
Combining the Effects of Global Warming, Land Use Change and Dispersal Limitations to Predict the Future Distributions of East Asian Cerris Oaks (Quercus Section Cerris, Fagaceae) in China


Abstract
:1. Introduction
2. Materials and Methods
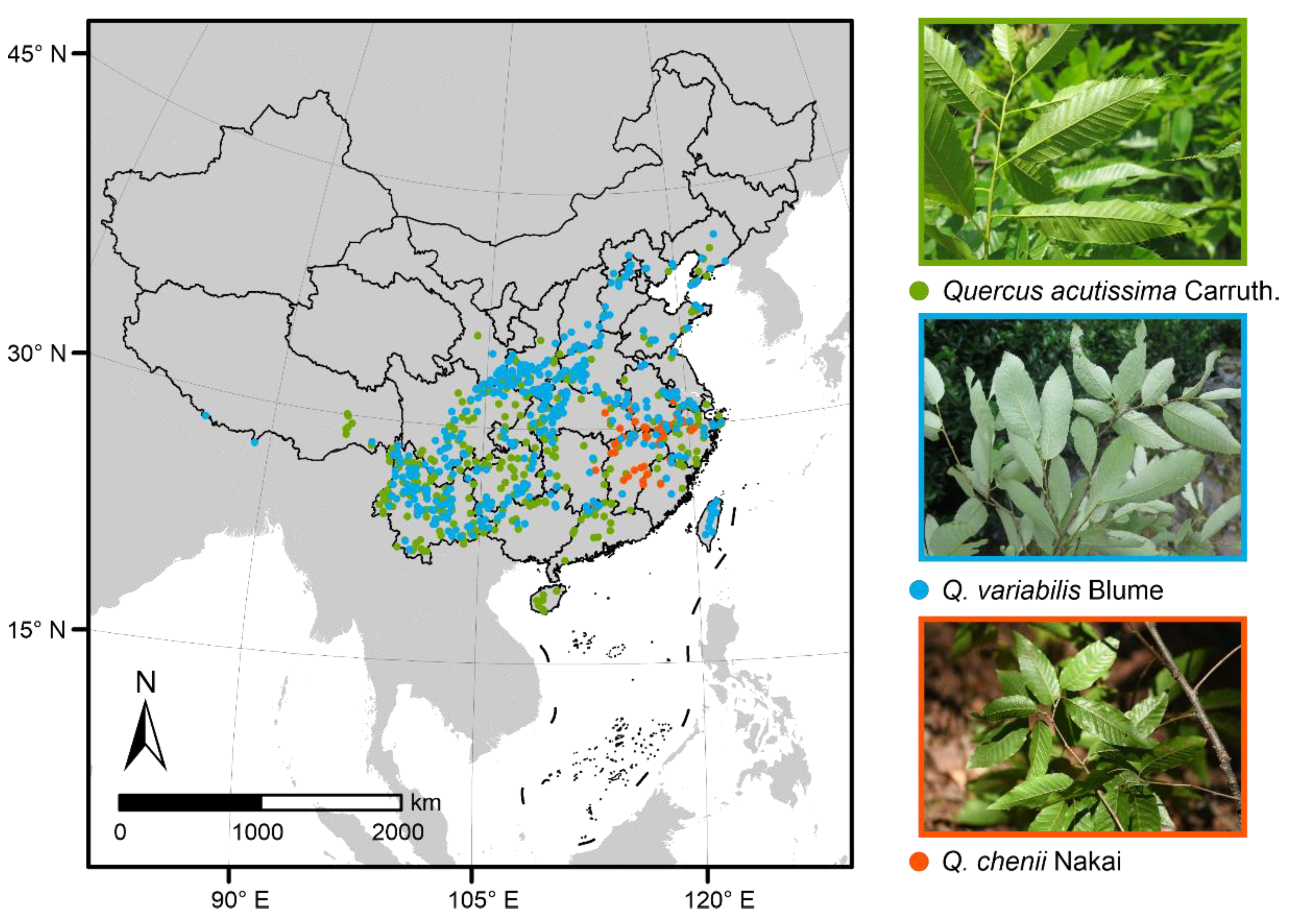
2.1. Species Occurrence Records and Climatic Data
2.2. SDMs Incorporating CC
2.3. SDMs Incorporating LULC
2.4. SDMs Incorporating Dispersal Ability
3. Results
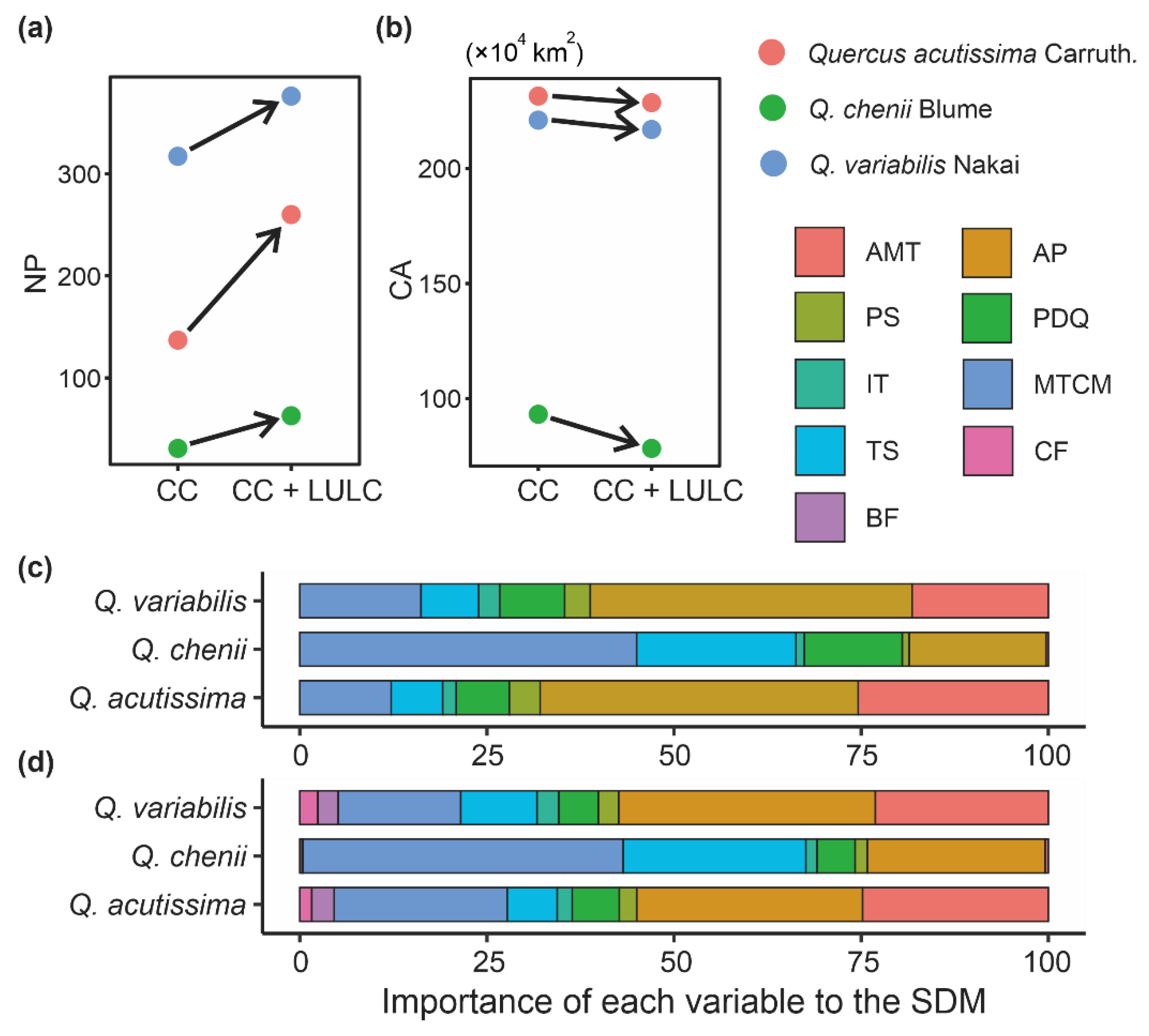
3.1. Model Performance and Key Climatic Factors
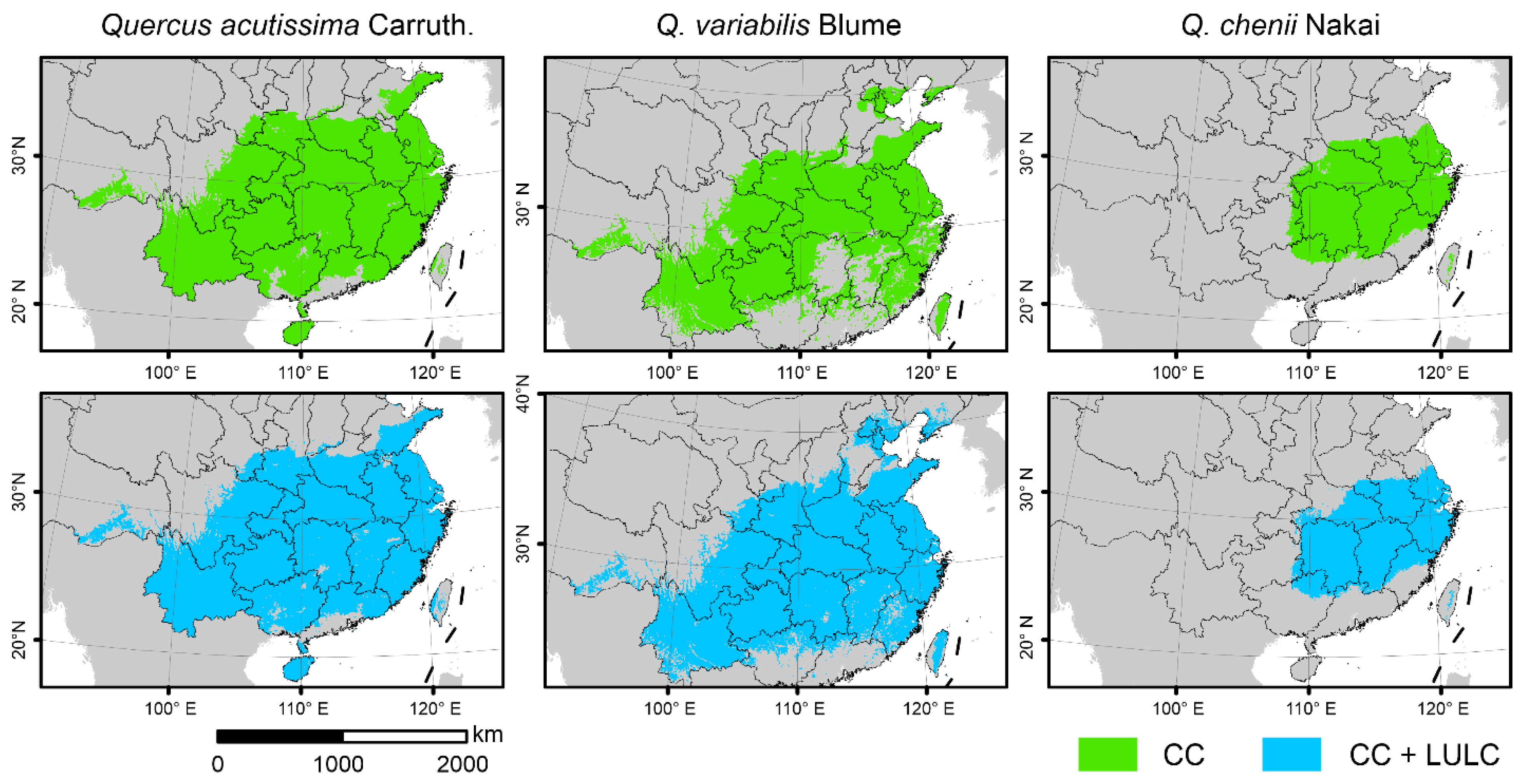
3.2. Sensitivity to LULC in SDMs
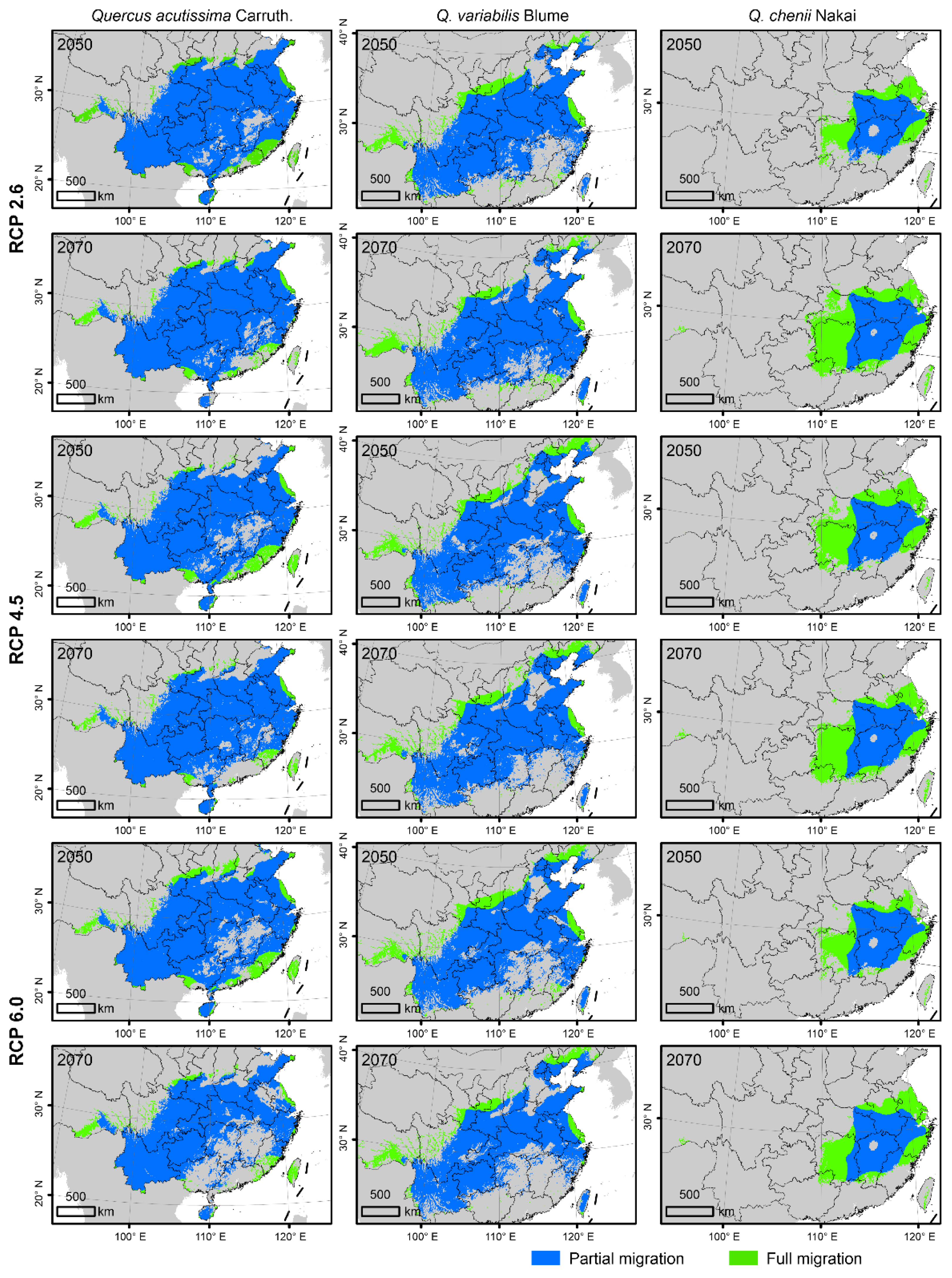
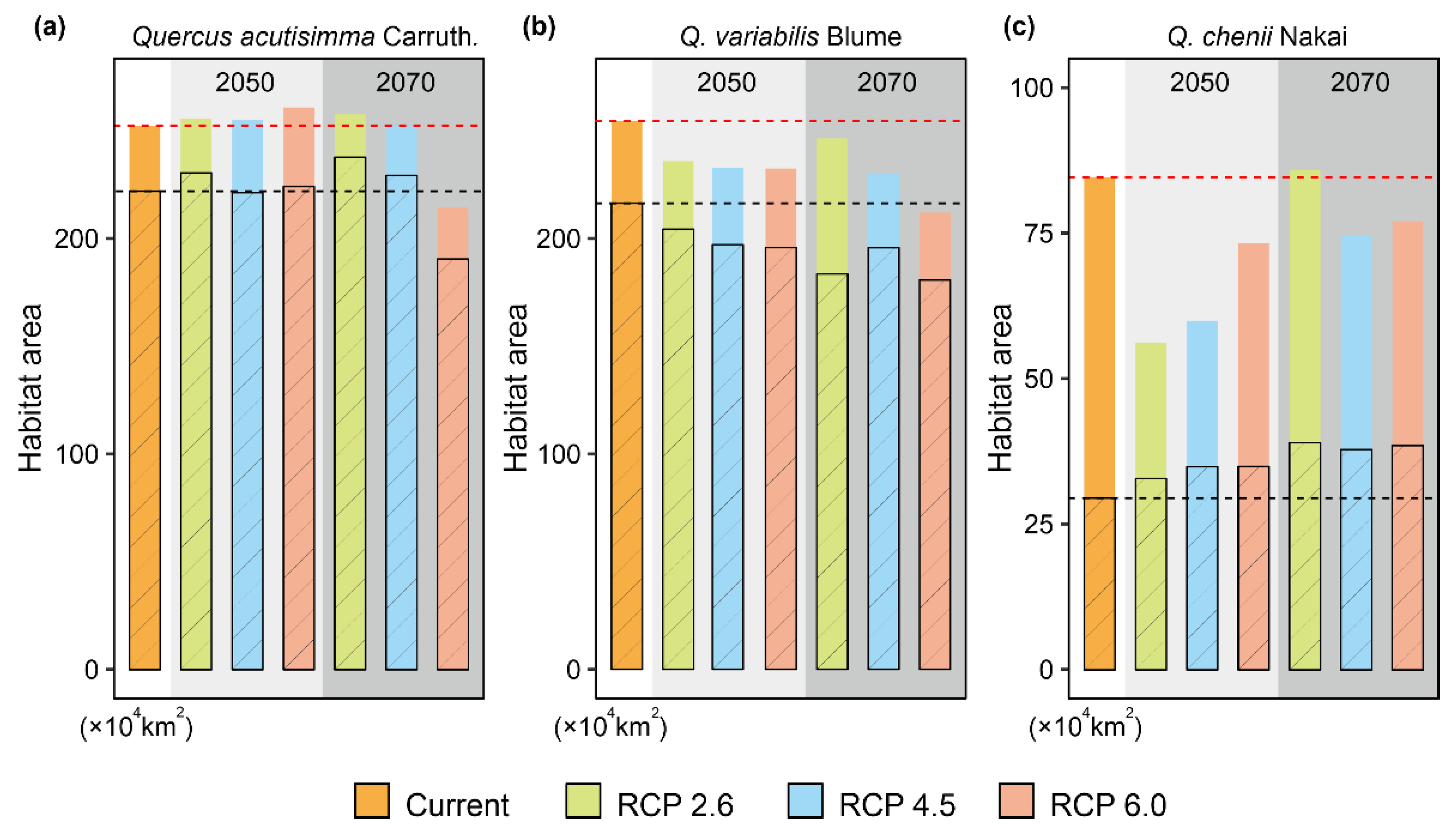
3.3. Projected Future Changes in Species Habitats
4. Discussion
4.1. Key Variables Shaping Species Distributions
4.2. Ecological Niches of East Asian Cerris Oaks
4.3. Future Habitats under Dispersal Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Powers, R.P.; Jetz, W. Global habitat loss and extinction risk of terrestrial vertebrates under future land-use-change scenarios. Nat. Clim. Chang. 2019, 9, 323–329. [Google Scholar] [CrossRef]
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid Range Shifts of Species Associated with High Levels of Climate Warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Lippmann, R.; Babben, S.; Menger, A.; Delker, C.; Quint, M. Development of Wild and Cultivated Plants under Global Warming Conditions. Curr. Biol. 2019, 29, R1326–R1338. [Google Scholar] [CrossRef] [PubMed]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Nicotra, A.; Atkin, O.; Bonser, S.; Davidson, A.; Finnegan, E.; Mathesius, U.; Poot, P.; Purugganan, M.; Richards, C.; Valladares, F.; et al. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef]
- He, X.; Burgess, K.S.; Yang, X.; Ahrends, A.; Gao, L.; Li, D. Upward elevation and northwest range shifts for alpine Meconopsis species in the Himalaya–Hengduan Mountains region. Ecol. Evol. 2019, 9, 4055–4064. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J. Cross-validation of species distribution models: Removing spatial sorting bias and calibration with a null model. Ecology 2012, 93, 679–688. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; Mcdonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2014, 16, 1424–1435. [Google Scholar] [CrossRef]
- Austin, M. Spatial prediction of species distribution: An interface between ecological theory and statistical modelling. Ecol. Model. 2002, 157, 101–118. [Google Scholar] [CrossRef] [Green Version]
- Morán-Ordóñez, A.; Lahoz-Monfort, J.J.; Elith, J.; Wintle, B.A. Evaluating 318 continental-scale species distribution models over a 60-year prediction horizon: What factors influence the reliability of predictions? Glob. Ecol. Biogeogr. 2016, 26, 371–384. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Nobis, M.P.; Normand, S. KISSMig—A simple model for R to account for limited migration in analyses of species distributions. Ecography 2014, 37, 1282–1287. [Google Scholar] [CrossRef]
- Liao, Z.; Zhang, L.; Nobis, M.P.; Wu, X.; Pan, K.; Wang, K.; Dakhil, M.A.; Du, M.; Xiong, Q.; Pandey, B.; et al. Climate change jointly with migration ability affect future range shifts of dominant fir species in Southwest China. Divers. Distrib. 2019, 26, 352–367. [Google Scholar] [CrossRef] [Green Version]
- Subba, B.; Sen, S.; Ravikanth, G.; Nobis, M.P. Direct modelling of limited migration improves projected distributions of Himalayan amphibians under climate change. Biol. Conserv. 2018, 227, 352–360. [Google Scholar] [CrossRef]
- Oliver, T.; Marshall, H.; Morecroft, M.D.; Brereton, T.; Prudhomme, C.; Huntingford, C. Interacting effects of climate change and habitat fragmentation on drought-sensitive butterflies. Nat. Clim. Chang. 2015, 5, 941–945. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Vernon, C.R.; Graham, N.T.; Hejazi, M.; Huang, M.; Cheng, Y.; Calvin, K. Global land use for 2015–2100 at 0.05° resolution under diverse socioeconomic and climate scenarios. Sci. Data 2020, 7, 1–11. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; Nijs, I. Microclimate shifts in a dynamic world. Science 2020, 368, 711–712. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, P.; Hu, W.; Chen, G.; Zhang, D.; Chen, B.; Lei, G. Modeling the Climate Suitability of Northernmost Mangroves in China under Climate Change Scenarios. Forests 2022, 13, 64. [Google Scholar] [CrossRef]
- Holloway, P.; Miller, J.A.; Gillings, S. Incorporating movement in species distribution models: How do simulations of dispersal affect the accuracy and uncertainty of projections? Int. J. Geogr. Inf. Sci. 2016, 30, 2050–2074. [Google Scholar] [CrossRef]
- Li, X.; Yu, L.; Sohl, T.; Clinton, N.; Li, W.; Zhu, Z.; Liu, X.; Gong, P. A cellular automata downscaling based 1 km global land use datasets (2010–2100). Sci. Bull. 2016, 61, 1651–1661. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.; Zhang, J.; Zhang, X.; Li, Y.; Liu, Y.; Wang, Z. Conservation of woody species in China under future climate and land-cover changes. J. Appl. Ecol. 2021, 59, 141–152. [Google Scholar] [CrossRef]
- Sala, O.E.; Chapin, F.S., 3rd; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global Biodiversity Scenarios for the Year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- Chai, Z.; Sun, C.; Wang, D.; Liu, W. Interspecific associations of dominant tree populations in a virgin old-growth oak forest in the Qinling Mountains, China. Bot. Stud. 2016, 57, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Simeone, M.C.; Cardoni, S.; Piredda, R.; Imperatori, F.; Avishai, M.; Grimm, G.W.; Denk, T. Comparative systematics and phylogeography of Quercus Section Cerris in western Eurasia: Inferences from plastid and nuclear DNA variation. PeerJ 2018, 6, e5793. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, X.; Fang, Y. Predicting the impact of global warming on the geographical distribution pattern of Quercus variabilis in China. Chin. J. Appl. Ecol. 2014, 25, 3381–3389. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Fang, Y. Responses of the distribution pattern of Quercus chenii to climate change following the Last Glacial Maximum. Chin. J. Plant Ecol. 2016, 40, 1164–1178. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, Y.; Wang, L.; Yu, H.; Zhang, X.; Liu, P. Resource Investigation and Distribution Pattern of Quercus variabilis Blume and Quercus acutissima Carr. as Woody Energy Plant in China. For. Resour. Manag. 2013, 1, 94–101. [Google Scholar] [CrossRef]
- Zilliox, C.; Gosselin, F. Tree species diversity and abundance as indicators of understory diversity in French mountain forests: Variations of the relationship in geographical and ecological space. For. Ecol. Manag. 2014, 321, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Asbeck, T.; Großmann, J.; Paillet, Y.; Winiger, N.; Bauhus, J. The Use of Tree-Related Microhabitats as Forest Biodiversity Indicators and to Guide Integrated Forest Management. Curr. For. Rep. 2021, 7, 59–68. [Google Scholar] [CrossRef]
- Chen, Y.; Shao, Y.; Xi, J.; Yuan, Z.; Ye, Y.; Wang, T. Community preferences of woody plant species in a heterogeneous temperate forest, China. Front. Ecol. Evol. 2020, 8, 165. [Google Scholar] [CrossRef]
- Chinese Virtual Herbarium Database. Available online: http://www.cvh.ac.cn/ (accessed on 10 January 2022).
- Plant Photo Bank of China. Available online: http://ppbc.iplant.cn/ (accessed on 10 January 2022).
- Getpoint Tool of Baidu Maps. Available online: http://api.map.baidu.com/lbsapi/getpoint/ (accessed on 10 January 2022).
- Senay, S.D.; Worner, S.P.; Ikeda, T. Novel Three-Step Pseudo-Absence Selection Technique for Improved Species Distribution Modelling. PLoS ONE 2013, 8, e71218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ArcGIS 10.3. Available online: http://www.esri.com (accessed on 10 January 2022).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Zurell, D.; Franklin, J.; König, C.; Bouchet, P.J.; Dormann, C.F.; Elith, J.; Fandos, G.; Feng, X.; Guillera-Arroita, G.; Guisan, A.; et al. A standard protocol for reporting species distribution models. Ecography 2020, 43, 1261–1277. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A.; NCEAS Predicting Species Distributions Working Group. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Edwards, T.C., Jr.; Guisan, A.; McMahon, S.M.; Normand, S.; Wilfried Thuiller, W.; Wüest, R.O.; Zimmermann, N.E.; Elith, J. What do we gain from simplicity versus complexity in species distribution models? Ecography 2014, 37, 1267–1281. [Google Scholar] [CrossRef]
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.A.; Stevens, B.; Stouffer, R.J.; Taylor, K.E. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 2016, 9, 1937–1958. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Jiang, Z.; Li, L. Biases and improvements in three dynamical downscaling climate simulations over China. Clim. Dyn. 2016, 47, 3235–3251. [Google Scholar] [CrossRef] [Green Version]
- Franklin, J. Moving beyond static species distribution models in support of conservation biogeography. Divers. Distrib. 2010, 16, 321–330. [Google Scholar] [CrossRef]
- Chen, M.; Vernon, C.R.; Huang, M.; Calvin, K.V.; Kraucunas, I.P. Calibration and analysis of the uncertainty in downscaling global land use and land cover projections from GCAM using Demeter (v1.0.0). Geosci. Model Dev. 2019, 12, 1753–1764. [Google Scholar] [CrossRef] [Green Version]
- Jeremy, V.; Lorena, F.; Stephanie, J.; Luke, S.; Collin, S. SDMTools: Species Distribution Modelling Tools: Tools for Processing Data Associated with Species Distribution Modelling Exercises. Available online: http://www.rforge.net/SDMTools/ (accessed on 20 January 2022).
- Weng, H.; Gao, Y.; Su, X.; Yang, X.; Cheng, F.; Ma, R.; Liu, Y.; Zhang, W.; Zheng, L. Spatial-Temporal Changes and Driving Force Analysis of Green Space in Coastal Cities of Southeast China over the Past 20 Years. Land 2021, 10, 537. [Google Scholar] [CrossRef]
- Wei, Y.; Hoganson, H.M. Landscape impacts from valuing core area in national forest planning. For. Ecol. Manag. 2005, 218, 89–106. [Google Scholar] [CrossRef]
- Brewer, S.; Cheddadi, R.; de Beaulieu, J.; Reille, M. The spread of deciduous Quercus throughout Europe since the last glacial period. For. Ecol. Manag. 2002, 156, 27–48. [Google Scholar] [CrossRef]
- Della Rocca, F.; Milanesi, P. Combining climate, land use change and dispersal to predict the distribution of endangered species with limited vagility. J. Biogeogr. 2020, 47, 1427–1438. [Google Scholar] [CrossRef]
- Ramírez-Preciado, R.P.; Gasca-Pineda, J.; Arteaga, M.C. Effects of global warming on the potential distribution ranges of six Quercus species (Fagaceae). Flora 2018, 251, 32–38. [Google Scholar] [CrossRef]
- Xu, Z.; Huang, F.; Liu, Q.; Fu, C. Global pattern of historical and future changes in rapid temperature variability. Environ. Res. Lett. 2020, 15, 124073. [Google Scholar] [CrossRef]
- Alegria, C.; Roque, N.; Albuquerque, T.; Gerassis, S.; Fernandez, P.; Ribeiro, M.M. Species Ecological Envelopes under Climate Change Scenarios: A Case Study for the Main Two Wood-Production Forest Species in Portugal. Forests 2020, 11, 880. [Google Scholar] [CrossRef]
- Zhu, H.; Jiang, Z.; Li, J.; Li, W.; Sun, C.; Li, L. Does CMIP6 Inspire More Confidence in Simulating Climate Extremes over China? Adv. Atmos. Sci. 2020, 37, 1119–1132. [Google Scholar] [CrossRef]
- Dhyani, S.; Kadaverugu, R.; Pujari, P. Predicting impacts of climate variability on Banj oak (Quercus leucotrichophora A. Camus) forests: Understanding future implications for Central Himalayas. Reg. Environ. Chang. 2020, 20, 1–13. [Google Scholar] [CrossRef]
- Preißer, M.; Bucher, S. Is the Seasonal Variation in Frost Resistance and Plant Performance in Four Oak Species Affected by Changing Temperatures? Forests 2021, 12, 369. [Google Scholar] [CrossRef]
- Ma, L.; Mi, C.; Qu, J.; Ge, D.; Yang, Q.; Wilcove, D.S. Predicting range shifts of pikas (Mammalia, Ochotonidae) in China under scenarios incorporating land use change, climate change and dispersal limitations. Divers. Distrib. 2021, 27, 2384–2396. [Google Scholar] [CrossRef]
- Rathore, P.; Roy, A.; Karnatak, H. Assessing the vulnerability of Oak (Quercus) forest ecosystems under projected climate and land use land cover changes in Western Himalaya. Biodivers. Conserv. 2018, 28, 2275–2294. [Google Scholar] [CrossRef]
- Chung, M.G.; Myers, E.R.; Chung, J.M.; Kim, K.-J.; Park, C.-W.; Sun, B.-Y.; Pak, J.-H.; Chung, M.G. Spatial Genetic Structure in Disturbed Populations of Quercus acutissima (Fagaceae). Silvae Genet. 2004, 53, 206–211. [Google Scholar] [CrossRef]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.; Thuiller, W.; Fortin, M.-J.; Randin, C.F.; Zimmermann, N.; et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2011, 21, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Zhou, S.; Yin, X.; Zhang, C.; Li, R.; Chen, J.; Ma, D.; Wang, Y.; Yu, Z.; Chen, Y. Patterns of tree species richness in Southwest China. Environ. Monit. Assess. 2021, 193, 1–13. [Google Scholar] [CrossRef]
- Li, P.; Liu, C.; Cao, H. Quantitative Evaluation of Ecological Stress Caused by Land Use Transitions Considering the Location of Incremental Construction Lands: The Case of Southern Jiangsu in Yangtze River Delta Region. Land 2022, 11, 175. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Wang, L.; Sork, V.L.; Mao, L.; Fang, Y. Influence of Pliocene and Pleistocene climates on hybridization patterns between two closely related oak species in China. Ann. Bot. 2021, 129, 231–245. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Fang, Y. Landscape Features and Climatic Forces Shape the Genetic Structure and Evolutionary History of an Oak Species (Quercus chenii) in East China. Front. Plant Sci. 2019, 10, 1060. [Google Scholar] [CrossRef]
- Axer, M.; Schlicht, R.; Kronenberg, R.; Wagner, S. The Potential for Future Shifts in Tree Species Distribution Provided by Dispersal and Ecological Niches: A Comparison between Beech and Oak in Europe. Sustainability 2021, 13, 13067. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | RCPs | Q. acutissima | Q. variabilis | Q. chenii |
|---|---|---|---|---|
| current | 228.66 | 217.00 | 78.33 | |
| 2050 | RCP 2.6 | 221.22 | 189.32 | 48.41 |
| RCP 4.5 | 208.98 | 180.66 | 64.86 | |
| RCP 6.0 | 215.46 | 179.98 | 51.74 | |
| 2070 | RCP 2.6 | 223.45 | 198.48 | 75.41 |
| RCP 4.5 | 207.96 | 178.80 | 68.32 | |
| RCP 6.0 | 165.42 | 158.95 | 66.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Li, Y.; Mao, L. Combining the Effects of Global Warming, Land Use Change and Dispersal Limitations to Predict the Future Distributions of East Asian Cerris Oaks (Quercus Section Cerris, Fagaceae) in China. Forests 2022, 13, 367. https://doi.org/10.3390/f13030367
Chen Y, Li Y, Mao L. Combining the Effects of Global Warming, Land Use Change and Dispersal Limitations to Predict the Future Distributions of East Asian Cerris Oaks (Quercus Section Cerris, Fagaceae) in China. Forests. 2022; 13(3):367. https://doi.org/10.3390/f13030367
Chicago/Turabian StyleChen, Yuheng, Yao Li, and Lingfeng Mao. 2022. "Combining the Effects of Global Warming, Land Use Change and Dispersal Limitations to Predict the Future Distributions of East Asian Cerris Oaks (Quercus Section Cerris, Fagaceae) in China" Forests 13, no. 3: 367. https://doi.org/10.3390/f13030367
APA StyleChen, Y., Li, Y., & Mao, L. (2022). Combining the Effects of Global Warming, Land Use Change and Dispersal Limitations to Predict the Future Distributions of East Asian Cerris Oaks (Quercus Section Cerris, Fagaceae) in China. Forests, 13(3), 367. https://doi.org/10.3390/f13030367