
Down-Regulation of Photosynthesis to Elevated CO2 and N Fertilization in Understory Fraxinus rhynchophylla Seedlings

 ,
,
Abstract
:1. Introduction
- Down-regulation of photosynthesis under eCO2 occurs in low N availability.
- Leaf N allocation changes with different N and CO2 concentrations.
- Dilution occur other organs changes with different N availability under eCO2.
2. Materials and Methods
2.1. Study Site
2.2. Leaf Gas Exchange Measurements and Sample Collection
2.3. Total Nonstructural Carbohydrates
2.4. Measurement of Rubisco and Chlorophyll Contents
2.5. Statistical Analysis
3. Results
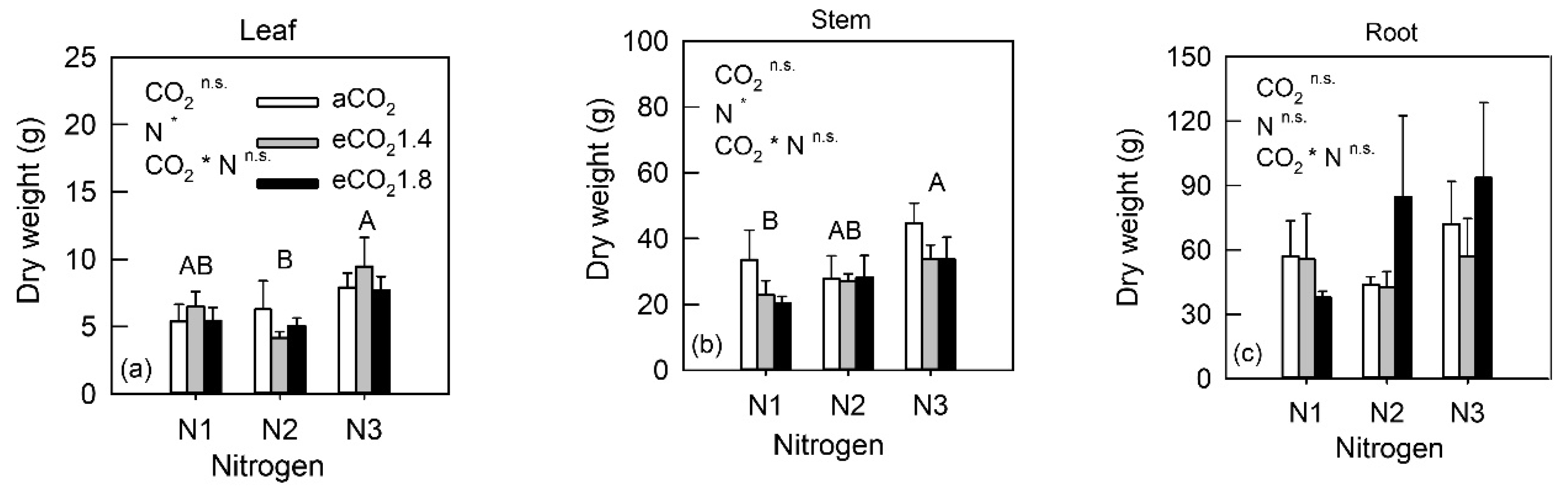
3.1. Whole Plant Biomass Allocation
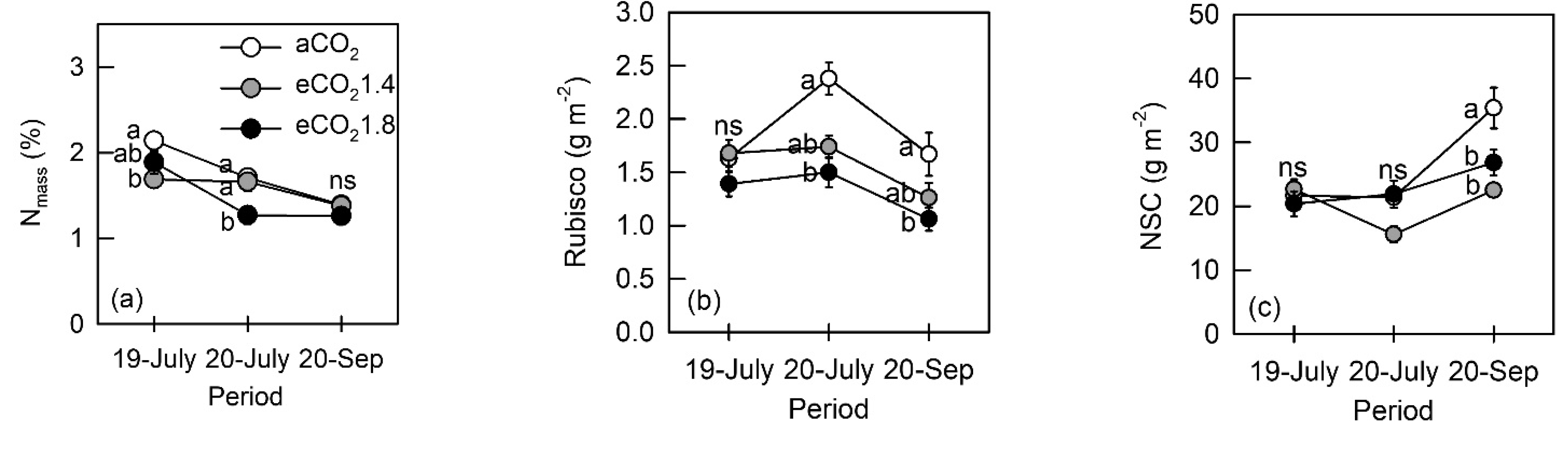
3.2. Photosynthetic Characteristics and Nonstructural Carbohydrates
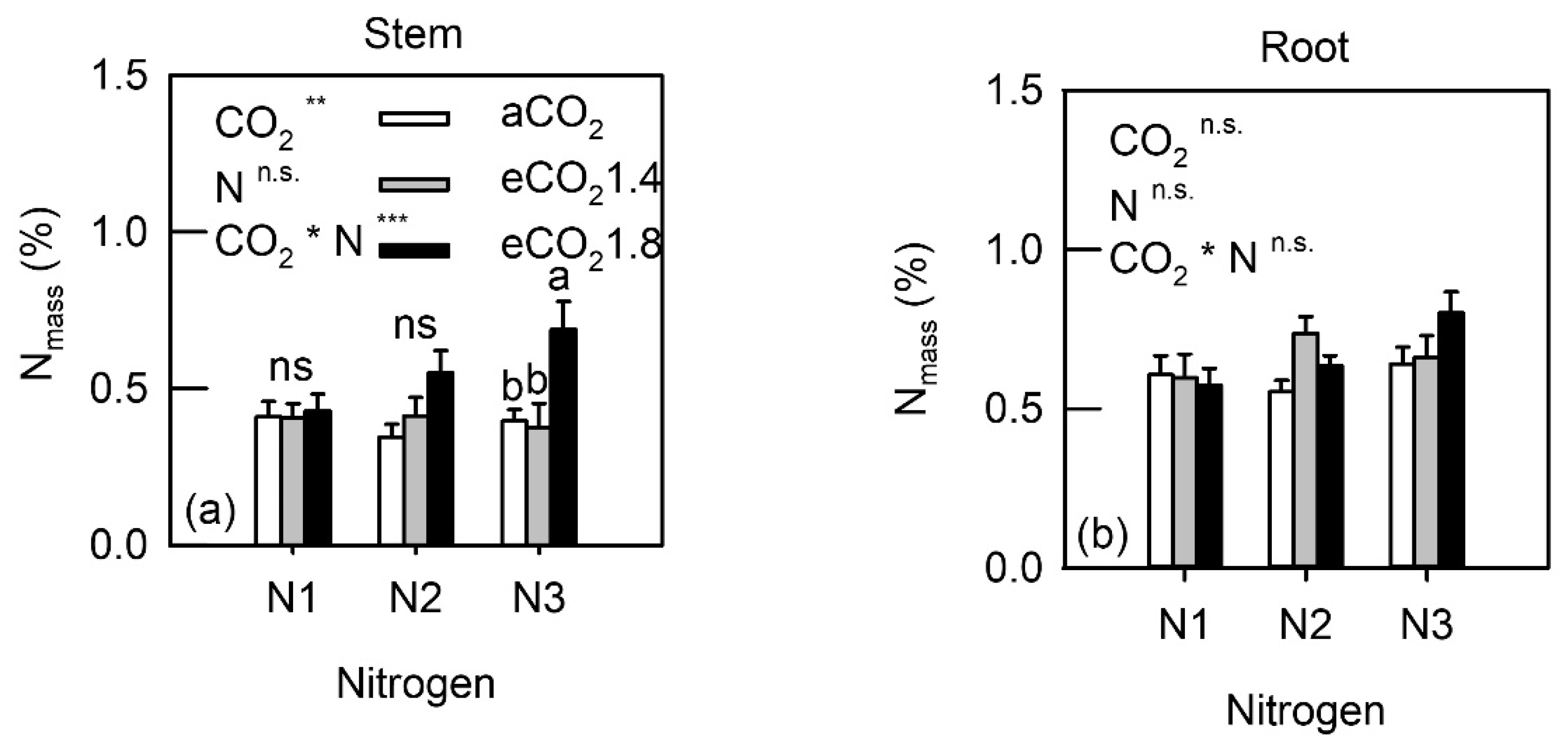
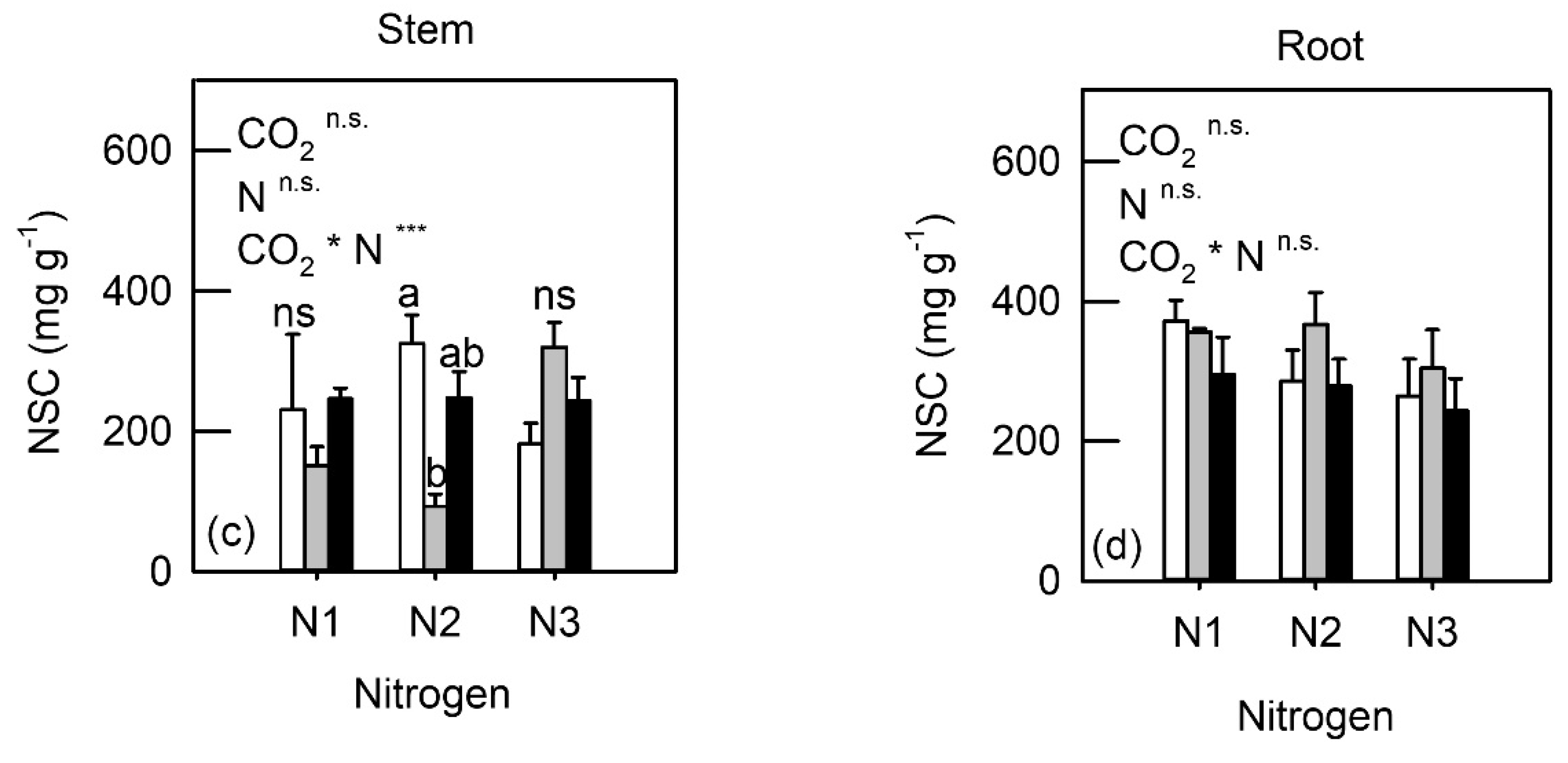
3.3. Whole-Plant Biochemical Characteristics
4. Discussion
4.1. Biomass under eCO2 and N Fertilization
4.2. Down-Regulation of Photosynthesis and NSC under eCO2 and N Fertilization
4.3. PNUE and N Allocation under eCO2 Conditions and N Fertilization
4.4. Biochemical Changes in Sink Organs under eCO2 and N Fertilization
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Drake, B.G.; Gonzalez-Meler, M.A.; Long, S.P. More efficient plants: A consequence of rising atmospheric CO2? Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 609–639. [Google Scholar] [CrossRef] [Green Version]
- Reich, P.B.; Hobbie, S.E.; Lee, T.D.; Pastore, M.A. Unexpected reversal of C3 versus C4 grass response to elevated CO2 during a 20-year field experiment. Science 2018, 360, 317–320. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef]
- Norby, R.J.; Warren, J.M.; Iversen, C.M.; Medlyn, B.E.; McMurtrie, R.E. CO2 enhancement of forest productivity constrained by limited nitrogen availability. Proc. Natl. Acad. Sci. USA 2010, 107, 19368–19373. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Medlyn, B.E.; Drake, J.E.; Duursma, R.A.; Anderson, I.C.; Barton, C.V.; Boer, M.M.; Carrillo, Y.; Castañeda-Gómez, L.; Collins, L. The fate of carbon in a mature forest under carbon dioxide enrichment. Nature 2020, 580, 227–231. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Y.; Ju, W.; Chen, J.M.; Ciais, P.; Cescatti, A.; Sardans, J.; Janssens, I.A.; Wu, M.; Berry, J.A. Recent global decline of CO2 fertilization effects on vegetation photosynthesis. Science 2020, 370, 1295–1300. [Google Scholar] [CrossRef]
- Luo, Y.; Su, B.; Currie, W.S.; Dukes, J.S.; Finzi, A.C.; Hartwig, U.; Hungate, B.; McMurtrie, R.E.; Oren, R.; Parton, W.J.; et al. Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide. Bioscience 2004, 54, 731–739. [Google Scholar] [CrossRef] [Green Version]
- Leakey, A.D.B.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. J. Exp. Bot. 2009, 60, 2859–2876. [Google Scholar] [CrossRef]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of C 3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef]
- Moore, B.; Cheng, S.H.; Sims, D.; Seemann, J. The biochemical and molecular basis for photosynthetic acclimation to elevated atmospheric CO2. Plant Cell Environ. 1999, 22, 567–582. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A.; Nelson, R.; Long, S.P. Testing the “source–sink” hypothesis of down-regulation of photosynthesis in elevated [CO2] in the field with single gene substitutions in Glycine max. Agric. For. Meteorol. 2004, 122, 85–94. [Google Scholar] [CrossRef]
- Hovenden, M.J. Photosynthesis of coppicing poplar clones in a free-air CO2 enrichment (FACE) experiment in a short-rotation forest. Funct. Plant Biol. 2003, 30, 391–400. [Google Scholar] [CrossRef]
- Kelly, A.A.; van Erp, H.; Quettier, A.-L.; Shaw, E.; Menard, G.; Kurup, S.; Eastmond, P.J. The sugar-dependent1 lipase limits triacylglycerol accumulation in vegetative tissues of Arabidopsis. Plant Physiol. 2013, 162, 1282–1289. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, D.; Watanabe, C.K.; Betsuyaku, E.; Terashima, I. Sink–source balance and down-regulation of photosynthesis in Raphanus sativus: Effects of grafting, N and CO2. Plant Cell Physiol. 2017, 58, 2043–2056. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Vera, U.M.; De Souza, A.P.; Long, S.P.; Ort, D.R. The role of sink strength and nitrogen availability in the down-regulation of photosynthetic capacity in field-grown Nicotiana tabacum L. at elevated CO2 concentration. Front. Plant Sci. 2017, 8, 998. [Google Scholar] [CrossRef]
- Kanno, K.; Suzuki, Y.; Makino, A. A small decrease in Rubisco content by individual suppression of RBCS genes leads to improvement of photosynthesis and greater biomass production in rice under conditions of elevated CO2. Plant Cell Physiol. 2017, 58, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Wujeska-Klause, A.; Crous, K.Y.; Ghannoum, O.; Ellsworth, D.S. Lower photorespiration in elevated CO2 reduces leaf N concentrations in mature Eucalyptus trees in the field. Glob. Chang. Biol. 2019, 25, 1282–1295. [Google Scholar] [CrossRef]
- Gifford, R.M.; Barrett, D.J.; Lutze, J.L. The effects of elevated [CO2] on the C: N and C: P mass ratios of plant tissues. Plant Soil 2000, 224, 1–14. [Google Scholar] [CrossRef]
- Taub, D.R.; Wang, X. Why are nitrogen concentrations in plant tissues lower under elevated CO2? A critical examination of the hypotheses. J. Integr. Plant Biol. 2008, 50, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- Taub, D.R.; Miller, B.; Allen, H. Effects of elevated CO2 on the protein concentration of food crops: A meta-analysis. Glob. Chang. Biol. 2008, 14, 565–575. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Schjoerring, J.K. Effects of elevated atmospheric CO2 on physiology and yield of wheat (Triticum aestivum L.): A meta-analytic test of current hypotheses. Agric. Ecosyst. Environ. 2013, 178, 57–63. [Google Scholar] [CrossRef]
- Li, W.; Hartmann, H.; Adams, H.D.; Zhang, H.; Jin, C.; Zhao, C.; Guan, D.; Wang, A.; Yuan, F.; Wu, J. The sweet side of global change–dynamic responses of non-structural carbohydrates to drought, elevated CO2 and nitrogen fertilization in tree species. Tree Physiol. 2018, 38, 1706–1723. [Google Scholar] [CrossRef]
- Yin, X. Responses of leaf nitrogen concentration and specific leaf area to atmospheric CO2 enrichment: A retrospective synthesis across 62 species. Glob. Chang. Biol. 2002, 8, 631–642. [Google Scholar] [CrossRef]
- Bazzaz, F.; Miao, S. Successional Status, Seed Size, and Responses of Tree Seedlings to CO 2, Light, and Nutrients. Ecology 1993, 74, 104–112. [Google Scholar] [CrossRef]
- Sholtis, J.D.; Gunderson, C.A.; Norby, R.J.; Tissue, D.T. Persistent stimulation of photosynthesis by elevated CO2 in a sweetgum (Liquidambar styraciflua) forest stand. New Phytol. 2004, 162, 343–354. [Google Scholar] [CrossRef]
- Ellsworth, D.S.; Reich, P.B.; Naumburg, E.S.; Koch, G.W.; Kubiske, M.E.; Smith, S.D. Photosynthesis, carboxylation and leaf nitrogen responses of 16 species to elevated pCO2 across four free-air CO2 enrichment experiments in forest, grassland and desert. Glob. Chang. Biol. 2004, 10, 2121–2138. [Google Scholar] [CrossRef] [Green Version]
- Kerstiens, G. Meta-analysis of the interaction between shade-tolerance, light environment and growth response of woody species to elevated CO2. Acta Oecol. 2001, 22, 61–69. [Google Scholar] [CrossRef]
- Dirnböck, T.; Kraus, D.; Grote, R.; Klatt, S.; Kobler, J.; Schindlbacher, A.; Seidl, R.; Thom, D.; Kiese, R. Substantial understory contribution to the C sink of a European temperate mountain forest landscape. Landsc. Ecol. 2020, 35, 483–499. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Liu, Z.; Chen, D.; Huang, G.; Zhou, L.; Fu, S. Understory plants can make substantial contributions to soil respiration: Evidence from two subtropical plantations. Soil Biol. Biochem. 2011, 43, 2355–2357. [Google Scholar] [CrossRef]
- Stocker, T.F.; Qin, D.; Plattner, G.-K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change 2013: The Physical Science Basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013; Volume 1535. [Google Scholar]
- Lee, J.-C.; Kim, D.-H.; Kim, G.-N.; Kim, P.-G.; Han, S.-H. Long-term climate change research facility for trees: CO2-enriched open top chamber system. Korean J. Agric. For. Meteorol. 2012, 14, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, H.; Eklundh, L.; Hall, K.; Lindroth, A. Estimating LAI in deciduous forest stands. Agric. For. Meteorol. 2005, 129, 27–37. [Google Scholar] [CrossRef]
- Billings, S.A.; Ziegler, S.E. Altered patterns of soil carbon substrate usage and heterotrophic respiration in a pine forest with elevated CO2 and N fertilization. Glob. Chang. Biol. 2008, 14, 1025–1036. [Google Scholar] [CrossRef]
- Kitao, M.; Löw, M.; Heerdt, C.; Grams, T.E.; Häberle, K.-H.; Matyssek, R. Effects of chronic elevated ozone exposure on gas exchange responses of adult beech trees (Fagus sylvatica) as related to the within-canopy light gradient. Environ. Pollut. 2009, 157, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups; Springer Science & Business Media, Institute of Botany University of Innsbruck Sternwartestrasse: Innsbruck, Austria, 2003. [Google Scholar]
- Sharkey, T.D. What gas exchange data can tell us about photosynthesis. Plant Cell Environ. 2016, 39, 1161–1163. [Google Scholar] [CrossRef] [PubMed]
- Ashwell, G. New colorimetric methods of sugar analysis. In Methods in Enzymology; Elsevier, Michigan State University: East Lansing, MI, USA, 1966; Volume 8, pp. 85–95. [Google Scholar]
- Hikosaka, K.; Shigeno, A. The role of Rubisco and cell walls in the interspecific variation in photosynthetic capacity. Oecologia 2009, 160, 443–451. [Google Scholar] [CrossRef]
- Shinano, T.; Lei, T.; Kawamukai, T.; Inoue, M.; Koike, T.; Tadano, T. Dimethylsulfoxide method for the extraction of chlorophylls a and b from the leaves of wheat, field bean, dwarf bamboo, and oak. Photosynthetica 1996, 32, 409–415. [Google Scholar]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- van der Kooi, C.J.; Reich, M.; Löw, M.; De Kok, L.J.; Tausz, M. Growth and yield stimulation under elevated CO2 and drought: A meta-analysis on crops. Environ. Exp. Bot. 2016, 122, 150–157. [Google Scholar] [CrossRef]
- McCarthy, H.R.; Oren, R.; Johnsen, K.H.; Gallet-Budynek, A.; Pritchard, S.G.; Cook, C.W.; LaDeau, S.L.; Jackson, R.B.; Finzi, A.C. Re-assessment of plant carbon dynamics at the Duke free-air CO2 enrichment site: Interactions of atmospheric [CO2] with nitrogen and water availability over stand development. New Phytol. 2010, 185, 514–528. [Google Scholar] [CrossRef]
- Ryan, M.G. Three decades of research at Flakaliden advancing whole-tree physiology, forest ecosystem and global change research. Tree Physiol. 2013, 33, 1123–1131. [Google Scholar] [CrossRef] [Green Version]
- Souza, L.; Belote, R.T.; Kardol, P.; Weltzin, J.F.; Norby, R.J. CO2 enrichment accelerates successional development of an understory plant community. J. Plant Ecol. 2010, 3, 33–39. [Google Scholar] [CrossRef]
- Körner, C.; Arnone, J.A. Responses to elevated carbon dioxide in artificial tropical ecosystems. Science 1992, 257, 1672–1675. [Google Scholar] [CrossRef] [PubMed]
- Hättenschwiler, S.e.; Körner, C.r. Tree seedling responses to in situ CO2-enrichment differ among species and depend on understorey light availability. Glob. Chang. Biol. 2000, 6, 213–226. [Google Scholar] [CrossRef]
- Bloor, J.; Barthes, L.; Leadley, P.W. Effects of elevated CO2 and N on tree–grass interactions: An experimental test using Fraxinus excelsior and Dactylis glomerata. Funct. Ecol. 2008, 22, 537–546. [Google Scholar] [CrossRef]
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Tian, D.; Li, P.; Fang, W.; Xu, J.; Luo, Y.; Yan, Z.; Zhu, B.; Wang, J.; Xu, X.; Fang, J. Growth responses of trees and understory plants to nitrogen fertilization in a subtropical forest in China. Biogeosciences 2017, 14, 3461–3469. [Google Scholar] [CrossRef] [Green Version]
- Son, Y.; Lee, M.-H.; Noh, N.J.; Kang, B.H.; Kim, K.O.; Yi, M.J.; Byun, J.K.; Yi, K. Fertilization effects on understory vegetation biomass and structure in four different plantations. J. Korean Soc. For. Sci. 2007, 96, 520–527. [Google Scholar]
- Wang, F.; Chen, F.; Wang, G.G.; Mao, R.; Fang, X.; Wang, H.; Bu, W. Effects of experimental nitrogen addition on nutrients and nonstructural carbohydrates of dominant understory plants in a Chinese fir plantation. Forests 2019, 10, 155. [Google Scholar] [CrossRef] [Green Version]
- Rogers, A.; Ellsworth, D.S. Photosynthetic acclimation of Pinus taeda (loblolly pine) to long-term growth in elevated pCO(2) (FACE). Plant Cell Environ. 2002, 25, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Stitt, M.; Krapp, A. The interaction between elevated carbon dioxide and nitrogen nutrition: The physiological and molecular background. Plant Cell Environ. 1999, 22, 583–621. [Google Scholar] [CrossRef]
- Zotz, G.; Cueni, N.; Körner, C. In situ growth stimulation of a temperate zone liana (Hedera helix) in elevated CO2. Funct. Ecol. 2006, 20, 763–769. [Google Scholar] [CrossRef]
- Ziska, L.H.; Bunce, J.A. Sensitivity of field-grown soybean to future atmospheric CO2: Selection for improved productivity in the 21st century. Funct. Plant Biol. 2000, 27, 979–984. [Google Scholar] [CrossRef]
- Ziska, L.H.; Bunce, J.A.; Caulfield, F.A. Rising atmospheric carbon dioxide and seed yield of soybean genotypes. Crop Sci. 2001, 41, 385–391. [Google Scholar] [CrossRef]
- Bloom, A.J.; Burger, M.; Asensio, J.S.R.; Cousins, A.B. Carbon dioxide enrichment inhibits nitrate assimilation in wheat and Arabidopsis. Science 2010, 328, 899–903. [Google Scholar] [CrossRef] [Green Version]
- Sefcik, L.T.; Zak, D.R.; Ellsworth, D.S. Photosynthetic responses to understory shade and elevated carbon dioxide concentration in four northern hardwood tree species. Tree Physiol. 2006, 26, 1589–1599. [Google Scholar] [CrossRef] [Green Version]
- Hättenschwiler, S. Tree seedling growth in natural deep shade: Functional traits related to interspecific variation in response to elevated CO2. Oecologia 2001, 129, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Leakey, A.; Press, M.C.; Scholes, J.; Watling, J. Relative enhancement of photosynthesis and growth at elevated CO2 is greater under sunflecks than uniform irradiance in a tropical rain forest tree seedling. Plant Cell Environ. 2002, 25, 1701–1714. [Google Scholar] [CrossRef]
- Granados, J.; Körner, C. In deep shade, elevated CO2 increases the vigor of tropical climbing plants. Glob. Chang. Biol. 2002, 8, 1109–1117. [Google Scholar] [CrossRef]
- Novriyanti, E.; Watanabe, M.; Kitao, M.; Utsugi, H.; Uemura, A.; Koike, T. High nitrogen and elevated [CO2] effects on the growth, defense and photosynthetic performance of two eucalypt species. Environ. Pollut. 2012, 170, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Davey, P.; Parsons, A.; Atkinson, L.; Wadge, K.; Long, S.P. Does photosynthetic acclimation to elevated CO2 increase photosynthetic nitrogen-use efficiency? A study of three native UK grassland species in open-top chambers. Funct. Ecol. 1999, 13, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.R.; Seemann, J.R. The allocation of protein nitrogen in the photosynthetic apparatus: Costs, consequences, and control. Photosynthesis 1989, 8, 183–205. [Google Scholar]
- Choi, D.; Watanabe, Y.; Guy, R.D.; Sugai, T.; Toda, H.; Koike, T. Photosynthetic characteristics and nitrogen allocation in the black locust (Robinia pseudoacacia L.) grown in a FACE system. Acta Physiol. Plant. 2017, 39. [Google Scholar] [CrossRef]
- Sallas, L.; Luomala, E.-M.; Utriainen, J.; Kainulainen, P.; Holopainen, J.K. Contrasting effects of elevated carbon dioxide concentration and temperature on Rubisco activity, chlorophyll fluorescence, needle ultrastructure and secondary metabolites in conifer seedlings. Tree Physiol. 2003, 23, 97–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Mao, Z.; Xu, J. Gas exchange, chlorophyll and growth responses of Betula Platyphylla seedlings to elevated CO2 and nitrogen. Int. J. Biol. 2010, 2, 143. [Google Scholar] [CrossRef]
- Nie, M.; Lu, M.; Bell, J.; Raut, S.; Pendall, E. Altered root traits due to elevated CO2: A meta-analysis. Glob. Ecol. Biogeogr. 2013, 22, 1095–1105. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Ineson, P.; Scott, A. Elevated CO2 reduces the nitrogen concentration of plant tissues. Glob. Change Biol. 1998, 4, 43–54. [Google Scholar] [CrossRef]
- Kim, H.Y.; Lieffering, M.; Kobayashi, K.; Okada, M.; Miura, S. Seasonal changes in the effects of elevated CO2 on rice at three levels of nitrogen supply: A free air CO2 enrichment (FACE) experiment. Glob. Change Biol. 2003, 9, 826–837. [Google Scholar] [CrossRef]
- Zhao, H.; Xu, X.; Zhang, Y.; Korpelainen, H.; Li, C. Nitrogen deposition limits photosynthetic response to elevated CO2 differentially in a dioecious species. Oecologia 2011, 165, 41–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | df | Amax | VCmax | Jmax | Nmass | Narea | Rubisco | NSC | PNUE | Chlorophyll | Chlorophyll: Rubisco |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CO2 | 2 | 0.020 | 0.083 | 0.007 | <0.001 | <0.001 | <0.001 | <0.001 | 0.021 | <0.001 | 0.708 |
| N | 2 | 0.297 | 0.927 | 0.384 | 0.021 | 0.870 | 0.783 | 0.256 | 0.442 | 0.485 | 0.813 |
| Period | 2 | 0.167 | 0.001 | 0.002 | <0.001 | <0.001 | 0.002 | <0.001 | 0.219 | <0.001 | 0.588 |
| CO2 × N | 4 | 0.348 | 0.249 | 0.199 | 0.496 | 0.913 | 0.321 | 0.140 | 0.631 | 0.925 | 0.157 |
| CO2 × Period | 4 | 0.996 | 0.257 | 0.219 | <0.001 | 0.117 | 0.029 | 0.010 | 0.063 | 0.611 | 0.549 |
| N × Period | 4 | 0.360 | 0.117 | 0.010 | 0.872 | 0.542 | 0.710 | 0.693 | 0.192 | 0.031 | 0.944 |
| Nitrogen | Chamber | Amax (μmol m−2s−1) | VCmax (μmol m−2s−1) | Jmax (μmol m−2s−1) | Nmass (%) | Narea (g m−2) | Rubisco (g m−2) | NSC (g m−2) | PNUE (μmol (molN)−1s−1) | Chlorophyll (g m−2) |
|---|---|---|---|---|---|---|---|---|---|---|
| N1 | aCO2 | 5.57 ± 0.53 ns | 35.03 ± 5.55 ns | 58.20 ± 5.08 ns | 1.61 ± 0.11 ns | 0.99 ± 0.10 a | 1.56 ± 0.12 ns | 23.68 ± 1.58 ns | 78.8 ± 4.4 b | 0.15 ± 0.01 ns |
| eCO21.4 | 6.76 ± 0.70 | 42.41 ± 8.68 | 63.81 ± 8.89 | 1.38 ± 0.11 | 0.73 ± 0.07 ab | 1.41 ± 0.13 | 18.59 ± 1.28 | 133.9 ± 12.1 a | 0.13 ± 0.02 | |
| eCO21.8 | 7.43 ± 0.91 | 29.03 ± 6.52 | 51.10 ± 6.66 | 1.20 ± 0.17 | 0.65 ± 0.08 b | 1.18 ± 0.12 | 24.69 ± 2.56 | 142.0 ± 17.8 a | 0.12 ± 0.02 | |
| N2 | aCO2 | 6.77 ± 0.52 ns | 38.45 ± 7.00 ns | 65.66 ± 4.82 ns | 1.85 ± 0.14 ns | 1.01 ± 0.09 a | 1.72 ± 0.15 ns | 24.77 ± 2.40 ns | 95.52 ± 9.2 b | 0.17 ± 0.01 ns |
| eCO21.4 | 8.52 ± 0.80 | 37.24 ± 4.67 | 70.31 ± 6.29 | 1.76 ± 0.07 | 1.00 ± 0.09 ab | 1.63 ± 0.11 | 20.35 ± 1.33 | 130.6 ± 13.6 ab | 0.18 ± 0.01 | |
| eCO21.8 | 9.19 ± 0.85 | 35.78 ± 4.19 | 59.21 ± 5.50 | 1.55 ± 0.08 | 0.80 ± 0.05 b | 1.37 ± 0.14 | 21.34 ± 1.48 | 158.8 ± 14.0 a | 0.15 ± 0.01 | |
| N3 | aCO2 | 7.97 ± 1.00 ns | 42.90 ± 5.87 ns | 75.07 ± 6.45 a | 1.75 ± 0.15 ns | 1.17 ± 0.08 ns | 2.27 ± 0.20 a | 30.67 ± 3.89 a | 97.0 ± 11.2 b | 0.18 ± 0.01 ns |
| eCO21.4 | 9.62 ± 0.75 | 50.84 ± 5.08 | 72.14 ± 5.52 ab | 1.65 ± 0.08 | 0.96 ± 0.13 | 1.65 ± 0.14 ab | 22.63 ± 1.89 b | 155.3 ± 22.1 ab | 0.16 ± 0.02 | |
| eCO21.8 | 10.10 ± 1.04 | 23.72 ± 4.45 | 50.73 ± 10.39 b | 1.74 ± 0.15 | 0.89 ± 0.04 | 1.38 ± 0.11 b | 22.97 ± 2.18 b | 154.5 ± 16.2 a | 0.18 ± 0.01 | |
| Average | aCO2 | 6.80 ± 0.44 b | 38.46 ± 3.47 ns | 65.61 ± 3.30 a | 1.74 ± 0.08 a | 1.05 ± 0.05 a | 1.87 ± 0.11 a | 26.26 ± 1.61 a | 90.9 ± 3.4 b | 0.17 ± 0.01 a |
| eCO21.4 | 8.17 ± 0.47 a | 42.69 ± 4.20 | 67.96 ± 4.43 a | 1.58 ± 0.06 b | 0.89 ± 0.06 b | 1.57 ± 0.08 b | 20.48 ± 0.89 b | 138.2 ± 8.7 a | 0.16 ± 0.01 ab | |
| eCO21.8 | 8.91 ± 0.55 a | 29.73 ± 3.05 | 53.79 ± 4.26 b | 1.49 ± 0.08 b | 0.78 ± 0.04 b | 1.31 ± 0.08 c | 23.04 ± 1.23 ab | 151.4 ± 9.2 a | 0.15 ± 0.01 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byeon, S.; Kim, K.; Hong, J.; Kim, S.; Kim, S.; Park, C.; Ryu, D.; Han, S.-H.; Oh, C.; Kim, H.S. Down-Regulation of Photosynthesis to Elevated CO2 and N Fertilization in Understory Fraxinus rhynchophylla Seedlings. Forests 2021, 12, 1197. https://doi.org/10.3390/f12091197
Byeon S, Kim K, Hong J, Kim S, Kim S, Park C, Ryu D, Han S-H, Oh C, Kim HS. Down-Regulation of Photosynthesis to Elevated CO2 and N Fertilization in Understory Fraxinus rhynchophylla Seedlings. Forests. 2021; 12(9):1197. https://doi.org/10.3390/f12091197
Chicago/Turabian StyleByeon, Siyeon, Kunhyo Kim, Jeonghyun Hong, Seohyun Kim, Sukyung Kim, Chanoh Park, Daun Ryu, Sim-Hee Han, Changyoung Oh, and Hyun Seok Kim. 2021. "Down-Regulation of Photosynthesis to Elevated CO2 and N Fertilization in Understory Fraxinus rhynchophylla Seedlings" Forests 12, no. 9: 1197. https://doi.org/10.3390/f12091197
APA StyleByeon, S., Kim, K., Hong, J., Kim, S., Kim, S., Park, C., Ryu, D., Han, S.-H., Oh, C., & Kim, H. S. (2021). Down-Regulation of Photosynthesis to Elevated CO2 and N Fertilization in Understory Fraxinus rhynchophylla Seedlings. Forests, 12(9), 1197. https://doi.org/10.3390/f12091197