
A Stand-Level Comparison of Carbon and Nitrogen Distribution in an Exotic Japanese Cedar Plantation and a Natural Oak Stand




Abstract
:1. Introduction
2. Materials and Methods
2.1. Stand Characteristics
2.2. Carbon and Nitrogen Stocks in the Aboveground Tree Biomass
2.3. Carbon and Nitrogen Stocks in the Forest Floor and Mineral Soils
2.4. Data Analysis
3. Results
3.1. Stand and Soil Characteristics
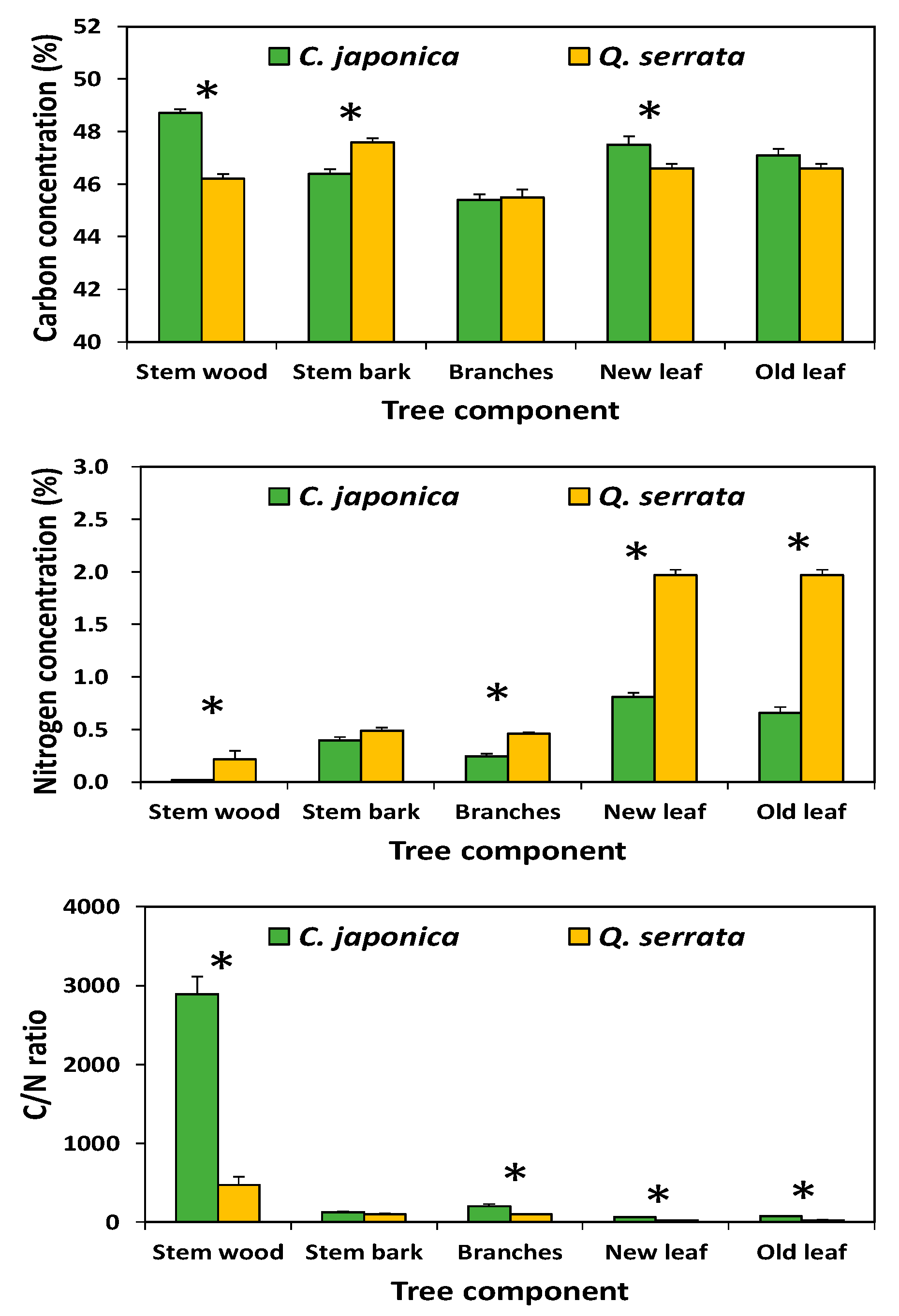
3.2. Carbon and Nitrogen Concentration in Aboveground Tree Biomass and the Forest Floor
3.3. Allometric Equation for Estimation of Carbon and Nitrogen Content
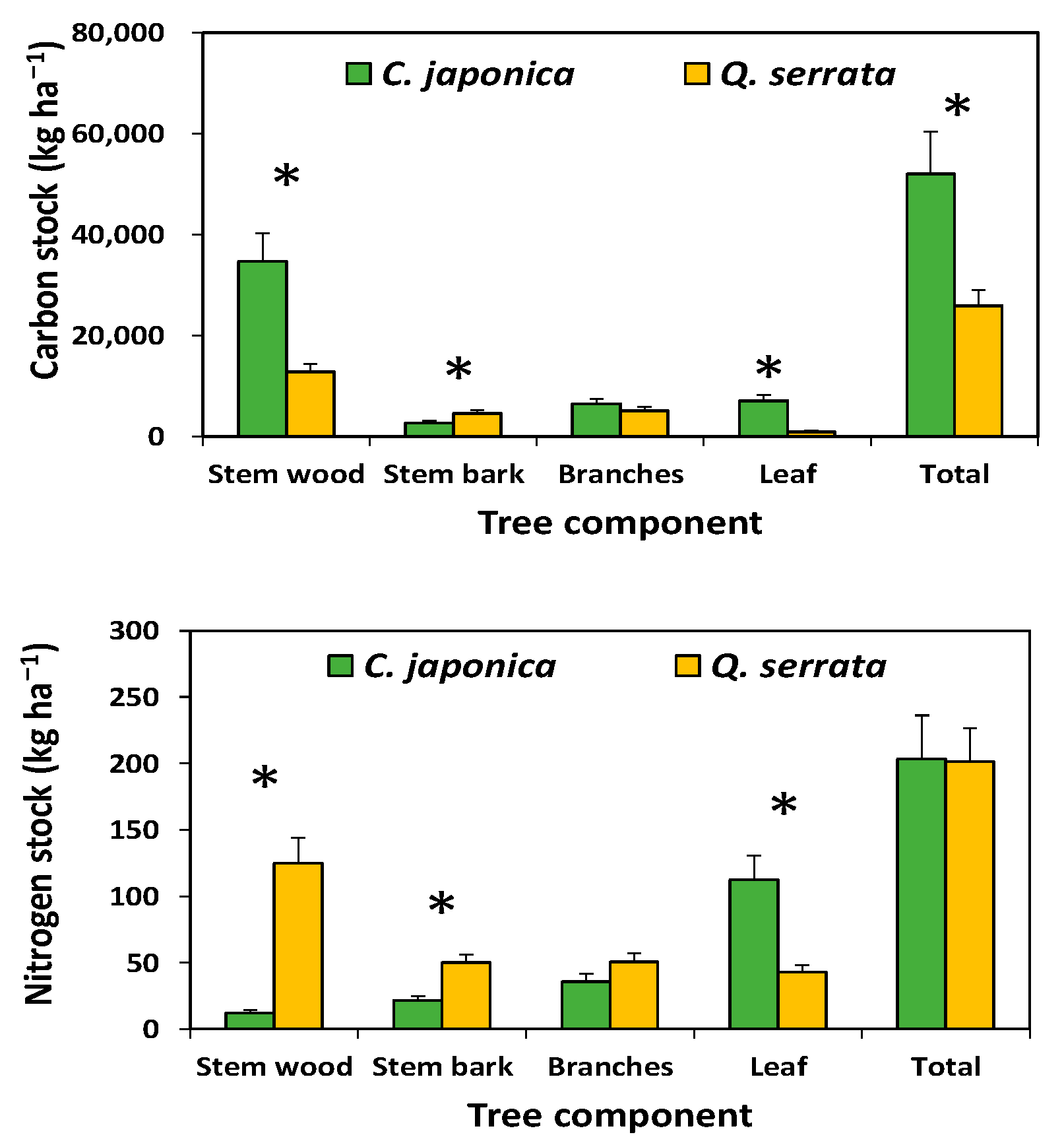
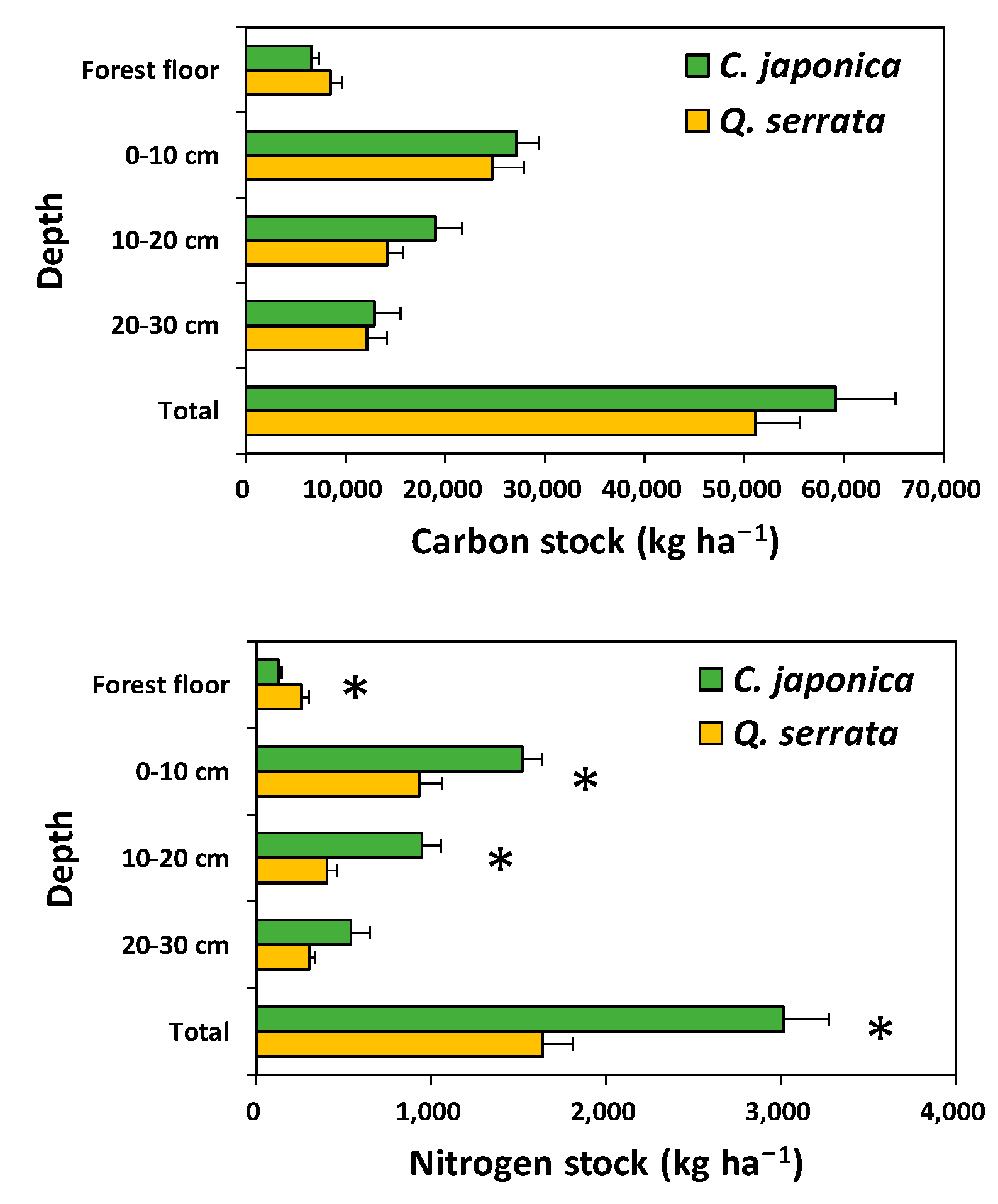
3.4. Carbon and Nitrogen Stocks at the Stand Level
3.5. Relationship between Soil C or N Stocks and Soil Property
4. Discussion
4.1. Carbon and Nitrogen Status of the Aboveground Tree Biomass, the Forest Floor, and the Mineral Soils
4.2. Carbon and Nitrogen Stocks in Aboveground Tree Biomass, the Forest Floor, and the Mineral Soils
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | Tree Species | Component (y) | Regression Coefficient | R2 | RMSE | p-Value | CF | |
|---|---|---|---|---|---|---|---|---|
| a | b | |||||||
| Carbon | C. japonica | Stem wood (kg) | −0.9340 | 1.9320 | 0.7593 | 0.1169 | 0.0010 | 1.016 |
| Stem bark (kg) | −1.6232 | 1.6217 | 0.5982 | 0.1428 | 0.0087 | 1.024 | ||
| Branch (kg) | −2.1295 | 2.2440 | 0.8159 | 0.1145 | 0.0003 | 1.015 | ||
| Old leaf (kg) | −1.4949 | 1.7770 | 0.7978 | 0.0961 | 0.0005 | 1.031 | ||
| New leaf (kg) | −1.7683 | 1.5590 | 0.5164 | 0.1621 | 0.0192 | 1.031 | ||
| Total (kg) | −0.7838 | 1.9496 | 0.8420 | 0.0908 | 0.0002 | 1.010 | ||
| Q. serrata | Stem wood (kg) | −1.1565 | 2.0252 | 0.9810 | 0.0411 | 0.0011 | 1.002 | |
| Stem bark (kg) | −1.2255 | 1.6996 | 0.9568 | 0.0659 | <0.0001 | 1.005 | ||
| Branch (kg) | −3.0163 | 3.2595 | 0.9305 | 0.1624 | <0.0001 | 1.031 | ||
| Leaf (kg) | −3.6166 | 3.1693 | 0.8830 | 0.1961 | 0.0002 | 1.045 | ||
| Total (kg) | −1.0704 | 2.2131 | 0.9507 | 0.0919 | <0.0001 | 1.010 | ||
| Nitrogen | C. japonica | Stem wood (kg) | −4.3213 | 1.8869 | 0.5380 | 0.1879 | 0.0158 | 1.041 |
| Stem bark (kg) | −3.9832 | 1.8200 | 0.6603 | 0.1403 | 0.0043 | 1.023 | ||
| Branch (kg) | −4.8970 | 2.5896 | 0.5762 | 0.2387 | 0.0109 | 1.068 | ||
| Old leaf (kg) | −4.1905 | 2.3732 | 0.8221 | 0.1186 | 0.0003 | 1.016 | ||
| New leaf (kg) | −4.1620 | 2.0023 | 0.5865 | 0.1806 | 0.0098 | 1.038 | ||
| Total (kg) | −3.8238 | 2.3763 | 0.8878 | 0.0908 | <0.0001 | 1.010 | ||
| Q. serrata | Stem wood (kg) | −5.4669 | 3.9385 | 0.5893 | 0.4793 | 0.0451 | 1.203 | |
| Stem bark (kg) | −3.6655 | 2.1109 | 0.9428 | 0.0948 | <0.0001 | 1.010 | ||
| Branch (kg) | −4.9095 | 3.1625 | 0.9215 | 0.1683 | <0.0001 | 1.437 | ||
| Leaf (kg) | −4.9525 | 3.1384 | 0.8783 | 0.1986 | 0.0002 | 1.046 | ||
| Total (kg) | −3.6436 | 2.6088 | 0.9023 | 0.1565 | <0.0001 | 1.029 | ||
References
- Chen, X.; Li, B.-L. Change in soil carbon and nutrient storage after human disturbance of a primary Korean pine forest in northeast China. For. Ecol. Manag. 2003, 186, 197–206. [Google Scholar] [CrossRef]
- Li, D.; Niu, S.; Luo, Y. Global patterns of the dynamics of soil carbon and nitrogen stocks following afforestation: A meta-analysis. New Phytol. 2012, 195, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Han, S.H.; Meng, L.; Park, G.S.; Kim, S.B.; Cho, M.S.; Park, B.B. Characteristics of soil carbon and nutrient stocks across land use types in a forest region of central Korea. For. Sci. Technol. 2017, 13, 93–99. [Google Scholar] [CrossRef]
- Kim, C.; Yoo, B.O.; Jung, S.Y.; Lee, K.S. Allometric equations to assess biomass, carbon and nitrogen content of black pine and red pine trees in southern Korea. Forest 2017, 10, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.J.; Yim, J.S.; Son, Y.M.; Son, Y.; Kim, R. Estimation of forest carbon stocks for National greenhouse gas inventory reporting in South Korea. Forests 2018, 9, 625. [Google Scholar] [CrossRef] [Green Version]
- Binkley, D.; Giardina, C. Why do tree species affect soils? The warp and woof of tree-soil interactions. Biogeochemistry 1998, 42, 89–106. [Google Scholar] [CrossRef]
- Augusto, L.; Ranger, J.; Binkley, D.; Rothe, A. Impact of several common tree species of European temperate forests on soil fertility. Ann. For. Sci. 2002, 59, 233–253. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Xu, W.; Zhou, G.; Bai, Y.; Li, J.; Tang, X.; Chen, D.; Liu, Q.; Ma, W.; Xiong, G.; et al. Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 2018, 115, 4033–4038. [Google Scholar] [CrossRef] [Green Version]
- Omoro, L.M.A.; Starr, M.; Pellikka, P.K.E. Tree biomass and soil carbon stocks in indigenous forests in comparison to plantations of exotic species in the Taita Hills of Kenya. Silva Fenn. 2013, 47, 935. [Google Scholar] [CrossRef] [Green Version]
- Dawud, S.M.; Vesterdal, L.; Raulund-Rasmussen, K. Mixed-species effects on soil C and N stocks, C/N ratio and pH using a transboundary approach in adjacent common garden Douglas-fir and beech stands. Forests 2017, 8, 95. [Google Scholar] [CrossRef] [Green Version]
- Liao, C.; Luo, Y.; Fang, C.; Li, B. Ecosystem carbon stock influenced by plantation practice: Implications for planting forests as a measure of climate change mitigation. PLoS ONE 2010, 5, e10867. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Baek, G.; Choi, B.; Ha, J.; Bae, E.J.; Lee, K.S.; Son, Y.M. Carbon stocks of tree, forest floor, and mineral soil in Cryptomeria japonica and Chamaecyparis obtusa stands. J. Korean Soc. For. Sci. 2020, 109, 169–178. [Google Scholar]
- Paré, D.; Bernier, P.; Lafleur, B.; Titus, B.D.; Thiffault, E.; Maynard, D.G.; Guo, X. Estimating stand-scale biomass, nutrient contents, and associated uncertainties for tree species of Canadian forests. Can. J. For. Res. 2013, 43, 599–608. [Google Scholar] [CrossRef]
- Yang, K.; Zhu, J.; Xu, S.; Zheng, X. Conversion from temperate secondary forests into plantations (Larix spp.): Impact on belowground carbon and nutrient pools in northeastern China. Land Degrad. Dev. 2018, 29, 4129–4139. [Google Scholar] [CrossRef]
- Government of South Korea. Carbon Neutrality Strategies by 2050; Government of South Korea: Seoul, Korea, 2021; p. 117.
- Kim, C.; Lim, J.H.; Lee, I.K.; Park, B.B.; Chun, J.H. Annual variations of litterfall production in a broadleaved deciduous forest at the Mt. Keumsan LTER site. J. Korean For. Soc. 2013, 102, 210–215. [Google Scholar]
- Korea Forest Research Institute. Survey Manual for Biomass and Soil Carbon; Korea Forest Research Institute: Seoul, Korea, 2010; p. 60. [Google Scholar]
- Martin, A.R.; Erickson, D.L.; Kress, W.J.; Thomas, S.C. Wood nitrogen concentrations in tropical trees: Phylogenetic patterns and ecological correlates. New phytol. 2014, 204, 484–495. [Google Scholar] [CrossRef]
- Balboa-Murias, M.A.; Rojo, A.; Álvarez, J.G.; Merino, A. Carbon and nutrient stocks in mature Quercus robur L. stands in NW Spain. Ann. For. Sci. 2006, 63, 557–565. [Google Scholar] [CrossRef] [Green Version]
- Kalra, Y.P.; Maynard, D.G. Methods Manual for Forest Soil and Plant Analysis; Forestry Canada, Northwest Region, Information Report NOR-X-319E; Northern Forestry Centre: Edmonton, AB, Canada, 1991; p. 116. [Google Scholar]
- Garcia Villacorta, A.M.; Martin, T.A.; Jokela, E.J.; Cropper Jr, W.P.; Gezan, S.A. Variation in biomass distribution and nutrient content in loblolly pine (Pinus taeda L.) clones having contrasting crown architecture and growth efficiency. For. Ecol. Manag. 2015, 342, 84–92. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT Statistical Software, version 9.1; SAS publishing: Cary, NC, USA, 2003. [Google Scholar]
- ter Braak, C.J.F.; Śmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, version 5.10; Microcomputer Power: Ithaca, NY, USA, 2018; p. 536. [Google Scholar]
- Kim, C. Carbon and nitrogen distribution of tree components in Larix kaempferi Carriere and Quercus variabilis Blume stands in Gyeongnam province. J. Korean Soc. For. Sci. 2019, 108, 139–146. [Google Scholar]
- Lin, J.-C.; Chiu, C.M.; Lin, Y.-J.; Liu, W.-Y. Thinning effects on biomass and carbon stock for young Taiwania plantations. Sci. Rep. 2018, 8, 3070. [Google Scholar] [CrossRef]
- Kim, C.; Son, Y.; Lee, W.K.; Jeong, J.; Noh, N.J. Influences of forest tending works on carbon distribution and cycling in a Pinus densiflora S. et Z. stand in Korea. For. Ecol. Manag. 2009, 257, 1420–1426. [Google Scholar] [CrossRef]
- Martin, A.R.; Gezahegn, S.; Thomas, S.C. Variation in carbon and nitrogen concentration among major woody tissue types in temperate trees. Can. J. For. Res. 2015, 45, 744–757. [Google Scholar] [CrossRef]
- Bert, D.; Danjon, F. Carbon concentration variations in the roots, stem and crown of mature Pinus pinaster (Ait.). For. Ecol. Manag. 2006, 222, 279–295. [Google Scholar] [CrossRef]
- Chong, S.H.; Park, B.S. Wood Properties of the Useful Tree Species Grown in Korea; New Research Book No. 29; Korea Forest Research Institute: Seoul, Korea, 2008; p. 390. [Google Scholar]
- Meerts, P. Mineral nutrient concentrations in sapwood and heartwood: A literature review. Ann. For. Sci. 2002, 59, 713–722. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.-Y.; Cha, Y.; Oh, N.-H. The effects of tree species on soil organic carbon content in South Korea. JGR Biogeosci. 2019, 124, 708–716. [Google Scholar] [CrossRef]
- Noguchi, K.; Konopka, B.; Satomura, T.; Kaneko, S.; Takahashi, M. Biomass and production of fine roots in Japanese forests. J. For. Res. 2007, 12, 83–95. [Google Scholar] [CrossRef]
- Van Do, T.; Sato, T.; Kozen, O. A new approach for estimating fine root production in forests: A combination of ingrowth core and scanner. Trees 2016, 30, 545–554. [Google Scholar] [CrossRef]
- Noguchi, K.; Nagakura, J.; Kaneke, S. Biomass and morphology of fine roots of sugi (Cryptomeria japonica) after 3 years of nitrogen fertilization. Front. Plant Sci. 2013, 4, 347. [Google Scholar] [CrossRef] [Green Version]
- Han, S.H.; Kim, S.; Cha, H.; Kim, H.-J.; Khamzina, A.; Son, Y. Soil depth-and root diameter-related variations affect root decomposition in temperate pine and oak forests. J. Plant. Ecol. 2019, 12, 871–881. [Google Scholar] [CrossRef]
- Flores, O.; Coomes, D.A. Estimating the wood density of species for carbon stock assessments. Method Ecol. Evol. 2011, 2, 214–220. [Google Scholar] [CrossRef]
- Kim, C. Biomass and nutrient concentrations of fine roots in a Korean pine plantation and a sawtooth oak stand. For. Sci. Technol. 2012, 8, 187–191. [Google Scholar] [CrossRef]
- Li, S.; Su, J.; Liu, W.; Lang, X.; Huang, X.; Jia, C.; Zhang, Z.; Tong, Q. Changes in biomass carbon and soil organic carbon stocks following the conversion from a secondary coniferous forest to a pine plantation. PLoS ONE 2015, 10, e0135946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
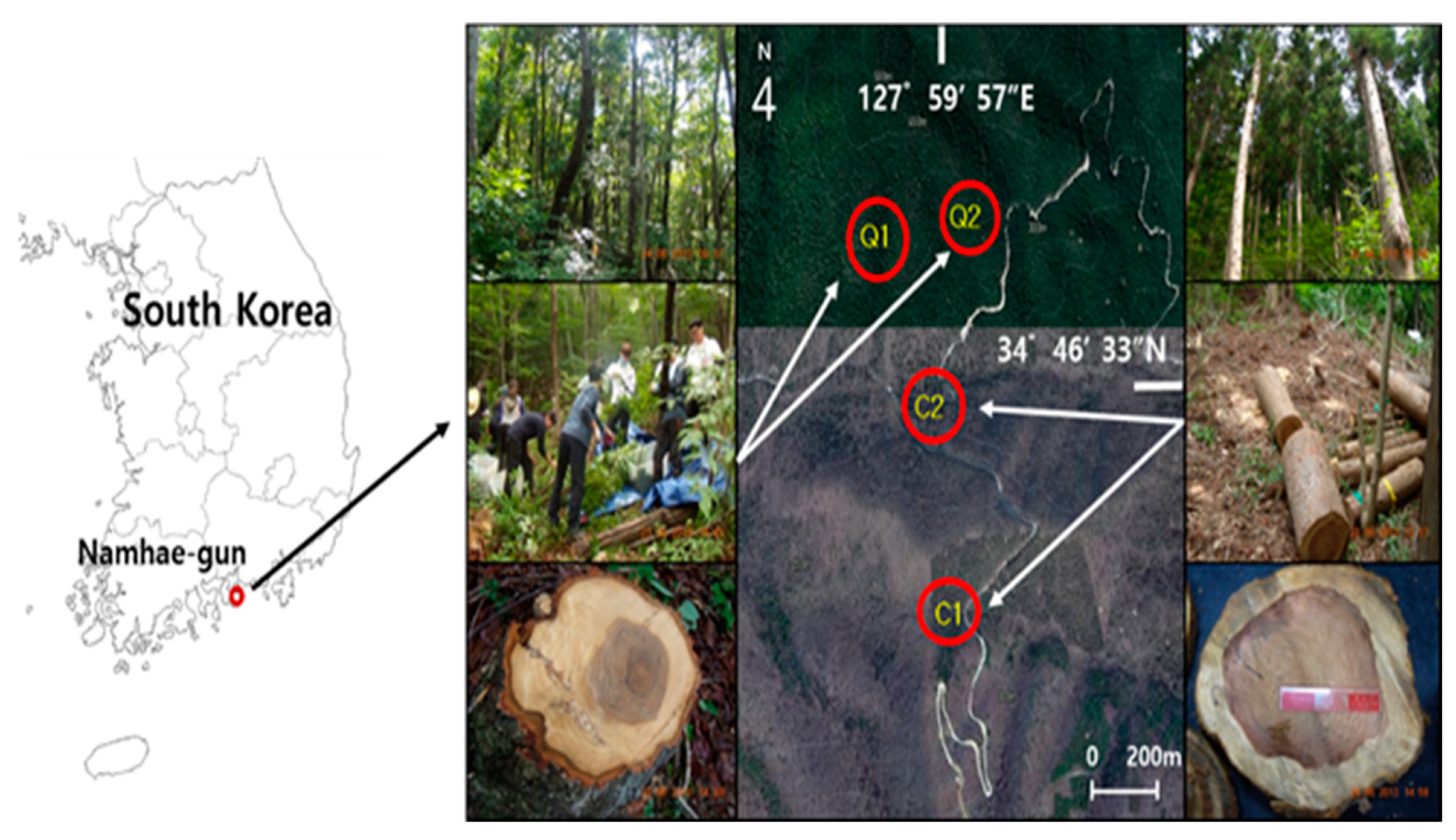

| Stand | Plot | Age | Location | Aspect | Elevation (m) | Slope (°) | Parent Rocks | Stand Density (Tree ha–1) | DBH (cm) | Height (m) | Basal Area (m2 ha–1) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| C. japonica | 1 | 49 | 34°46′21′′ N 128°03′02′′ E | SE | 355 | 30 | Granite | 667 (44) | 25.4 (1.8) | 16.2 (1.5) | 35.54 (7.7) |
| 2 | 47 | 34°46′33′′ N 127°59′59′′ E | NE | 298 | 15 | Granite | 267 (17) | 32.3 (2.6) | 13.3 (1.6) | 23.37 (3.8) | |
| Mean | 48 | 467 (92) | 28.9 (2.1) | 14.8 (1.6) | 29.5 (4.7) | ||||||
| Q. serrata | 1 | 37 | 34°46′36′′ N 127°59′58′′ E | SW | 330 | 20 | Granite | 816 (142) | 12.2 (0.7) | 9.8 (1.5) | 10.0 (1.8) |
| 2 | 36 | 34°46′41′′ N 128°07′97′′ E | S | 334 | 25 | Granite | 1316 (101) | 11.9 (0.6) | 8.2 (0.7) | 16.3 (1.5) | |
| Mean | 37 | 1067 (136) | 12.0 (0.4) | 9.0 (1.3) | 13.5 (1.6) |
| Depth (cm) | Stand | Bulk Density (g cm–3) | Coarse Fragment (g g–1) | Organic C (%) | Total N (%) | C/N Ratio | Extractable P (mg kg–1) | Exchangeable (cmolc kg–1) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| K+ | Ca2+ | Mg2+ | ||||||||
| 0–10 | C. japonica | 0.88 (0.06) | 0.47 (0.02) | 6.1 (0.74) | 0.34 (0.04) | 17.8 (0.7) | 20.7 (3.2) | 0.37 (0.03) | 3.45 (0.61) | 0.51 (0.05) |
| Q. serrata | 1.04 (0.04) | 0.51 (0.03) | 5.0 (0.76) | 0.19 (0.03) | 27.5 (1.9) | 17.3 (0.7) | 0.21 (0.03) | 0.79 (0.42) | 0.22 (0.07) | |
| p-value | 0.04 | 0.27 | 0.32 | 0.01 | 0.003 | 0.33 | 0.005 | 0.005 | 0.008 | |
| 10–20 | C. japonica | 1.03 (0.04) | 0.47 (0.02) | 3.5 (0.49) | 0.17 (0.02) | 19.9 (0.7) | 19.8 (0.7) | 0.22 (0.02) | 0.87 (0.14) | 0.17 (0.01) |
| Q. serrata | 1.16 (0.04) | 0.57 (0.03) | 2.9 (0.43) | 0.08 (0.01) | 36.6 (2.8) | 14.2 (1.4) | 0.17 (0.03) | 0.37 (0.16) | 0.15 (0.05) | |
| p-value | 0.03 | 0.02 | 0.38 | 0.004 | 0.002 | 0.008 | 0.02 | 0.04 | 0.71 | |
| 20–30 | C. japonica | 1.26 (0.07) | 0.56 (0.04) | 2.4 (0.43) | 0.10 (0.02) | 24.4 (1.9) | 15.8 (0.7) | 0.22 (0.02) | 0.34 (0.07) | 0.09 (0.01) |
| Q. serrata | 1.24 (0.04) | 0.53 (0.03) | 2.1 (0.30) | 0.05 (0.01) | 39.3 (3.5) | 12.8 (1.6) | 0.18 (0.03) | 0.35 (0.12) | 0.20 (0.07) | |
| p-value | 0.80 | 0.55 | 0.57 | 0.04 | 0.004 | 0.12 | 0.23 | 0.95 | 0.15 | |
| Mean | C. japonica | 1.06 (0.04) | 0.50 (0.01) | 4.0 (0.46) | 0.21 (0.02) | 19.4 (0.7) | 18.8 (1.4) | 0.29 (0.02) | 1.53 (0.25) | 0.26 (0.02) |
| Q. serrata | 1.14 (0.02) | 0.54 (0.02) | 3.3 (0.33) | 0.11 (0.01) | 31.4 (1.0) | 14.8 (1.1) | 0.19 (0.03) | 0.50 (0.23) | 0.19 (0.06) | |
| p-value | 0.06 | 0.18 | 0.26 | 0.006 | 0.001 | 0.04 | 0.02 | 0.01 | 0.35 | |
| Stand | Depth (cm) | Dry Weight (kg ha–1) | C | N | C/N Ratio |
|---|---|---|---|---|---|
| (%) | |||||
| C. japonica | 4.0 (0.3) | 14,911 (1379) | 43.5 (2.3) | 0.84 (0.05) | 52.7 (4.7) |
| Q. serrata | 5.1 (0.5) | 18,500 (2343) | 45.5 (0.6) | 1.39 (0.10) | 34.1 (3.7) |
| p-value | 0.08 | 0.22 | 0.42 | 0.002 | 0.01 |
| Nutrient Stock | Forest Types | Tree Biomass | Forest Floor | Soils | References |
|---|---|---|---|---|---|
| (Mg ha−1) | |||||
| C | Cryptomeria japonica (p) | 52.0 | 6.5 | 59.11 | This study |
| Quercus serrata (n) | 25.8 | 8.5 | 51.10 | ||
| N | Cryptomeria japonica (p) | 0.20 | 0.13 | 3.02 | |
| Quercus serrata (n) | 0.20 | 0.26 | 1.64 | ||
| C | Douglas-fir (p) | - * | 11.3 | 46.8 | [10] |
| Beech (n) | - | 7.4 | 46.4 | ||
| N | Douglas-fir (p) | - | 0.3 | 3.4 | |
| Beech (n) | - | 0.2 | 3.6 | ||
| C | Plantation forests (p) | 79.5 | 5.1 | 103.9 | [11] |
| Pimary or secondary forests (n) | 121.1 | 6.1 | 128.8 | ||
| C | Eucalyptus saligna, Pinus spp. (p) | 195 | - | 78 | [9] |
| Macaranga conglomerate (n) | 360 | - | 146 | ||
| C | Pinus kesiya var. langbianensis (p) | 141.43 | 2.38 | 57.2 | [38] |
| Secondary coniferous forests (n) | 64.03 | 2.78 | 50.6 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baek, G.; Bae, E.-J.; Kim, C. A Stand-Level Comparison of Carbon and Nitrogen Distribution in an Exotic Japanese Cedar Plantation and a Natural Oak Stand. Forests 2021, 12, 963. https://doi.org/10.3390/f12080963
Baek G, Bae E-J, Kim C. A Stand-Level Comparison of Carbon and Nitrogen Distribution in an Exotic Japanese Cedar Plantation and a Natural Oak Stand. Forests. 2021; 12(8):963. https://doi.org/10.3390/f12080963
Chicago/Turabian StyleBaek, Gyeongwon, Eun-Ji Bae, and Choonsig Kim. 2021. "A Stand-Level Comparison of Carbon and Nitrogen Distribution in an Exotic Japanese Cedar Plantation and a Natural Oak Stand" Forests 12, no. 8: 963. https://doi.org/10.3390/f12080963
APA StyleBaek, G., Bae, E.-J., & Kim, C. (2021). A Stand-Level Comparison of Carbon and Nitrogen Distribution in an Exotic Japanese Cedar Plantation and a Natural Oak Stand. Forests, 12(8), 963. https://doi.org/10.3390/f12080963