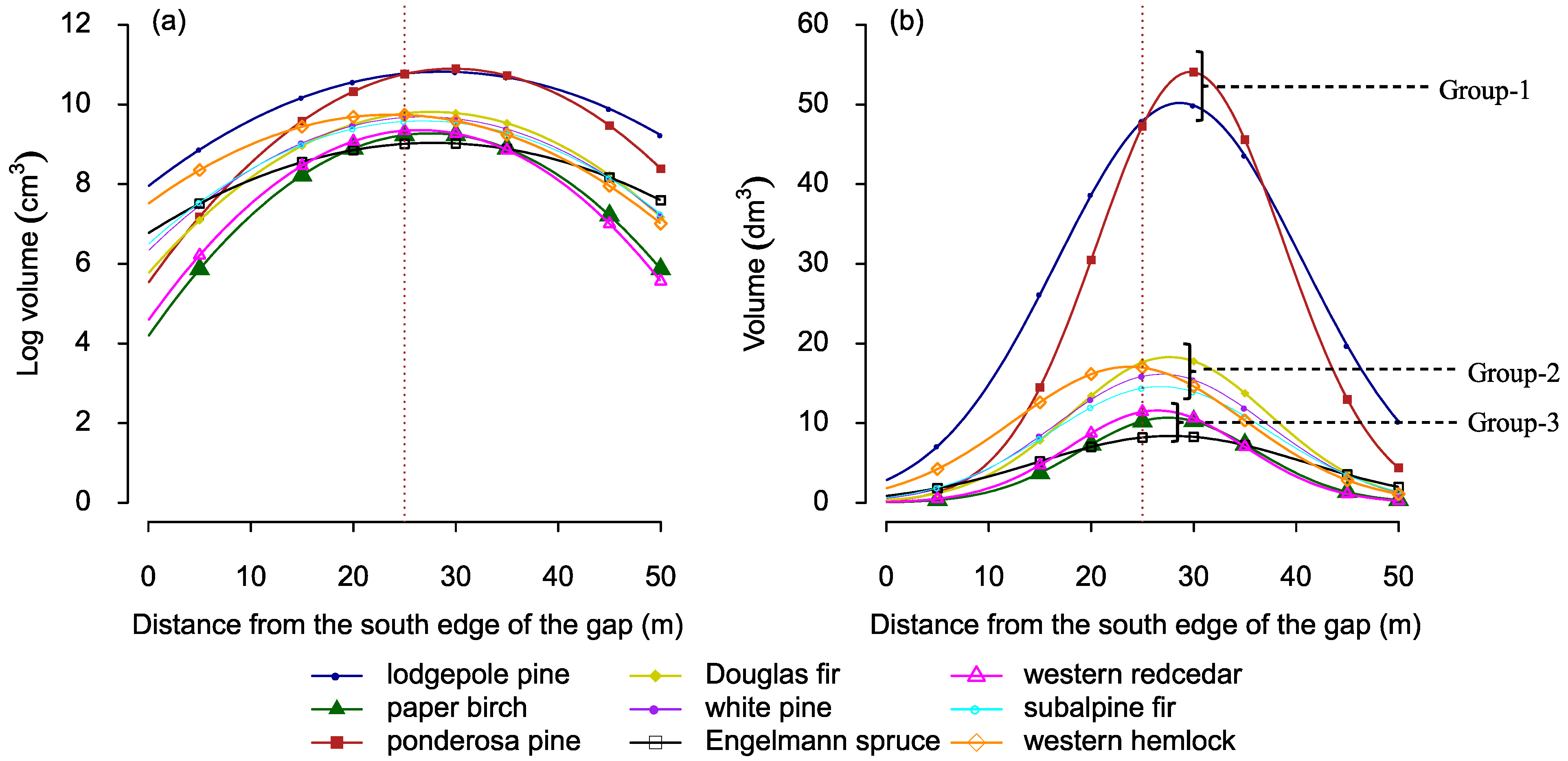
3.2. Growth Response in Relation to Gap-Positions
3.3. Edge Influence
4. Discussion
4.1. Mixed Species Regeneration Appears Feasible
4.2. Species Exhibited Some Evidence of Within Gap-Partitioning
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Newsome, T.A.; Heineman, J.L.; Nemec, A.F.L.; Comeau, P.G.; Arsenault, A.; Waterhouse, M. Ten-year regeneration responses to varying levels of overstory retention in two productive southern British Columbia ecosystems. For. Ecol. Manag. 2010, 260, 132–145. [Google Scholar] [CrossRef]
- DeLong, D.L.; Simard, S.W.; Comeau, P.G.; Dykstra, P.R.; Mitchell, S.J. Survival and growth response of seedlings in root disease infected partial cuts in the Interior Cedar Hemlock zone of southeastern British Columbia. For. Ecol. Manag. 2005, 206, 365–379. [Google Scholar] [CrossRef]
- Kern, C.C.; Burton, J.I.; Raymond, P.; D’Amato, A.W.; Keeton, W.S.; Royo, A.A.; Walters, M.B.; Webster, C.R.; Willis, J.L. Challenges facing gap-based silviculture and possible solutions for mesic northern forests in North America. Forestry 2017, 90, 4–17. [Google Scholar] [CrossRef]
- York, R.A.; Battles, J.J.; Heald, R.C. Edge effects in mixed conifer group selection openings: Tree height response to resource gradients. For. Ecol. Manag. 2003, 179, 107–121. [Google Scholar] [CrossRef] [Green Version]
- Shabaga, J.A.; Jones, T.A.; Elliott, K.A. Group-selection silviculture conditionally enhances recruitment of yellow birch in a shade-tolerant hardwood forest. For. Ecol. Manag. 2019, 444, 244–255. [Google Scholar] [CrossRef]
- Bagnato, S.; Marziliano, P.A.; Sidari, M.; Mallamaci, C.; Marra, F.; Muscolo, A. Effects of Gap Size and Cardinal Directions on Natural Regeneration, Growth Dynamics of Trees outside the Gaps and Soil Properties in European Beech Forests of Southern Italy. Forests 2021, 12, 1563. [Google Scholar] [CrossRef]
- Hossain, K.L.; Comeau, P.G. Characterizing light across a strip shelterwood in a mixed conifer forest. For. Ecol. Manag. 2019, 445, 134–145. [Google Scholar] [CrossRef]
- Sack, L.; Grubb, P.J. Why do species of woody seedlings change rank in relative growth rate between low and high irradiance? Funct. Ecol. 2001, 15, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Van Couwenberghe, R.; Gégout, J.C.; Lacombe, E.; Collet, C. Light and competition gradients fail to explain the coexistence of shade-tolerant Fagus sylvatica and shade-intermediate Quercus petraea seedlings. Ann. Bot. 2013, 112, 1421–1430. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Laanisto, L.; Niinemets, U.; Zavala, M.A. Shedding light on shade: Ecological perspectives of understorey plant life. Plant Ecol. Divers. 2016, 9, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Coates, K.D.; Burton, P.J. Growth of planted tree seedlings in response to ambient light levels in northwestern interior cedar-hemlock forests of British Columbia. Can. J. For. Res. 1999, 29, 1374–1382. [Google Scholar] [CrossRef]
- Kitajima, K.; Bolker, B.M. Testing performance rank reversals among coexisting species: Crossover point irradiance analysis by Sack & Grubb (2001) and alternatives. Funct. Ecol. 2003, 17, 276–281. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Niinemets, Ü. Shade Tolerance, a Key Plant Feature of Complex Nature and Consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- Zdors, L.; Donis, J. Evaluating the edge effect on the initial survival and growth of scots pine and Norway spruce after planting in different size gaps in shelterwood. Balt. For. 2017, 23, 534–543. [Google Scholar]
- Carter, R.E.; Klinka, K. Variation in shade tolerance of Douglas fir, western hemlock, and western red cedar in coastal British Columbia. For. Ecol. Manag. 1992, 55, 87–105. [Google Scholar] [CrossRef]
- Klinka, K.; Wang, Q.; Kayahara, G.J.; Carter, R.E.; Blackwell, B.A. Light-growth response relationships in Pacific silver fir (Abies amabilis) and subalpine fir (Abies lasiocarpa). Can. J. Bot. 1992, 70, 1919–1930. [Google Scholar] [CrossRef]
- Bradshaw, F.J. Quantifying edge effect and patch size for multiple-use silviculture—A discussion paper. For. Ecol. Manag. 1992, 48, 249–264. [Google Scholar] [CrossRef]
- Avalos, G. Shade tolerance within the context of the successional process in tropical rain forests. Rev. Biol. Trop. 2019, 67, 53–77. [Google Scholar] [CrossRef]
- MacKillop, J.D.; Ehman, J.A. A Field Guide to Site Classification and Identification for Southeast British Columbia: The South-Central Columbia Mountains; Province of British Columbia: Victoria, BC, Canada, 2016; 70p. [Google Scholar]
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought, and waterlogging of temperate northern hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Klinka, K.; Chourmouzis, C. Ecological and silvical characteristics of major tree species in British Columbia. In Forestry Handbook for British Columbia, 5th ed.; Watts, S.B., Tolland, L., Eds.; The Forestry Undergraduate Society Faculty of Forestry University of British Columbia: Vancouver, BC, Canada, 2005; ISBN 0-88865-447-2. [Google Scholar]
- Weetman, G.; Vyse, A. Natural regeneration. In Regenerating British Columbia’s Forests; Lavender, D., Parish, R., Johnson, C., Montgomery, G., Vyse, A., Willis, R., Winston, D., Eds.; University of British Columbia Press: Vancouver, BC, Canada, 1990; p. 127. [Google Scholar]
- Haeussler, S.; Coates, D.; Matter, J. Autecology of Common Plants in British Columbia: A Literature Review (No. 158); FRDA Research Program; Research Branch, Ministry of Forests and Lands: Victoria, BC, Canada, 1990; pp. 56–61.
- Fielder, P.; Comeau, P. Construction and Testing of an Inexpensive PAR Sensor; Working Paper; Research Branch, Ministry of Forest: Victoria, BC, Canada, 2000; 53p.
- Honer, T.G.; Ker, M.F.; Alemdag, I.S. Metric Timber Tables for the Commercial Tree Species of Central and Eastern Canada; Information Report No. M–X-140; Canadian Forestry Service-Maritimes: Fredericton, NB, Canada, 1983.
- Pitt, D.G.; Mihajlovich, M.; Proudfoot, L.M. Juvenile stand responses and potential outcomes of conifer release efforts on Alberta’s spruce-aspen mixedwood sites. For. Chron. 2004, 80, 583–597. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; The R Project for Statistical Computing: Vienna, Austria; Available online: http//www.R-project.org/ (accessed on 18 February 2022).
- Pacala, S.W.; Canham, C.D.; Silander Jnr, J.A.; Kobe, R.K. Sapling growth as a function of resources in a north temperate forest. Can. J. For. Res. 1994, 24, 2172–2183. [Google Scholar] [CrossRef]
- Wright, E.F.; Coates, K.D.; Canham, C.D.; Bartemucci, P. Species variability in growth response to light across climatic regions in northwestern British Columbia. Can. J. For. Res. 1998, 28, 871–886. [Google Scholar] [CrossRef]
- Latham, R.E. Co-occurring tree species change rank in seedling performance with resources varied experimentally. Ecology 1992, 73, 2129–2144. [Google Scholar] [CrossRef]
- Claveau, Y.; Messier, C.; Comeau, P.G.; Coates, K.D. Growth and crown morphological responses of boreal conifer seedlings and saplings with contrasting shade tolerance to a gradient of light and height. Can. J. For. Res. 2002, 32, 458–468. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Harcombe, P.A.; Fulton, M.R.; Hall, R.W. Sapling growth and survivorship as a function of light in a mesic forest of southeast Texas, USA. Oecologia 2002, 132, 428–435. [Google Scholar] [CrossRef]
- Gravel, D.; Canham, C.D.; Beaudet, M.; Messier, C. Shade tolerance, canopy gaps and mechanisms of coexistence of forest trees. Oikos 2010, 119, 475–484. [Google Scholar] [CrossRef] [Green Version]
- Burns, R.M.; Honkala, B.H. Silvics of North America. In Agriculture Handbook 654; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990. [Google Scholar]
- Busing, R.T.; Brokaw, N. Tree species diversity in temperate and tropical forest gaps: The role of lottery recruitment. FOLIA Geobot. 2002, 37, 33–43. [Google Scholar] [CrossRef]
- Coates, K.D. Tree recruitment in gaps of various size, clearcuts and undisturbed mixed forest of interior British Columbia, Canada. For. Ecol. Manag. 2002, 155, 387–398. [Google Scholar] [CrossRef]
- Holgén, P.; Hånell, B. Performance of planted and naturally regenerated seedlings in Picea abies-dominated shelterwood stands and clearcuts in Sweden. For. Ecol. Manag. 2000, 127, 129–138. [Google Scholar] [CrossRef]
- De Montigny, L.E.; Smith, N.J. The effects of gap size in a group selection silvicultural system on the growth response of young, planted Douglas-fir: A sector plot analysis. Forestry 2017, 90, 426–435. [Google Scholar] [CrossRef] [Green Version]
- Drever, C.R.; Lertzman, K.P. Light-growth responses of coastal Douglas-fir and western redcedar saplings under different regimes of soil moisture and nutrients. Can. J. For. Res. 2001, 31, 2124–2133. [Google Scholar] [CrossRef]
- Mailly, D.; Kimmins, J.P. Growth of Pseudotsuga menziesii and Tsuga heterophylla seedlings along a light gradient: Resource allocation and morphological acclimation. Can. J. Bot. 1997, 75, 1424–1435. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tolerance Ranking 1 | Silvical Characteristics 2 | |||||
|---|---|---|---|---|---|---|
| Tree Species | Shade | Drought | Suitability for Shelterwood Systems | Suitability for Selection System | Potential for Natural Regen. at Low Light | Potential for Natural Regen. in the Open |
| lodgepole pine | 1.48 | 4.21 | M | L | L | H |
| paper birch | 1.54 | 2.02 | L | L | L | H |
| ponderosa pine | 1.64 | 4.32 | M | L-H | L | H |
| Douglas fir | 2.78 | 2.62 | L-H | L-H | L-H | H |
| white pine | 2.97 | 2.42 | M | L-M | H | L |
| Engelmann spruce | 4.53 | 2.58 | M | M | L | H |
| western redcedar | 4.73 | 2.23 | M | H | M | H |
| subalpine fir | 4.83 | 2.02 | M | H | H | L |
| western hemlock | 4.96 | 1.17 | M | H | H | H |
| Parameters | Chi Square | DF | p Value |
|---|---|---|---|
| Intercept | 2.99 | 1 | 0.083 |
| Log (Light) | 68.12 | 1 | <0.001 |
| Species | 47.22 | 8 | <0.001 |
| Log (Light) × species | 43.41 | 8 | <0.001 |
| Parameters | Chi Square | DF | p Value |
|---|---|---|---|
| Intercept | 1.18 | 1 | 0.278 |
| Species | 60.53 | 8 | <0.001 |
| Gap-position | 45.91 | 1 | <0.001 |
| Gap-position2 | 41.84 | 1 | <0.001 |
| Species × gap-position | 44.84 | 8 | <0.001 |
| Species × gap-position2 | 40.39 | 8 | <0.001 |
| Volume (dm3) and Growth Reductions (%) | |||
|---|---|---|---|
| Species | South Edge | Peak | North Edge |
| lodgepole pine | 8.3 (−83%) | 50.1 | 19.6 (−61%) |
| paper birch | 0.5 (−96%) | 10.7 | 1.4 (−87%) |
| ponderosa pine | 1.8 (−97%) | 56.2 | 13.4 (−76%) |
| Douglas fir | 1.6 (−92%) | 19.5 | 3.7 (−81%) |
| white pine | 2.1 (−87%) | 16.2 | 3.5 (−78%) |
| Engelmann spruce | 2.1 (−75%) | 8.3 | 3.6 (−57%) |
| western redcedar | 0.7 (−94%) | 11.5 | 1.1 (−90%) |
| subalpine fir | 2.2 (−85%) | 15.1 | 3.5 (−77%) |
| western hemlock | 4.9 (−70%) | 16.5 | 2.8 (−83%) |
| Light level (%) | 18 | 50 | 37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, K.L.; Hamann, A.; Comeau, P.G. Competitive Hierarchy Processes Support Mixed Species Regeneration in Strip-Cuts in the British Columbia Interior. Forests 2022, 13, 379. https://doi.org/10.3390/f13030379
Hossain KL, Hamann A, Comeau PG. Competitive Hierarchy Processes Support Mixed Species Regeneration in Strip-Cuts in the British Columbia Interior. Forests. 2022; 13(3):379. https://doi.org/10.3390/f13030379
Chicago/Turabian StyleHossain, Kazi L., Andreas Hamann, and Philip G. Comeau. 2022. "Competitive Hierarchy Processes Support Mixed Species Regeneration in Strip-Cuts in the British Columbia Interior" Forests 13, no. 3: 379. https://doi.org/10.3390/f13030379
APA StyleHossain, K. L., Hamann, A., & Comeau, P. G. (2022). Competitive Hierarchy Processes Support Mixed Species Regeneration in Strip-Cuts in the British Columbia Interior. Forests, 13(3), 379. https://doi.org/10.3390/f13030379