
Effect of Climate Change on the Growth of Endangered Scree Forests in Krkonoše National Park (Czech Republic)


 , ,
, ,
Abstract
1. Introduction
2. Material and Methods
2.1. Study Site
2.2. Data Collection
2.3. Data Analysis
3. Results
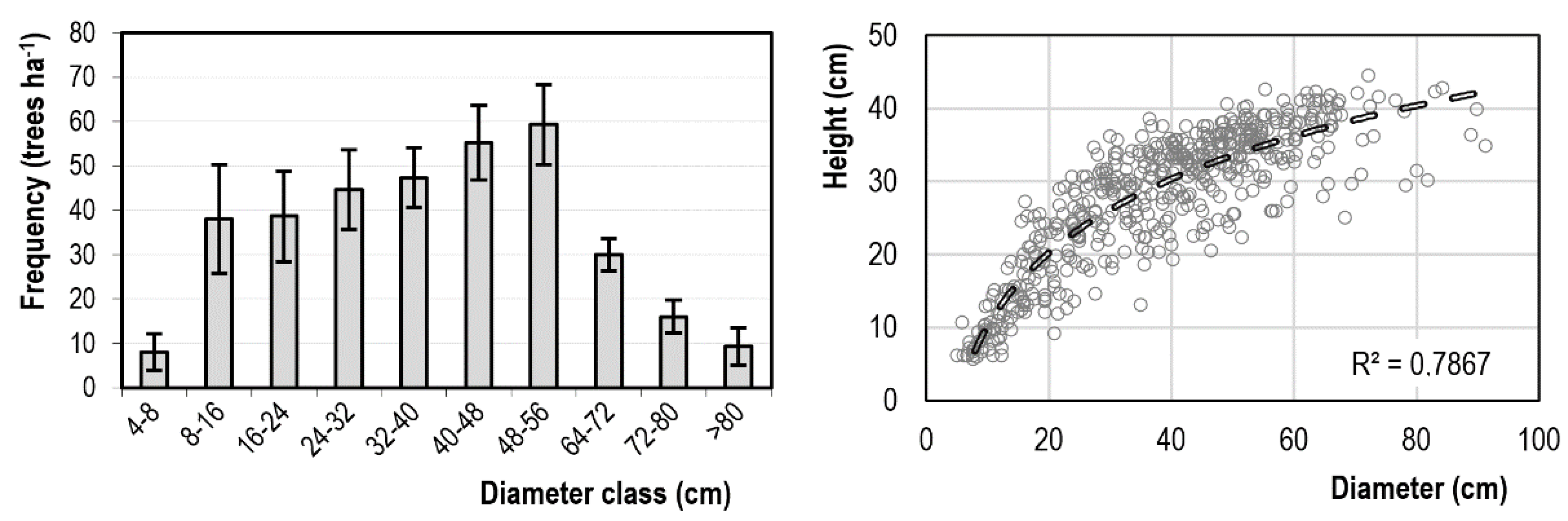
3.1. Stand Structure and Production
3.2. Tree Layer Biodiversity
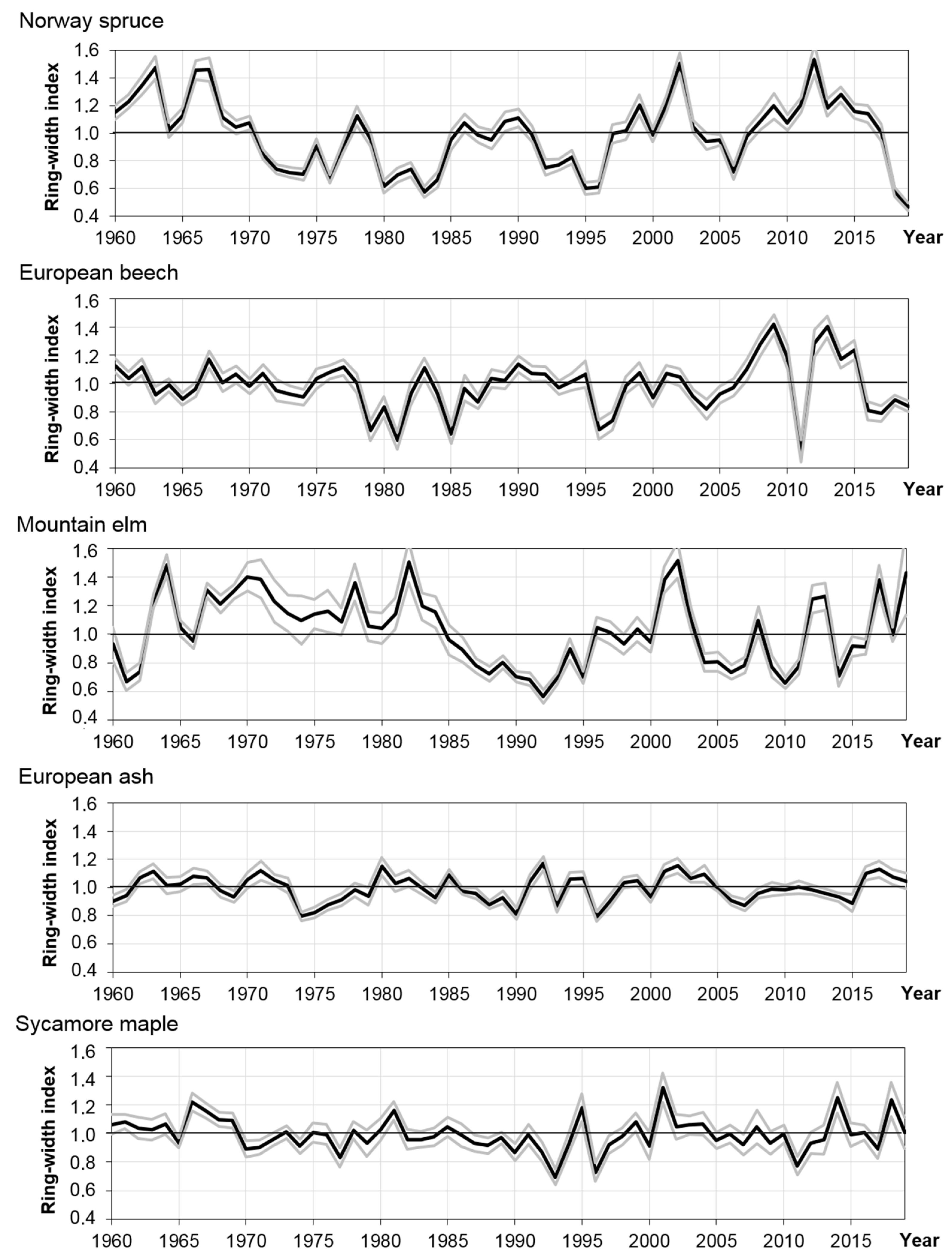
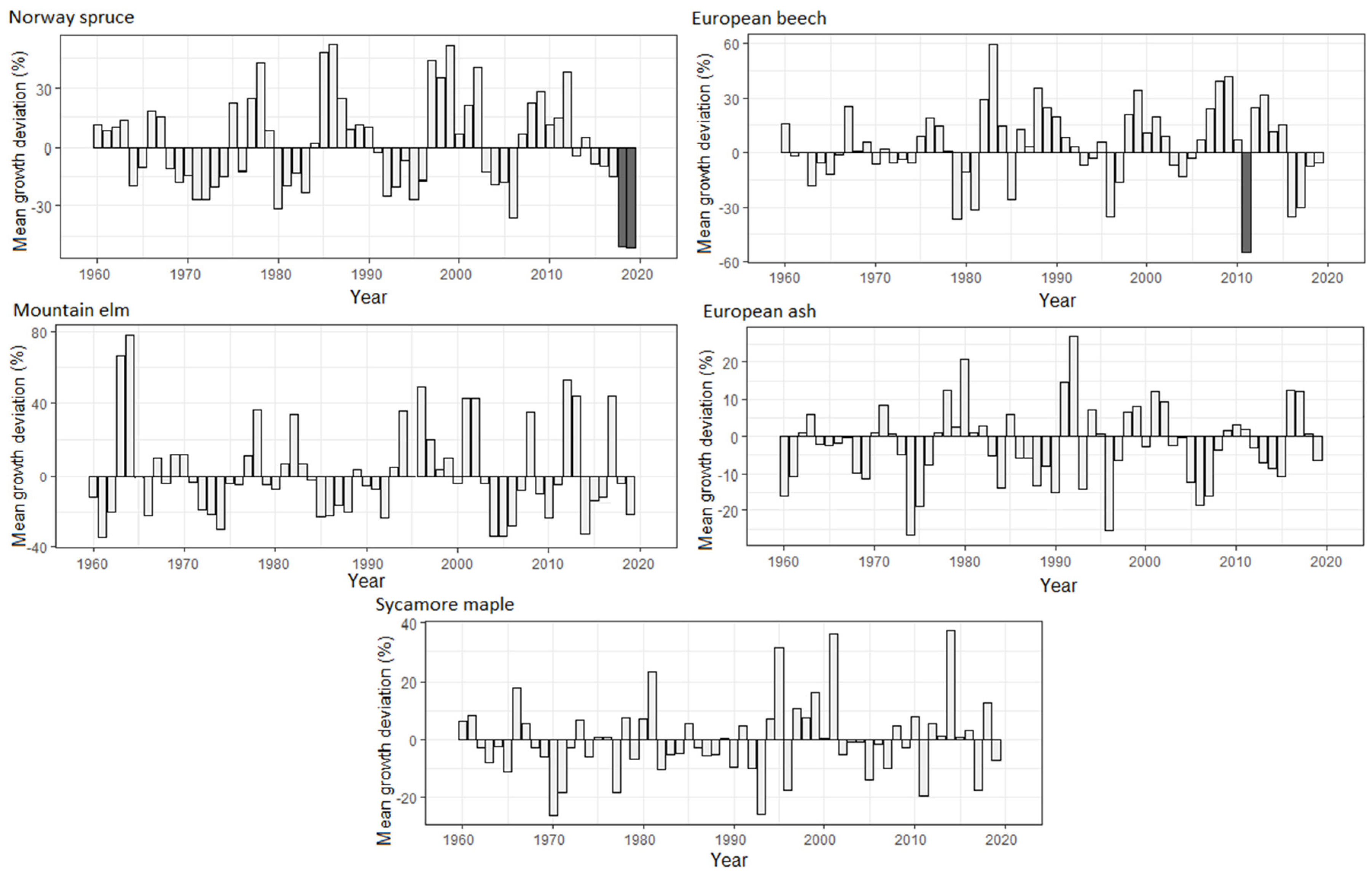
3.3. Dynamics of Radial Growth
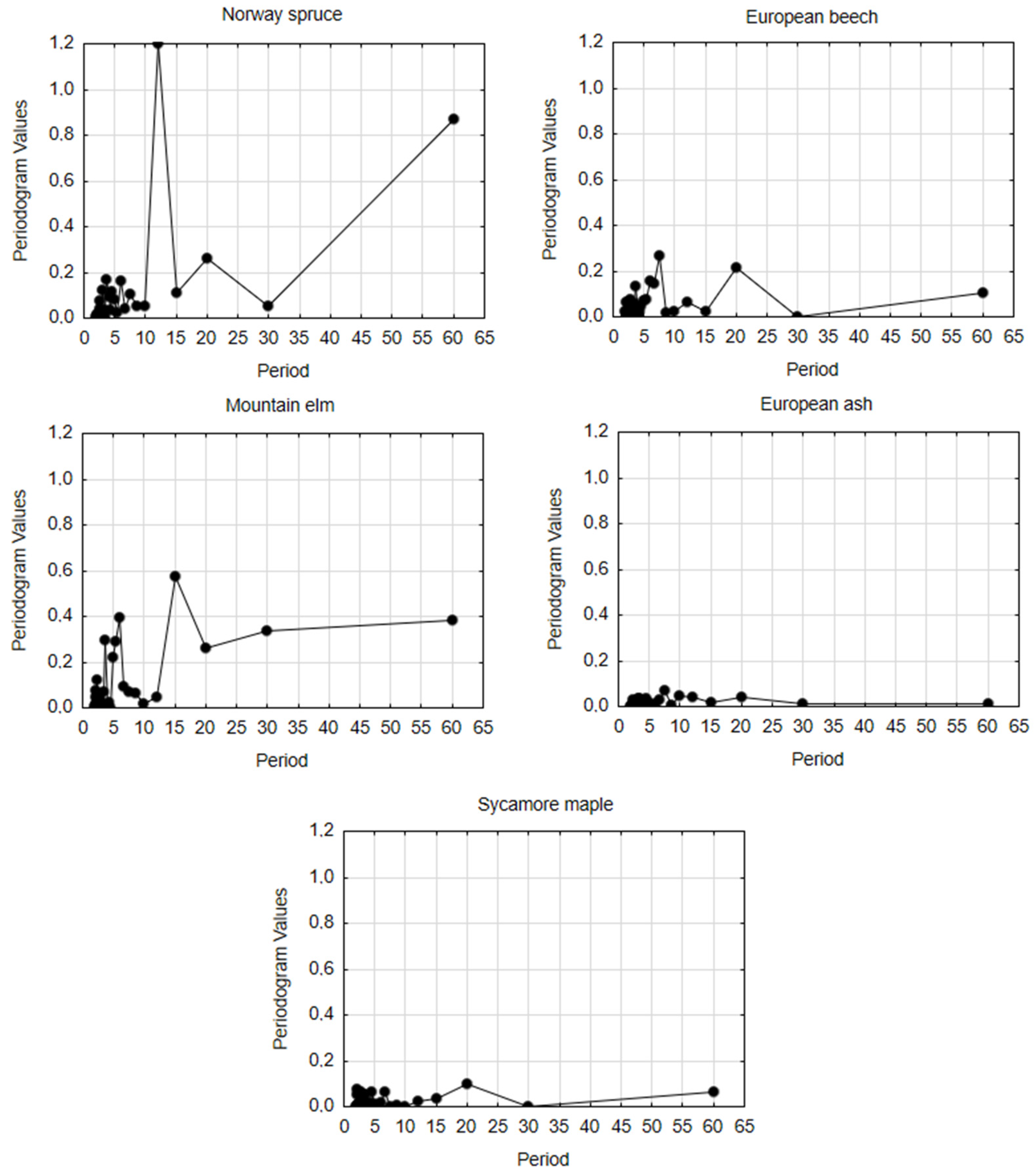
3.4. Spectral Analysis in Radial Growth
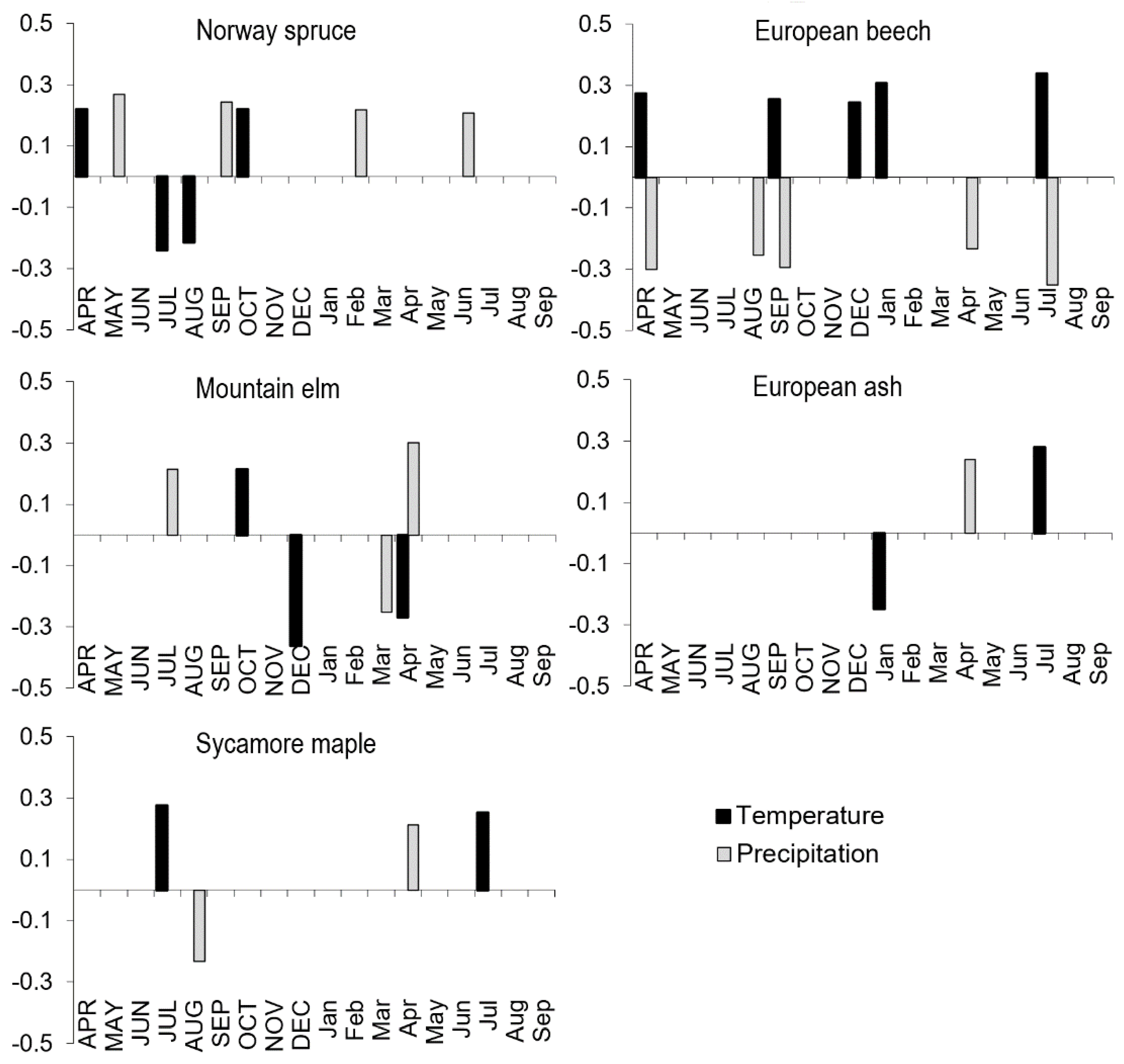
3.5. Effect of Climate on Radial Growth
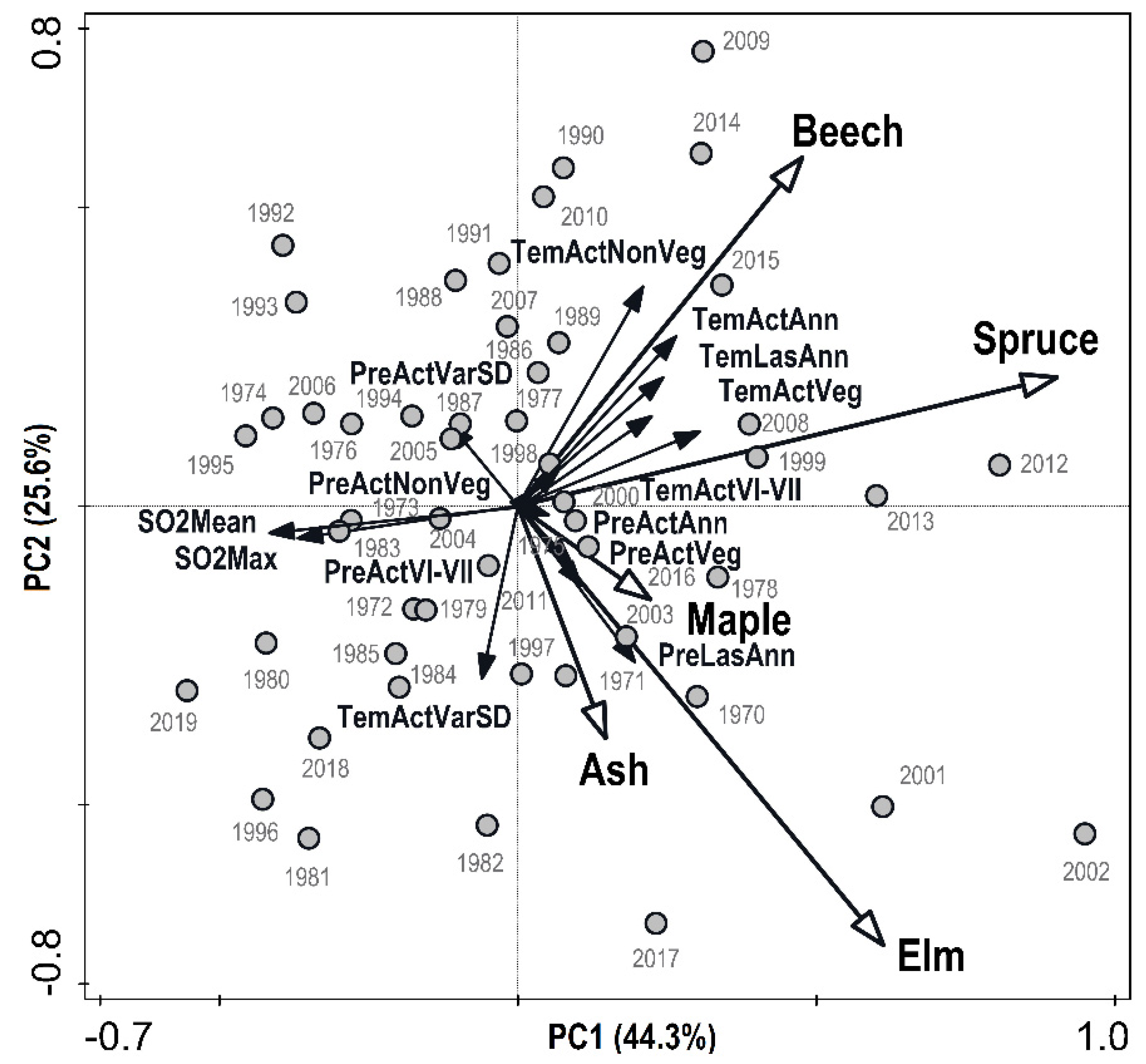
3.6. Interaction between Climate, Air pollution, and Radial Growth of Tree Species
4. Discussion
4.1. Stand Structure and Biodiversity
4.2. Effect of Non-Climatic Factors to Radial Growth
4.3. Effect of Climate to Radial Growth
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Bošela, M.; Štefančík, I.; Petráš, R.; Vacek, S. The effects of climate warming on the growth of European beechforests depend critically on thinning strategy and site productivity. Agric. For. Meteorol. 2016, 222, 21–31. [Google Scholar] [CrossRef]
- Brázdil, R.; Trnka, M.; Dobrovolný, P.; CHromá, K.; Hlavinka, P.; Žalud, Z. Variability of droughts in the Czech Republic, 1881–2006. Theor. Appl. Climatol. 2009, 97, 297–315. [Google Scholar] [CrossRef]
- Kolström, M.; Lindner, M.; Vilén, T.; Maroschek, M.; Seidl, R.; Lexer, M.J.; Netherer, S.; Kremer, A.; Delzon, S.; Barbati, A.; et al. Reviewing the science and implementation of climate change adaptation measures in European forestry. Forests 2011, 2, 961–982. [Google Scholar] [CrossRef]
- Seidl, R.; Rammer, W.; Lexer, M.J. Climate change vulnerability of sustainable forest management in the Eastern Alps. Clim. Chang. 2011, 106, 225–254. [Google Scholar] [CrossRef]
- Martín-Martín, C.; Bunce, R.G.H.; Saura, S.; Elena-Rosselló, R. Changes and interactions between forest landscape connectivity and burnt area in Spain. Ecol. Indic. 2013, 33, 129–138. [Google Scholar] [CrossRef]
- Šimůnek, V.; Sharma, R.P.; Vacek, Z.; Vacek, S.; Hůnová, I. Sunspot area as unexplored trend inside radial growth of European beech in Krkonoše Mountains: A forest science from different perspective. Eur. J. For. Res. 2020, 139, 999–1013. [Google Scholar] [CrossRef]
- Cukor, J.; Vacek, Z.; Linda, R.; Bílek, L. Carbon sequestration in soil following afforestation of former agricultural land in the Czech Republic. Cent. Eur. For. J. 2017, 63, 97–104. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-resolution global maps of 21st-century forest cover change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef]
- Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions. New EU Forest Strategy for 2030; European Commission: Brussels, Belgium, 2021; 27p, Available online: https://ec.europa.eu/info/sites/default/files/communication-new-eu-forest-strategy-2030_with-annex_en.pdf (accessed on 19 August 2021).
- European Union Forest Action Plan; EUR-Lex—Europa: Brussels, Belgium, 2006. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=LEGISSUM:l24277 (accessed on 19 August 2021).
- Brus, D.J.; Hengeveld, G.M.; Walvoort, D.J.J.; Goedhart, P.W.; Heidema, A.H.; Nabuurs, G.J.; Gunia, K. Statistical mapping of tree species over Europe. Eur. J. For. Res. 2012, 131, 145–157. [Google Scholar] [CrossRef]
- Sabatini, F.M.; Burrascano, S.; Keeton, W.S.; Levers, C.; Lindner, M.; Pötzschner, F.; Verkerk, P.J.; Bauhus, J.; Buchwald, E.; Chaskovsky, O.; et al. Where are Europe’s last primary forests? Divers. Distrib. 2018, 24, 1–14. [Google Scholar] [CrossRef]
- Ott, E.; Frehner, M.; Frey, H.U.; Lüscher, P. Gebirgsnadelwäulder: Ein Praxisorientierter Leitfaden für Eine Standortsgerechte Waldbehandlung; Verlag Paul Haupt: Bern, Switzerland; Stuttgart, Germany; Wien, Austria, 1997. [Google Scholar]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of drought tolerance in major European temperate forest trees: Climatic drivers and levels of variability. Glob. Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef] [PubMed]
- Klopčič, M.; Mina, M.; Bugmann, H.; Bončina, A. The prospects of silver fir (Abies alba Mill.) and Norway spruce (Picea abies (L.) Karst) in mixed mountain forests under various management strategies, climate change and high browsing pressure. Eur. J. For. Res. 2017, 136, 1071–1090. [Google Scholar] [CrossRef]
- Diaci, J.; Rozman, J.; Rozman, A. Regeneration gap and microsite niche artitioning in a high alpine forest: Are Norway spruce seedlings more drought-tolerant than beech seedlings? For. Ecol. Manag. 2020, 455, 117688. [Google Scholar] [CrossRef]
- Slanař, J.; Vacek, Z.; Vacek, S.; Bulušek, D.; Cukor, J.; Štefančík, I.; Bílek, L.; Krl, J. Long-term transformation of submontane spruce-beech forests in the Jizerské hory Mts.: Dynamics of natural regeneration. Cent. Eur. For. J. 2017, 63, 212–224. [Google Scholar] [CrossRef]
- Štefančík, I.; Štefančík, L. Effect of long-term tending on qualitative and quantitative production in mixed stands of spruce, fir and beech on Motyčky research plot. J. For. Sci. 2003, 49, 108–124. [Google Scholar] [CrossRef]
- Knoke, T.; Ammer, C.; Stimm, B.; Mosandl, R. Admixing broadleaved to coniferous tree species: A review on yield, ecological stability and economics. Eur. J. For. Res. 2008, 127, 89–101. [Google Scholar] [CrossRef]
- Ruosch, M.; Spahni, R.; Joos, F.; Henne, P.D.; van der Knaap, W.O.; Tinner, W. Past and future evolution of Abies alba forests in Europe—Comparison of a dynamic vegetation model with palaeo data and observations. Glob. Chang. Biol. 2016, 22, 727–740. [Google Scholar] [CrossRef]
- Mikulenka, P.; Prokůpková, A.; Vacek, Z.; Vacek, S.; Bulušek, D.; Simon, J.; Šimůnek, V.; Hájek, V. Effect of climate and air pollution on radial growth of mixed forests: Abies alba Mill. vs. Picea abies (L.) Karst. Cent. Eur. For. J. 2020, 66, 23–36. [Google Scholar] [CrossRef]
- Chamagne, J.; Tanadini, M.; Frank, D.; Matula, R.; Paine, C.E.T.; Philipson, C.D.; Svátek, M.; Turnbull, L.A.; Volařík, D.; Hector, A. Forest diversity promotes individual tree growth in central European forest stands. J. Appl. Ecol. 2017, 54, 71–79. [Google Scholar] [CrossRef]
- Jactel, H.; Gritti, E.S.; Drössler, L.; Forrester, D.I.; Mason, W.L.; Morin, X.; Pretzsch, H.; Castagneyrol, B. Positive biodiversity–Productivity relationships in forests: Climate matters. Biol. Lett. 2018, 14, 20170747. [Google Scholar] [CrossRef]
- Bošela, M.; Tobin, B.; Šebeň, V.; Petráš, R.; Larocque, G.R.; Šeben, V.; Petráš, R.; Larocque, G.R. Different mixtures of Norway spruce, silver fir, and European beech modify competitive interactions in central Europeanmaturemixed forests. Can. J. For. Res. 2015, 45, 1577–1586. [Google Scholar] [CrossRef]
- Toïgo, M.; Vallet, P.; Perot, T.; Bontemps, J.D.; Piedallu, C.; Courbaud, B. Overyielding in mixed forests decreases with site productivity. J. Ecol. 2015, 103, 502–512. [Google Scholar] [CrossRef]
- Metz, J.; Annighöfer, P.; Schall, P.; Zimmermann, J.; Kahl, T.; Schulze, E.D.; Ammer, C. Site-adapted admixed tree species reduce drought susceptibility of mature European beech. Glob. Chang. Biol. 2016, 22, 903–920. [Google Scholar] [CrossRef]
- Vitali, V.; Büntgen, U.; Bauhus, J. Silver fir and Douglas fir are more tolerant to extreme droughts than Norway spruce in south-western Germany. Glob. Chang. Biol. 2017, 23, 5108–5119. [Google Scholar] [CrossRef] [PubMed]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidl, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Fyllas, N.M.; Christopoulou, A.; Galanidis, A.; Michelaki, C.Z.; Dimitrakopoulos, P.G.; Fulé, P.Z.; Arianoutsou, M. Tree growth-climate relationships in a forest-plot network on Mediterranean mountains. Sci. Total Environ. 2017, 598, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Kolář, T.; Čermák, P.; Trnka, M.; Žid, T.; Rybníček, M. Temporal changes in the climate sensitivity of Norway spruce and European beech along an elevation gradient in Central Europe. Agric. For. Meteorol. 2017, 239, 24–33. [Google Scholar] [CrossRef]
- Taylor, A.R.; Boulanger, Y.; Price, D.T.; Cyr, D.; McGarrigle, E.; Rammer, W.; Kershaw, J.A. Rapid 21st century climate change projected to shift composition and growth of Canada’s Acadian Forest Region. For. Ecol. Manag. 2017, 405, 284–294. [Google Scholar] [CrossRef]
- Vacek, S.; Prokupková, A.; Vacek, Z.; Bulu ek, D.; Simunek, V.; Králícek, I.; Prausová, R.; Hájek, V. Growth response of mixed beech forests to climate change, various management and game pressure in Central Europe. J. For. Sci. 2019, 65, 116–129. [Google Scholar] [CrossRef]
- Hájek, V.; Vacek, Z.; Vacek, S.; Bílek, L.; Prausová, R.; Linda, R.; Bulušek, D.; Králíček, I. Changes in diversity of protected scree and herb-rich beech forest ecosystems in Central Europe over 55 years. Cent. Eur. For. J. 2020, 66, 202–2017. [Google Scholar] [CrossRef]
- Schütz, J.P.; Götz, M.; Schmid, W.; Mandallaz, D. Vulnerability of spruce (Picea abies) and beech (Fagus sylvatica) forest stands to storms and consequences for silviculture. Eur. J. For. Res. 2006, 125, 291–302. [Google Scholar] [CrossRef]
- Bulušek, D.; Vacek, Z.; Vacek, S.; Král, J.; Bílek, L.; Králíček, I. Spatial pattern of relict beech (Fagus sylvatica L.) forests in the Sudetes of the Czech Republic and Poland. J. For. Sci. 2016, 62, 293–305. [Google Scholar] [CrossRef]
- Solberg, S.; Dobbertin, M.; Reinds, G.J.; Lange, H.; Andreassen, K.; Fernandez, P.G.; Hildingsson, A.; de Vries, W. Analyses of the impact of changes in atmospheric deposition and climate on forest growth in European monitoring plots: A stand growth approach. For. Ecol. Manag. 2009, 258, 1735–1750. [Google Scholar] [CrossRef]
- Vacek, S.; Hůnová, I.; Vacek, Z.; Hejcmanová, P.; Podrázský, V.; Král, J.; Putalová, T.; Moser, W.K. Effects of air pollution and climatic factors on Norway spruce forests in the Orlické hory Mts. (Czech Republic), 1979–2014. Eur. J. For. Res. 2015, 134, 1127–1142. [Google Scholar] [CrossRef]
- Vacek, Z.; Vacek, S.; Slanař, J.; Bílek, L.; Bulušek, D.; Štefančík, I.; Králíček, I.; Vančura, K. Adaption of Norway spruce and European beech forests under climate change: From resistance to close-to-nature silviculture. Cent. Eur. For. J. 2019, 65, 129–144. [Google Scholar] [CrossRef]
- Zapletal, M.; Juran, S.; Krpes, V.; Michna, K.; Cudlin, P.; Edwards, M. Effect of ozone flux on selected structural and antioxidant characteristics of a mountain norway spruce forest. Balt. For. 2018, 24, 261–267. [Google Scholar]
- Vacek, S.; Podrázský, V.; Mikeska, M.; Moser, W.K. Introskeletal erosion threat in mountain forests of the Czech Republic. J. For. Sci. 2003, 49, 313–320. [Google Scholar] [CrossRef]
- Machar, I.; Vlckova, V.; Bucek, A.; Vozenilek, V.; Salek, L.; Jerabkova, L. Modelling of climate conditions in forest vegetation zones as a support tool for forest management strategy in European beech dominated forests. Forests 2017, 8, 82. [Google Scholar] [CrossRef]
- Castagneri, D.; Battipaglia, G.; Von Arx, G.; Pacheco, A.; Carrer, M. Tree-ring anatomy and carbon isotope ratio show both direct and legacy effects of climate on bimodal xylem formation in Pinus pinea. Tree Physiol. 2018, 38, 1098–1109. [Google Scholar] [CrossRef] [PubMed]
- Keenan, T.F.; Gray, J.; Friedl, M.A.; Toomey, M.; Bohrer, G.; Hollinger, D.Y.; Munger, J.W.; O’Keefe, J.; Schmid, H.P.; Wing, I.S.; et al. Net carbon uptake has increased through warming-induced changes in temperate forest phenology. Nat. Clim. Chang. 2014, 4, 598. [Google Scholar] [CrossRef]
- Babushkina, E.; Zhirnova, D.; Belokopytova, L.; Vaganov, E. Warming induced changes in wood matter accumulation in tracheid walls of spruce. J. Mt. Sci. 2020, 17, 16–30. [Google Scholar] [CrossRef]
- Begović, K.; Rydval, M.; Mikac, S.; Čupić, S.; Svobodova, K.; Mikoláš, M.; Kozák, D.; Kameniar, O.; Frankovič, M.; Pavlin, J.; et al. Climate-growth relationships of Norway Spruce and silver fir in primary forests of the Croatian Dinaric mountains. Agric. For. Meteorol. 2020, 288–289, 108000. [Google Scholar] [CrossRef]
- Yousefpour, R.; Temperli, C.; Jacobsen, J.B.; Thorsen, B.J.; Meilby, H.; Lexer, M.J.; Lindner, M.; Bugmann, H.; Borges, J.G.; Palma, J.H.N.; et al. A framework for modeling adaptive forest management and decision making under climate change. Ecol. Soc. 2017, 22, 40. [Google Scholar] [CrossRef]
- Vacek, Z.; Prokůpková, A.; Vacek, S.; Cukor, J.; Bílek, L.; Gallo, J.; Bulušek, D. Silviculture as a tool to support stability and diversity of forests under climate change: Study from Krkonoše Mountains. Cent. Eur. For. J. 2020, 66, 116–129. [Google Scholar] [CrossRef]
- Podrázský, V.; Vacek, S.; Vacek, Z.; Raj, A.; Mikeska, M.; Boček, M.; Schwarz, O.; Hošek, J.; Šach, F.; Černohous, V.; et al. Půdy Lesů a Ekosystémů nad Horní Hranicí Lesa v Národních Parcích Krkonoš; Kostelec nad Černými lesy, Lesnická práce, s. r. o.: Kostelec nad Černými lesy, Czech Republic, 2010. [Google Scholar]
- Neuhäuslová, Z.; Blažková, D.; Grulich, V.; Husová, M.; Chytrý, M.; Jeník, J.; Jirásek, J.; Kolbek, J.; Kropáč, Z.; Ložek, V.; et al. Mapa Potenciální Přirozené Vegetace České Republiky; Academia: Praha, Czech Republic, 1998. [Google Scholar]
- Chytrý, M.; Kučera, T.; Kočí, M.; Grulich, V.; Lustyk, P. (Eds.) Katalog Biotopů České Republiky; Agentura Ochrany Přírody a Krajiny ČR: Praha, Czech Republic, 2010. [Google Scholar]
- Vacek, S.; Vacek, Z.; Kalousková, I.; Cukor, J.; Bílek, L.; Moser, W.K.; Bulušek, D.; Podrázský, V.; Řeháček, D. Sycamore maple (Acer pseudoplatanus L.) stands on former agricultural land in the sudetes—Evaluation of ecological value and production potential. Dendrobiology 2018, 79, 61–76. [Google Scholar] [CrossRef]
- Vacek, S.; Moucha, P.; Bılek, L.; Mikeska, M. Péče o Lesní Ekosystémy v Chráněných Územích ČR; Ministerstvo Životního Prostředí: Praha, Czech Republic, 2012.
- Bodziarczyk, J.; Szwagrzyk, J. Species composition and structure of forest stands in Phyllitido-Aceretum community. Ekol. Pol. 1995, 43, 153–173. [Google Scholar]
- Paillet, Y.; Archaux, F.; Breton, V.; Brun, J.J. A quantitative assessment of the ecological value of sycamore maple habitats in the French Alps. Ann. For. Sci. 2008, 65, 713. [Google Scholar] [CrossRef][Green Version]
- Baran, J.; Bodziarczyk, J. Forest stands in Phyllitido-Aceretum community occuring on limestone rocks and shelves–comparison of their structure in strictly protected and managed forests. Pol. J. Ecol. 2019, 67, 105–121. [Google Scholar] [CrossRef]
- Baran, J.; Pielech, R.; Kauzal, P.; Kukla, W.; Bodziarczyk, J. Influence of forest management on stand structure in ravine forests. For. Ecol. Manag. 2020, 463, 118018. [Google Scholar] [CrossRef]
- Putalová, T.; Vacek, Z.; Vacek, S.; Štefančík, I.; Bulušek, D.; Král, J. Tree-ring widths as an indicator of air pollution stress and climate conditions in different Norway spruce forest stands in the Krkonoše Mts. Cent. Eur. For. J. 2019, 65, 21–33. [Google Scholar] [CrossRef]
- Köppen, W. Grundriss der Klimakunde. Zweite, Verbesserte Auflage der Klimate der Erde; Walter de Gruyter & Co.: Berlin, Germany, 1931. [Google Scholar]
- Král, J.; Vacek, S.; Vacek, Z.; Putalová, T.; Bulušek, D.; Štefančík, I. Structure, development and health status of spruce forests affected by air pollution in the western Krkonoše Mts. in 1979–2014. Cent. Eur. For. J. 2015, 61, 175–187. [Google Scholar] [CrossRef]
- Vacek, Z.; Prokůpková, A.; Vacek, S.; Bulušek, D.; Šimůnek, V.; Hájek, V.; Králíček, I. Mixed vs. monospecific mountain forests in response to climate change: Structural and growth perspectives of Norway spruce and European beech. For. Ecol. Manag. 2021, 488, 119019. [Google Scholar] [CrossRef]
- Fabrika, M.; Ďurský, J. Stromové Růstové Simulátory; EFRA: Zvolen, Slovakia, 2005. [Google Scholar]
- Petráš, R.; Pajtík, J. Sústava česko-slovenských objemových tabuliek drevín. Lesn. Časopis 1991, 37, 49–56. [Google Scholar]
- Reineke, L.H. Perfecting a stand-density index for even-aged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Crookston, N.L.; Stage, A.R. Percentcanopy Cover and Stand Structural Statistics Fromthe Forest Vegetation Simulator; RMRS GTR-24; USDA Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 1999.
- Halaj, J.; Řehák, J. Rastové Tabuľky Hlavných Drevín ČSSR [Yield Tables of the Main Tree Species of Czechoslovakia]; Príroda: Bratislava, Slovakia, 1987. [Google Scholar]
- Sharma, R.P.; Vacek, Z.; Vacek, S. Modeling individual tree height to diameter ratio for Norway spruce and European beech in Czech Republic. Trees 2016, 30, 1969–1982. [Google Scholar] [CrossRef]
- Sharma, R.P.; Vacek, Z.; Vacek, S.; Kučera, M. A nonlinear mixed-effects height-to-diameter ratio model for several tree species based on Czech national forest inventory data. Forests 2019, 10, 70. [Google Scholar] [CrossRef]
- Slodicak, M.; Novak, J. Silvicultural measures to increase the mechanical stability of pure secondary Norway spruce stands before conversion. For. Ecol. Manag. 2006, 224, 252–257. [Google Scholar] [CrossRef]
- Margalef, R. Information theory in ecology. Gen. Syst. 1958, 3, 36–71. [Google Scholar]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Pielou, E.C. Ecological Diversity; Wiley: New York, NY, USA, 1975. [Google Scholar]
- Clark, P.J.; Evans, F.C. Distance to Nearest Neighbor as a Measure of Spatial Relationships in Populations. Ecology 1954, 35, 445–453. [Google Scholar] [CrossRef]
- Pretzsch, H. Wissen nutzbar machen für das Management von Waldökosystemen. Allg. Forstz. Der Wald 2006, 61, 1158–1159. [Google Scholar]
- Jaehne, S.; Dohrenbusch, A. Ein Verfahren zur Beurteilung der Bestandesdiversität. Eur. J. For. Res. 1997, 116, 333–345. [Google Scholar] [CrossRef]
- Füldner, K. Strukturbeschreibung in Mischbeständen. Forstarchiv 1995, 66, 235–606. [Google Scholar]
- Bílek, L.; Remeš, J.; Zahradnik, D. Managed versus unmanaged. Structure of beech forest stands “Fagus sylvatica L.” after 50 years of development, Central Bohemian. For. Syst. 2011, 20, 122–138. [Google Scholar] [CrossRef]
- Larsson, L.-A. Cybis Elektronik & Data AB, Saltsjöbaden, Sweden, 2010. Available online: http://www.cybis.se/ (accessed on 19 August 2021).
- Team R Core. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Bunn, A.; Korpela, M. Chronology Building in dplR; R Foundation for Statistical Computing: Vienna, Austria, 2018; pp. 1–13. [Google Scholar]
- Bunn, A.G. Statistical and visual crossdating in R using the dplR library. Dendrochronologia 2010, 28, 251–258. [Google Scholar] [CrossRef]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Cook, E.R.; Shiyatov, S.G.; Mazepa, V.S.; Ecology, A.; Branch, U. Methods of Dendrochronology Applications; Tree-Ring Laboratory, Lamont-Ooherty Geological Observatory, Columbia University: New York, NY, USA, 1990. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press: Tucson, AZ, USA, 1976. [Google Scholar]
- Speer, J.H. Fundamentals of Tree-Ring Research. In Geoarchaeology; University of Arizona Press: Tuscon, AZ, USA, 2010. [Google Scholar]
- Schweingruber, F.H.; Eckstein, D.; Serre-Bachet, F.; Braker, O.U. Identification, Presentation and Interpretation of Event Years and Pointer Years in Dendrochronology. Dendrochronologia 1990, 8, 9–38. [Google Scholar]
- Neuwirth, B.; Schweingruber, F.H.; Winiger, M. Spatial patterns of central European pointer years from 1901 to 1971. Dendrochronologia 2007, 24, 79–89. [Google Scholar] [CrossRef]
- Biondi, F.; Waikul, K. Dendroclim 2002: AC++ program for statistical calibration of climate signals in tree ring chronologie. Comput. Geosci. 2004, 30, 303–311. [Google Scholar] [CrossRef]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using Canoco 5; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Vacek, Z.; Vacek, S.; Bílek, L.; Král, J.; Ulbrichová, I.; Simon, J.; Bulušek, D. Impact of applied silvicultural systems on spatial pattern of hornbeam-oak forests. Cent. Eur. For. J. 2018, 64, 33–45. [Google Scholar] [CrossRef]
- del Río, M.; Schütze, G.; Pretzsch, H. Temporal variation of competition and facilitation in mixed species forests in Central Europe. Plant Biol. 2014, 16, 166–176. [Google Scholar] [CrossRef]
- Vacek, S.; Vacek, Z.; Podrázský, V.; Bílek, L.; Bulušek, D.; Štefančík, I.; Remeš, J.; Štícha, V.; Ambrož, R. Structural diversity of autochthonous beech forests in Broumovské stěny National Nature Reserve, Czech Republic. Austrian J. For. Sci. 2014, 131, 191–214. [Google Scholar]
- Ghalandarayeshi, S.; Nord-Larsen, T.; Johannsen, V.K.; Larsen, J.B. Spatial patterns of tree species in Suserup Skov—A semi-natural forest in Denmark. For. Ecol. Manag. 2017, 406, 391–401. [Google Scholar] [CrossRef]
- Bílek, L.; Remeš, J.; Podrázský, V.; Rozenbergar, D.; Diaci, J.; Zahradnik, D. Gap regeneration in near-natural European beech forest stands in Central Bohemia—The role of heterogeneity and micro-habitat factors. Dendrobiology 2014, 71, 59–71. [Google Scholar] [CrossRef]
- Vacek, S.; Vacek, Z.; Bílek, L.; Simon, J.; Remeš, J.; Hůnová, I.; Král, J.; Putalová, T.; Mikeska, M. Structure, regeneration and growth of scots pine (Pinus sylvestris L.) stands with respect to changing climate and environmental pollution. Silva Fenn. 2016, 50, 1564. [Google Scholar] [CrossRef]
- Kang, H.; Zheng, Y.; Liu, S.; Chai, Z.; Chang, M.; Hu, Y.; Li, G.; Wang, D. Population structure and spatial pattern of predominant tree species in a pine-oak mosaic mixed forest in the qinling mountains, China. J. Plant Interact. 2017, 12, 78–86. [Google Scholar] [CrossRef]
- Králíček, I.; Vacek, Z.; Vacek, S.; Remeš, J.; Bulušek, D.; Král, J.; Štefančík, I.; Putalová, T. Dynamics and structure of mountain autochthonous spruce-beech forests: Impact of hilltop phenomenon, air pollutants and climate. Dendrobiology 2017, 77, 121–139. [Google Scholar] [CrossRef]
- Heiri, C.; Wolf, A.; Rohrer, L.; Bugmann, H. Forty years of natural dynamics in Swiss beech forests: Structure, composition, and the influence of former management. Ecol. Appl. 2009, 19, 1920–1934. [Google Scholar] [CrossRef]
- Horvat, V.; De Vicuña, J.G.; Biurrun, I.; García-Mijangos, I. Managed and unmanaged silver fir-beech forests show similar structural features in the western pyrenees. iForest 2018, 11, 698–704. [Google Scholar] [CrossRef]
- Altman, J.; Hédl, R.; Szabó, P.; Mazůrek, P.; Riedl, V.; Müllerová, J.; Kopecký, M.; Doležal, J. Tree-Rings Mirror Management Legacy: Dramatic Response of Standard Oaks to Past Coppicing in Central Europe. PLoS ONE 2013, 8, e55770. [Google Scholar] [CrossRef]
- Trouvé, R.; Bontemps, J.D.; Seynave, I.; Collet, C.; Lebourgeois, F. Stand density, tree social status and water stress influence allocation in height and diameter growth of Quercus petraea (Liebl.). Tree Physiol. 2015, 35, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Burrascano, S.; Keeton, W.S.; Sabatini, F.M.; Blasi, C. Commonality and variability in the structural attributes of moist temperate old-growth forests: A global review. For. Ecol. Manag. 2013, 291, 458–479. [Google Scholar] [CrossRef]
- Vacek, S.; Vacek, Z.; Bílek, L.; Nosková, I.; Schwarz, O. Structure and development of forest stands on permanent research plots in the Krkonoše Mts. J. For. Sci. 2010, 56, 518–530. [Google Scholar] [CrossRef]
- Wesely, N.; Fraver, S.; Kenefic, L.; Weiskittel, A.; Ruel, J.-C.; Thompson, M.; White, A. Structural attributes of old-growth and partially harvested northern whitecedar stands in Northeastern North America. Forests 2018, 9, 376. [Google Scholar] [CrossRef]
- Burton, J.I.; Zenner, E.K.; Frelich, L.E.; Cornett, M.W. Patterns of plant community structure within and among primary and second-growth northern hardwood forest stands. For. Ecol. Manag. 2009, 258, 2556–2568. [Google Scholar] [CrossRef]
- Commarmot, B.; Bachofen, H.; Bundziak, Y.; Bürgi, A.; Ramp, B.; Shparyk, Y.; Sukhariuk, D.; Viter, R.; Zingg, A. Structures of virgin and managed beech forests in Uholka (Ukraine) and Sihlwald (Switzerland): A comparative study. Proc. For. Snow Landsc. Res. 2005, 79, 45–56. [Google Scholar]
- Dieler, J.; Uhl, E.; Biber, P.; Müller, J.; Rötzer, T.; Pretzsch, H. Effect of forest stand management on species composition, structural diversity, and productivity in the temperate zone of Europe. Eur. J. For. Res. 2017, 136, 739–766. [Google Scholar] [CrossRef]
- Vannoppen, A.; Kint, V.; Ponette, Q.; Verheyen, K.; Muys, B. Tree species diversity impacts average radial growth of beech and oak trees in belgium, not their long-term growth trend. For. Ecosyst. 2019, 6, 10. [Google Scholar] [CrossRef]
- Tognetti, R.; Lasserre, B.; Di Febbraro, M.; Marchetti, M. Modeling regional drought-stress indices for beech forests in Mediterranean mountains based on tree-ring data. Agric. For. Meteorol. 2019, 265, 110–120. [Google Scholar] [CrossRef]
- Bošela, M.; Lukac, M.; Castagneri, D.; Sedmák, R.; Biber, P.; Carrer, M.; Konôpka, B.; Nola, P.; Nagel, T.A.; Popa, I.; et al. Contrasting effects of environmental change on the radial growth of co-occurring beech and fir trees across Europe. Sci. Total Environ. 2018, 615, 1460–1469. [Google Scholar] [CrossRef]
- Bošela, M.; Tumajer, J.; Cienciala, E.; Dobor, L.; Kulla, L.; Marčiš, P.; Popa, I.; Sedmák, R.; Sedmáková, D.; Sitko, R.V.; et al. Climate warming induced synchronous growth decline in Norway spruce populations across biogeographical gradients since 2000. Sci. Total Environ. 2020, 752, 141794. [Google Scholar] [CrossRef]
- UNECE. FAO State of Europe’s forests 2011. In Status & Trends in Sustainable Forest Management in Europe; UNECE, FAO: Rome, Italy, 2011. [Google Scholar]
- Šimůnek, V.; Vacek, Z.; Vacek, S.; Králíček, I.; Vančura, K. Growth variability of European beech (Fagus sylvatica L.) natural forests: Dendroclimatic study from Krkonoše National Park. Cent. Eur. For. J. 2019, 65, 3–11. [Google Scholar] [CrossRef]
- Krejcí, F.; Vacek, S.; Bílek, L.; Mikeska, M.; Hejcmanová, P.; Vacek, Z. The effects of climatic conditions and forest site types on disintegration rates in Picea abies occurring at the Modrava Peat Bogs in the Šumava National Park. Dendrobiology 2013, 70, 35–44. [Google Scholar] [CrossRef]
- Vacek, Z.; Vacek, S.; Bílek, L.; Remeš, J.; Štefančík, I. Changes in horizontal structure of natural beech forests on an altitudinal gradient in the Sudetes. Dendrobiology 2015, 73, 33–45. [Google Scholar] [CrossRef]
- Vacek, Z.; Vacek, S.; Bulušek, D.; Podrázský, V.; Remeš, J.; Král, J.; Putalová, T. Effect of fungial pathogenes and climatic factors on production, biodiversity and health status of ash in mountain forests. Dendrobiology 2017, 77, 161–175. [Google Scholar] [CrossRef]
- O’Callaghan, D.P.; Atkins, P.M.; Fairhurst, C.P. Behavioral responses of elm bark beetles to baited and unbaited elms killed by cacodylic acid. J. Chem. Ecol. 1984, 10, 1623–1634. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, J.N.; Brasier, C.M.; Webber, J.F. Dutch Elm Disease in Britain. In Forestry Commission Information Note 252; Forestry Commission Edinbourgh: Edinbourgh, UK, 1994. [Google Scholar]
- Mina, M.; del Río, M.; Huber, M.O.; Thürig, E.; Rohner, B. The symmetry of competitive interactions in mixed Norway spruce, silver fir and European beech forests. J. Veg. Sci. 2018, 29, 775–787. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef]
- Jandl, R. Climate-induced challenges of Norway spruce in Northern Austria. Trees For. People 2020, 1, 100008. [Google Scholar] [CrossRef]
- Nothdurft, A.; Engel, M. Climate sensitivity and resistance under pure– and mixed-stand scenarios in Lower Austria evaluated with distributed lag models and penalized regression splines for tree-ring time series. Eur. J. For. Res. 2020, 139, 189–211. [Google Scholar] [CrossRef]
- Forrester, D.I.; Kohnle, U.; Albrecht, A.T.; Bauhus, J. Complementarity in mixed-species stands of Abies alba and Picea abies varies with climate, site quality and stand density. For. Ecol. Manag. 2013, 304, 233–242. [Google Scholar] [CrossRef]
- Block, J. Disposition rheinland-pfälzischer Waldbodensubstrate gegenüber Versauerung; Forest Damage, Soil and Water Conservation through Airborne Substances in Rhineland-Palatinate; Ministry of the Environment and Forest of Rhineland-Palatinate: Mainz, Germany, 1997.
- Augusto, L.; Ranger, J.; Binkley, D.; Rothe, A. Impact of several common tree species of European temperate forests on soil fertility. Ann. For. Sci. 2002, 59, 233–253. [Google Scholar] [CrossRef]
- Forrester, D.I.; Albrecht, A.T. Light absorption and light-use efficiency in mixtures of Abies alba and Picea abies along a productivity gradient. For. Ecol. Manag. 2014, 328, 94–102. [Google Scholar] [CrossRef]
- Huber, M.O.; Sterba, H.; Bernhard, L. Site conditions and definition of compositional proportion modify mixture effects in Picea abies—Abies alba stands. Can. J. For. Res. 2014, 44, 1281–1291. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.J.; Nabuurs, G.J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Konôpková, A.; Kurjak, D.; Kmeť, J.; Klumpp, R.; Longauer, R.; Ditmarová, Ľ.; Gömöry, D. Differences in photochemistry and response to heat stress between silver fir (Abies alba Mill.) provenances. Trees Struct. Funct. 2018, 32, 73–86. [Google Scholar] [CrossRef]
- Zimmermann, J.; Hauck, M.; Dulamsuren, C.; Leuschner, C. Climate Warming-Related Growth Decline Affects Fagus sylvatica, But Not Other Broad-Leaved Tree Species in Central European Mixed Forests. Ecosystems 2015, 18, 560–572. [Google Scholar] [CrossRef]
- Lebourgeois, F.; Rathgeber, C.B.K.; Ulrich, E. Sensitivity of French temperate coniferous forests to climate variability and extreme events (Abies alba, Picea abies and Pinus sylvestris). J. Veg. Sci. 2010, 21, 364–376. [Google Scholar] [CrossRef]
- Cavlovic, J.; Boncina, A.; Bozic, M.; Goršic, E.; Simončič, T.; Teslak, K. Depression and growth recovery of silver fir in uneven-aged Dinaric forests in Croatia from 1901 to 2001. Forestry 2014, 88, 586–598. [Google Scholar] [CrossRef]
- Carrer, M.; Motta, R.; Nola, P. Significant Mean and Extreme Climate Sensitivity of Norway Spruce and Silver Fir at Mid-Elevation Mesic Sites in the Alps. PLoS ONE 2012, 7, e50755. [Google Scholar] [CrossRef]
- Rötzer, T.; Biber, P.; Moser, A.; Schäfer, C.; Pretzsch, H. Stem and root diameter growth of European beech and Norway spruce under extreme drought. For. Ecol. Manag. 2017, 406, 184–195. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter-specific facilitation. Plant Biol. 2013, 15, 438–495. [Google Scholar] [CrossRef]
- Spinoni, J.; Naumann, G.; Vogt, J.V.; Barbosa, P. The biggest drought events in Europe from 1950 to 2012. J. Hydrol. Reg. Stud. 2015, 3, 509–524. [Google Scholar] [CrossRef]
- Ols, C.; Bontemps, J.-D.; Gschwantner, T. Nadelbäume—Wie reagiert das Wachstum auf die Klimaerwärmung. BFW Prax. 2019, 50, 30–34. [Google Scholar]
- Vitali, V.; Buntegen, U.; Bauhus, J. Seasonality matters—The effects of past and projected seasonal climate change on the growth of native and exotic conifer species in Central Europe. Dendrochronologia 2018, 48, 1–9. [Google Scholar] [CrossRef]
- Gruber, A.; Zimmermann, J.; Wieser, G.; Oberhuber, W. Effects of climate variables on intra-annual stem radial increment in Pinus cembra (L.) along the alpine treeline ecotone. Ann. For. Sci. 2009, 66, 503. [Google Scholar] [CrossRef] [PubMed]
- Swidrak, I.; Gruber, A.; Kofler, W.; Oberhuber, W. Effects of environmental conditions on onset of xylem growth in Pinus sylvestris under drought. Tree Physiol. 2011, 31, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Hänninen, H.; Tanino, K. Tree seasonality in a warming climate. Trends Plant Sci. 2011, 16, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Oberhuber, W.; Gruber, A.; Kofler, W.; Swidrak, I. Radial stem growth in response to microclimate and soil moisture in a drought-prone mixed coniferous forest at an inner Alpine site. Eur. J. For. Res. 2014, 133, 467–479. [Google Scholar] [CrossRef]
- Ciceu, A.; Popa, I.; Leca, S.; Pitar, D.; Chivulescu, S.; Badea, O. Climate change effects on tree growth from Romanian forest monitoring Level II plots. Sci. Total Environ. 2020, 698, 134129. [Google Scholar] [CrossRef]
- Versace, S.; Gianelle, D.; Garfì, V.; Battipaglia, G.; Lombardi, F.; Marchetti, M.; Tognetti, R. Interannual radial growth sensitivity to climatic variations and extreme events in mixed-species and pure forest stands of silver fir and European beech in the Italian Peninsula. Eur. J. For. Res. 2020, 139, 1–19. [Google Scholar] [CrossRef]
- Klein, T. The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours. Funct. Ecol. 2014, 28, 1313–1320. [Google Scholar] [CrossRef]
- Roman, D.T.; Novick, K.A.; Brzostek, E.R.; Dragoni, D.; Rahman, F.; Phillips, R.P. The role of isohydric and anisohydric species in determining ecosystem-scale response to severe drought. Oecologia 2015, 179, 641–654. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.G.; Allen, C.D. Darcy’s law predicts widespread forest mortality under climate warming. Nat. Clim. Chang. 2015, 5, 669–672. [Google Scholar] [CrossRef]
- Kolb, T.E.; Stone, J.E. Differences in leaf gas exchange and water relations among species and tree sizes in an Arizona pine-oak forest. Tree Physiol. 2000, 20, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Barbeta, A.; Ogaya, R.; Peñuelas, J. Dampening effects of long-term experimental drought on growth and mortality rates of a Holm oak forest. Glob. Chang. Biol. 2013, 19, 3133–3144. [Google Scholar] [CrossRef]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A.; Steppe, K. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef]
- Hoffmann, N.; Schall, P.; Ammer, C.; Leder, B.; Vor, T. Drought sensitivity and stem growth variation of nine alien and native tree species on a productive forest site in Germany. Agric. For. Meteorol. 2018, 256–257, 431–444. [Google Scholar] [CrossRef]
- O’Brien, M.J.; Leuzinger, S.; Philipson, C.D.; Tay, J.; Hector, A. Drought survival of tropical tree seedlings enhanced by non-structural carbohydrate levels. Nat. Clim. Chang. 2014, 4, 710–714. [Google Scholar] [CrossRef]
- Pretzsch, H.; Rötzer, T.; Matyssek, R.; Grams, T.E.E.; Häberle, K.H.; Pritsch, K.; Kerner, R.; Munch, J.C. Mixed Norway spruce (Picea abies [L.] Karst) and European beech (Fagus sylvatica [L.]) stands under drought: From reaction pattern to mechanism. Trees Struct. Funct. 2014, 28, 1305–1321. [Google Scholar] [CrossRef]
- Rötzer, T.; Häberle, K.H.; Kallenbach, C.; Matyssek, R.; Schütze, G.; Pretzsch, H. Tree species and size drive water consumption of beech/spruce forests—A simulation study highlighting growth under water limitation. Plant Soil 2017, 418, 337–356. [Google Scholar] [CrossRef]
- Dănescu, A.; Albrecht, A.T.; Bauhus, J. Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany. Oecologia 2016, 182, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Gričar, J.; Čufar, K. Seasonal dynamics of phloem and xylem formation in silver fir and Norway spruce as affected by drought. Russ. J. Plant Physiol. 2008, 55, 538–543. [Google Scholar] [CrossRef]
- Swidrak, I.; Gruber, A.; Oberhuber, W. Xylem and phloem phenology in co-occurring conifers exposed to drought. Trees Struct. Funct. 2014, 28, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Harrington, C.A.; Gould, P.J.; St.Clair, J.B. Modeling the effects of winter environment on dormancy release of Dougla-fir. For. Ecol. Manag. 2010, 259, 798–808. [Google Scholar] [CrossRef]
- Rolland, C.; Michalet, R.; Desplanque, C.; Petetin, A.; Aimé, S. Ecological requirements of Abies alba in the French Alps derived from dendro-ecological analysisu. J. Veg. Sci. 1999, 10, 297–306. [Google Scholar] [CrossRef]
- Mäkinen, H.; Nöjd, P.; Saranpää, P. Seasonal changes in stem radius and production of new tracheids in Norway spruce. Tree Physiol. 2003, 23, 959–968. [Google Scholar] [CrossRef] [PubMed]
- van der Maaten, E. Climate sensitivity of radial growth in European beech (Fagus sylvatica L.) at different aspects in southwestern Germany. Trees 2012, 26, 777–788. [Google Scholar] [CrossRef]
- Hathaway, D.H. The solar cycle. Living Rev. Sol. Phys. 2015, 12, 4. [Google Scholar] [CrossRef]
- Šimůnek, V.; Vacek, Z.; Vacek, S.; Ripullone, F.; Hájek, V.; D’andrea, G. Tree rings of european beech (Fagus sylvatica L.) indicate the relationship with solar cycles during climate change in central and southern europe. Forests 2021, 12, 259. [Google Scholar] [CrossRef]
- Kasatkina, E.A.; Shumilov, O.I.; Timonen, M. Solar activity imprints in tree ring-data from northwestern Russia. J. Atmos. Sol. Terr. Phys. 2019, 193, 105075. [Google Scholar] [CrossRef]
- Matveev, S.M.; Chendev, Y.G.; Lupo, A.R.; Hubbart, J.A.; Timashchuk, D.A. Climatic Changes in the East-European Forest-Steppe and Effects on Scots Pine Productivity. Pure Appl. Geophys. 2017, 174, 427–443. [Google Scholar] [CrossRef]
- McCracken, K.G.; Beer, J.; McDonald, F.B. A five-year variability in the modulation of the galactic cosmic radiation over epochs of low solar activity. Geophys. Res. Lett. 2002, 29, 3–6. [Google Scholar] [CrossRef]
- Carl, G.; Doktor, D.; Koslowsky, D. Phase difference analysis of temperature and vegetation phenology for beech forest: A wavelet approach. Stoch. Environ. Res. Risk Assess. 2013, 5, 1221–1230. [Google Scholar] [CrossRef]
- Tsiropoula, G. Signatures of solar activity variability in meteorological parameters. J. Atmos. Sol. Terr. Phys. 2003, 65, 469–482. [Google Scholar] [CrossRef]
- Laurenz, L.; Lüdecke, H.J.; Lüning, S. Influence of solar activity changes on European rainfall. J. Atmos. Sol. Terr. Phys. 2019, 185, 29–42. [Google Scholar] [CrossRef]
- Shumilov, O.I.; Kasatkina, E.A.; Mielikainen, K.; Timonen, M.; Kanatjev, A.G. Palaeovolcanos, solar activity and pine tree-rings from the Kola Peninsula (northwestern Russia) over the last 560 years. Int. J. Environ. Res. 2011, 5, 855–864. [Google Scholar] [CrossRef]
- Dorotovič, I.; Louzada, J.L.; Rodrigues, J.C.; Karlovský, V. Impact of solar activity on the growth of pine trees: Case study. Eur. J. For. Res. 2014, 133, 639–648. [Google Scholar] [CrossRef]
- Nagovitsyn, Y.A. Specific features in the effect of solar activity on the Earth’s climate changes. Geomagn. Aeron. 2014, 54, 1010–1013. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PRP | GPS | Altitude (m) | Expo- Sition | Slope (°) | Forest Site Type | Geology | Soils | Species | Age of Tree Layers (Years) | Stand Volume (m3 ha−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 50°44′57″ N, 15°24′43″ E | 600 | SW | 15 | Acereto-Fagetum lapidosum | phyllite | cambic Rankers | Pa, Fs, Ap, Ug | 151/18 | 336 |
| 2 | 50°44′59″ N, 15°24′37″ E | 590 | SW | 17 | Acereto-Fagetum lapidosum | phyllite | cambic Rankers | Pa, Fs, Ap, Ug, Aa | 121/18 | 489 |
| 3 | 50°45′03″ N, 15°24′40″ E | 620 | SW | 13 | Acereto-Fagetum lapidosum | phyllite | ranker Cambisols | Pa, Fs, Ap, Aa | 107/17 | 394 |
| 4 | 50°45′03″ N, 15°24′42″ E | 610 | SW | 15 | Acereto-Fagetum lapidosum | phyllite | modal Rankers | Pa, Fs, Ap, Fe | 105/7 | 377 |
| 5 | 50°45′37″ N, 15°24′40″ E | 680 | SW | 13 | Piceeto-Fagetum lapidosum acidophilum | quartzite | cambic Rankers | Pa, Fs, Ap | 128/39/19 | 419 |
| 6 | 50°45′40″ N, 15°24′34″ E | 700 | SW | 32 | Piceeto-Fagetum saxatile | quartzite | modal Rankers | Pa, Fs, Ap, Fe | 130/24 | 484 |
| 7 | 50°45′38″ N, 15°24′25″ E | 650 | W | 15 | Piceeto-Fagetum lapidosum acidophilum | quartzite | ranker Cambisols | Pa, Fs, Ap, Ug | 136/19 | 359 |
| Criterion | Quantifiers | Label | Reference | Evaluation |
|---|---|---|---|---|
| Species diversity | Richness | D (Mi) | [70] | minimum D = 0, higher D = higher values |
| Heterogeneity | H’ (Si) | [71] | minimum H’ = 0, higher H’ = higher values | |
| Evenness | E (Pii) | [72] | range 0–1; minimum E = 0, maximum E = 1 | |
| Horizontal structure | Aggregation index | R (C&Ei) | [73] | mean value R = 1; aggregation R < 1; regularity R > 1 |
| Vertical structure | Arten-profile index | A (Pri) | [74] | range 0–1; balanced vertical structure A < 0.3; selection forest A > 0.9 |
| Vertical div. | S (J&Di) | [75] | low S < 0.3, medium S = 0.3–0.5, high S = 0.5–0.7, very high diversity S > 0.7 | |
| Structure differentiation | Diameter dif. | TMd (Fi) | [76] | range 0–1; low TM < 0.3; very high differentiation TM > 0.7 |
| Height dif. | TMh (Fi) | |||
| Crown dif. | K (J&Di) | [75] | low K < 1.0, medium K = 1.0–1.5, high K = 1.5–2.0, very high differentiation K > 2.0 | |
| Complex diversity | Stand diversity | B (J&Di) | [75] | monotonous structure B < 4; uneven structure B = 6–8; very diverse structure B > 9 |
| PRP | Age | DBH | h | f | v | N | BA | V | HDR | MAI | CC | CPA | SDI |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (yr) | (cm) | (m) | (m3) | (trees ha−1) | (m2 ha−1) | (m3 ha−1) | (m3 ha−1 yr−1) | (%) | (ha ha−1) | ||||
| 1 | 151 | 43.7 | 32.3 | 0.516 | 2.504 | 276 | 41.4 | 691 | 74.2 | 4.58 | 84.0 | 3.18 | 0.63 |
| 2 | 121 | 39.7 | 28.1 | 0.498 | 1.860 | 344 | 42.6 | 640 | 71.8 | 5.29 | 88.9 | 3.43 | 0.68 |
| 3 | 107 | 41.5 | 24.8 | 0.524 | 1.755 | 340 | 45.8 | 597 | 60.4 | 5.58 | 76.6 | 2.16 | 0.60 |
| 4 | 105 | 36.0 | 24.3 | 0.541 | 1.338 | 480 | 48.9 | 642 | 62.6 | 4.57 | 97.4 | 2.51 | 0.80 |
| 5 | 128 | 46.0 | 33.1 | 0.464 | 2.554 | 304 | 50.5 | 776 | 72.0 | 6.06 | 92.1 | 3.31 | 0.70 |
| 6 | 130 | 43.0 | 21.8 | 0.594 | 1.881 | 296 | 42.8 | 557 | 66.1 | 4.28 | 95.6 | 2.23 | 0.66 |
| 7 | 136 | 49.6 | 33.2 | 0.488 | 3.132 | 260 | 50.1 | 814 | 71.3 | 5.99 | 94.0 | 3.12 | 0.71 |
| PRP | D (Mi) | H’ (Si) | E (Pii) | R (C&Ei) | A (Pri) | S (J&Di) | TMd (Fi) | TMh (Fi) | K (J&Di) | B (J&Di) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 0.356 | ↘ | 0.332 | ↘ | 0.696 | → | 0.806 * | A | 0.457 | ↘ | 0.780 | ↗ | 0.372 | ↘ | 0.208 | ↘↘ | 1.620 | → | 6.152 | → |
| 2 | 0.392 | ↘ | 0.441 | ↘ | 0.631 | → | 0.920 | 0.474 | ↘ | 0.621 | → | 0.407 | ↘ | 0.231 | ↘↘ | 1.194 | ↘ | 6.954 | → | |
| 3 | 0.172 | ↘↘ | 0.120 | ↘↘ | 0.399 | ↘ | 1.036 | 0.314 | ↘ | 0.848 | ↗ | 0.454 | ↘ | 0.378 | ↘ | 2.998 | → | 6.073 | → | |
| 4 | 0.486 | ↘ | 0.426 | ↘ | 0.708 | ↗ | 0.982 | 0.682 | → | 0.864 | ↗ | 0.442 | ↘ | 0.320 | ↘ | 1.684 | → | 6.848 | → | |
| 5 | 0.175 | ↘↘ | 0.295 | ↘ | 0.980 | ↗↗ | 0.964 | 0.655 | → | 0.842 | ↗ | 0.403 | ↘ | 0.229 | ↘↘ | 1.756 | → | 6.845 | → | |
| 6 | 0.351 | ↘ | 0.424 | ↘ | 0.889 | ↗ | 1.082 | 0.732 | ↗ | 0.894 | ↗ | 0.461 | ↘ | 0.358 | ↘ | 1.756 | → | 6.845 | → | |
| 7 | 0.360 | ↘ | 0.321 | ↘ | 0.673 | → | 0.868 * | A | 0.571 | → | 0.782 | ↗ | 0.330 | ↘ | 0.157 | ↘↘ | 1.576 | → | 6.268 | → |
| Species | No. (Cores) | Age Min.–Max. (yr) | Mean RW (mm) | Min.–Max. RW (mm) | SD | Ar1 | R-Bar | EPS |
|---|---|---|---|---|---|---|---|---|
| Norway spruce | 27 | 85–135 | 2.41 | 1.774–3.354 | 1.27 | 0.76 | 0.46 | 0.95 |
| European beech | 30 | 104–152 | 1.43 | 1.065–2.463 | 0.70 | 0.68 | 0.27 | 0.89 |
| Mountain elm | 26 | 84–145 | 1.98 | 1.124–3.264 | 1.28 | 0.85 | 0.24 | 0.86 |
| European ash | 28 | 84–120 | 2.09 | 1.497–3.623 | 1.11 | 0.80 | 0.26 | 0.87 |
| Sycamore maple | 24 | 80–134 | 1.96 | 1.173–2.530 | 1.17 | 0.82 | 0.19 | 0.65 |
| RWI | PreAct Ann | PreLas Ann | PreAct Veg | PreAct VI-VII | PreAct NonVeg | PreAct VarSD | TemAct Ann | TemLas Ann | TemAct Veg |
|---|---|---|---|---|---|---|---|---|---|
| Beech | −0.07 | −0.12 | −0.07 | −0.27 * | 0.10 | −0.05 | 0.21 | 0.38 ** | 0.12 |
| Elm | −0.05 | 0.19 | 0.11 | 0.03 | −0.12 | −0.21 | −0.14 | −0.09 | −0.03 |
| Ash | −0.08 | −0.05 | −0.03 | −0.10 | 0.02 | −0.30 * | −0.02 | −0.02 | 0.17 |
| Maple | 0.03 | 0.11 | 0.07 | −0.09 | −0.02 | −0.09 | 0.05 | −0.03 | 0.08 |
| Spruce | 0.04 | 0.14 | 0.12 | 0.14 | −0.10 | −0.15 | 0.02 | 0.05 | 0.01 |
| RWI | TemAct VI-VII | TemAct NonVeg | TemAct VarSD | SO2 Mean | SO2 Max | NOX Mean | NOX Max | AOT40F Mean | AOT40F Max |
| Beech | 0.17 | 0.33 ** | −0.13 | −0.19 | −0.18 | −0.26 | −0.43 * | −0.39 * | −0.39 * |
| Elm | 0.07 | −0.26 * | 0.28 * | −0.11 | −0.10 | −0.07 | −0.03 | −0.16 | −0.22 |
| Ash | 0.26 * | −0.09 | 0.38 ** | −0.18 | 0.04 | −0.02 | −0.05 | 0.21 | −0.03 |
| Maple | 0.09 | 0.11 | 0.16 | −0.10 | −0.11 | −0.01 | −0.05 | −0.20 | −0.35 |
| Spruce | −0.08 | 0.03 | −0.03 | −0.43 ** | −0.37 ** | −0.31 | −0.43 * | −0.59 ** | −0.59 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hájek, V.; Vacek, S.; Vacek, Z.; Cukor, J.; Šimůnek, V.; Šimková, M.; Prokůpková, A.; Králíček, I.; Bulušek, D. Effect of Climate Change on the Growth of Endangered Scree Forests in Krkonoše National Park (Czech Republic). Forests 2021, 12, 1127. https://doi.org/10.3390/f12081127
Hájek V, Vacek S, Vacek Z, Cukor J, Šimůnek V, Šimková M, Prokůpková A, Králíček I, Bulušek D. Effect of Climate Change on the Growth of Endangered Scree Forests in Krkonoše National Park (Czech Republic). Forests. 2021; 12(8):1127. https://doi.org/10.3390/f12081127
Chicago/Turabian StyleHájek, Vojtěch, Stanislav Vacek, Zdeněk Vacek, Jan Cukor, Václav Šimůnek, Michaela Šimková, Anna Prokůpková, Ivo Králíček, and Daniel Bulušek. 2021. "Effect of Climate Change on the Growth of Endangered Scree Forests in Krkonoše National Park (Czech Republic)" Forests 12, no. 8: 1127. https://doi.org/10.3390/f12081127
APA StyleHájek, V., Vacek, S., Vacek, Z., Cukor, J., Šimůnek, V., Šimková, M., Prokůpková, A., Králíček, I., & Bulušek, D. (2021). Effect of Climate Change on the Growth of Endangered Scree Forests in Krkonoše National Park (Czech Republic). Forests, 12(8), 1127. https://doi.org/10.3390/f12081127