
Invertebrate Assemblages on Biscogniauxia Sporocarps on Oak Dead Wood: An Observation Aided by Squirrels


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Data Collection
2.3. Data Analysis
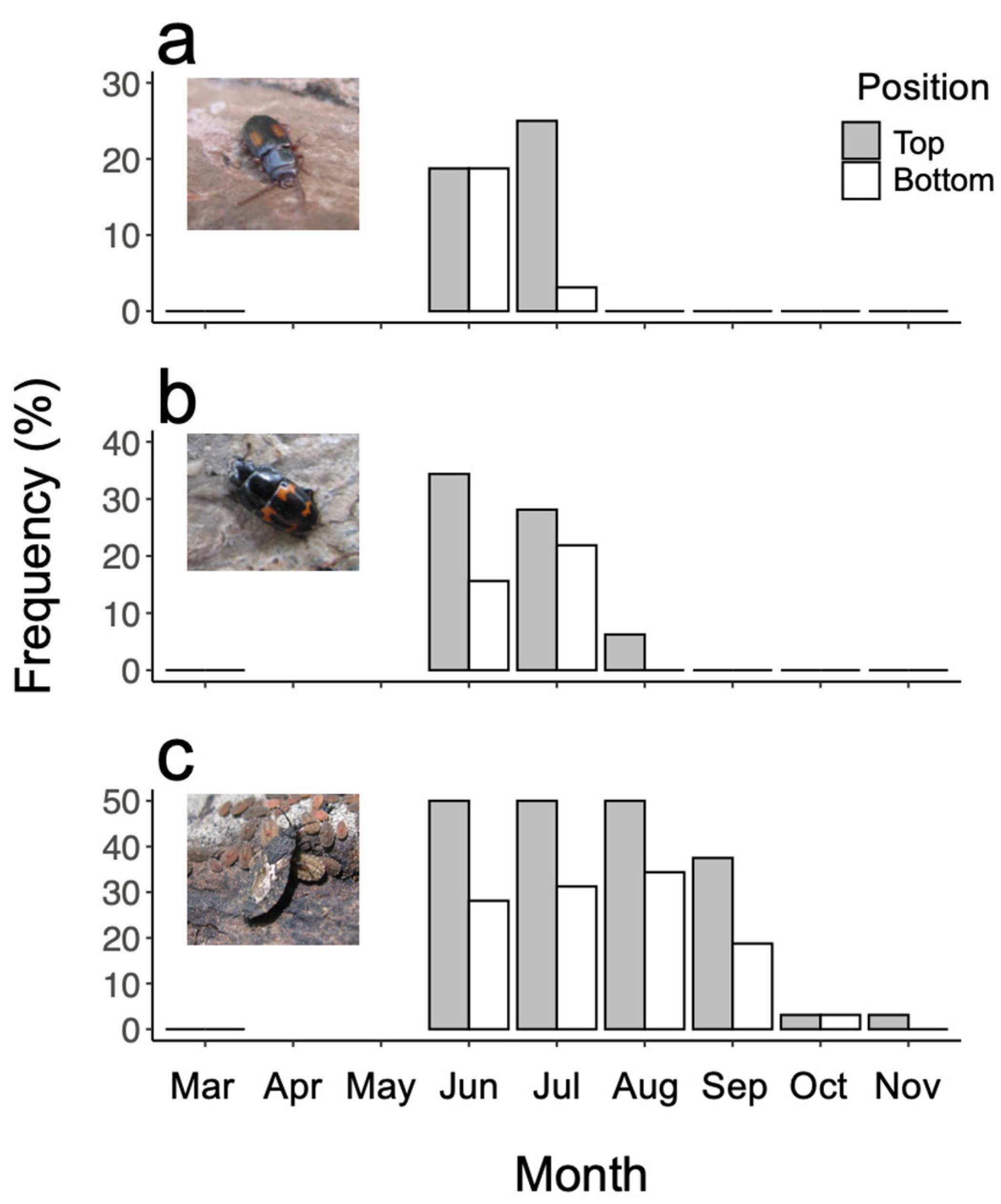
3. Results
4. Discussion
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Stokland, J.N.; Siitonen, J.; Jonsson, B.G. Biodiversity in Dead Wood; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Seibold, S.; Bässler, C.; Brandl, R.; Gossner, M.M.; Thorn, S.; Ulyshen, M.D.; Müller, J. Experimental studies of dead-wood biodiversity—A review identifying global gaps in knowledge. Biol. Conserv. 2015, 191, 139–149. [Google Scholar] [CrossRef]
- Rayner, A.D.M.; Boddy, L. Fungal Decomposition of Wood: Its Biology and Ecology; John Wiley & Sons: Chichester, UK, 1988. [Google Scholar]
- Fukasawa, Y.; Hyodo, F.; Kawakami, S. Foraging association between myxomycetes and fungal communities on coarse woody debris. Soil Biol. Biochem. 2018, 121, 95–102. [Google Scholar] [CrossRef]
- Hanski, I. Fungivory: Fungi, insects and Ecology. In Insect–Fungus Interactions; Wilding, N., Collins, N.M., Hammond, P.M., Webber, J.F., Eds.; Academic Press: London, UK, 1989. [Google Scholar]
- Currah, R.S.; Smreciu, E.A.; Lehesvirta, T.; Niemi, M.; Larsen, K.W. Fungi in the winter diets of northern flying squirrels and red squirrels in the boreal mixedwood forest of northern Alberta. Can. J. Bot. 2000, 78, 1514–1520. [Google Scholar]
- Elliott, T.F.; Elliott, K. Vertebrate consumption and dispersal of the Nothofagaceae associated ascomycete Cyttaria. Austral. Ecol. 2019, 44, 747–751. [Google Scholar] [CrossRef]
- Schigel, D.S. Fungivory and host associations of Coleoptera: A bibliography and review of research approaches. Mycology 2012, 3, 258–272. [Google Scholar] [CrossRef]
- Buxton, C.M.G. British Diptera associated with fungi. III. Flies of all families reared from about 150 species of fungi. Entomol. Mon. Mag. 1960, 96, 61–94. [Google Scholar]
- Elton, C.S. The Pattern of Animal Communities; Methuen and Co.: Slingsby, UK, 1966. [Google Scholar]
- Jonsell, M.; Nordlander, G. Host selection patterns in insects breeding in bracket fungi. Ecol. Entomol. 2004, 29, 697–705. [Google Scholar] [CrossRef]
- Yamashita, S.; Hijii, N. The role of fungal taxa and developmental stage of mushrooms in determining the composition of the mycophagous insect community in a Japanese forest. Eur. J. Entomol. 2007, 104, 225–233. [Google Scholar] [CrossRef]
- Nakamori, T.; Suzuki, A. Surface properties of the mushroom Strobilurus ohshimae result in food differentiation by collembolan species. Eur. J. Soil Biol 2008, 44, 478–482. [Google Scholar] [CrossRef]
- Yamashita, S.; Ando, K.; Hoshina, H.; Ito, N.; Katayama, Y.; Kawanabe, M.; Maruyama, M.; Itioka, T. Food web structure of the fungivorous insect community on bracket fungi in a Bornean tropical rain forest. Ecol. Entomol. 2015, 40, 390–400. [Google Scholar] [CrossRef]
- Kobayashi, T.; Sota, T. Evolution of host use in fungivorous ciid beetles (Coleoptera: Ciidae): Molecular phylogeny focusing on Japanese taxa. Mol. Phylogen Evol. 2021, 162, 107197. [Google Scholar] [CrossRef] [PubMed]
- Tuno, N. Insect feeding on spores of a bracked fungus, Elfvingia applanata (Pers.) Karst. (Ganodermataceae, Aphyllophorales). Ecol. Res. 1999, 14, 97–103. [Google Scholar] [CrossRef]
- Kadowaki, K.; Keschen, R.A.B.; Beggs, J.R. No evidence for a Ganoderma spore dispersal mutualism in an obligate spore-feeding beetle Zearagytodes maculifer. Fung Biol. 2011, 115, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Persson, Y.; Ihrmark, K.; Stenlid, J. Do bark beetles facilitate the establishment of rot fungi in Norway spruce? Fung. Ecol. 2011, 4, 262–269. [Google Scholar] [CrossRef]
- Kitabayashi, K.; Tuno, N. Soil burrowing Muscina angustifrons (Diptera: Muscidae) larvae excrete spores capable of forming mycorrhizae underground. Mycoscience 2018, 59, 252–258. [Google Scholar] [CrossRef]
- Seibold, S.; Müller, J.; Baldrian, P.; Cadotte, M.W.; Stursová, M.; Biedermann, P.H.W.; Krah, F.S.; Bässler, C. Fungi associated with beetles dispersing from dead wood—Let’s take the beetle bus! Fung. Ecol. 2019, 39, 100–108. [Google Scholar] [CrossRef]
- Takahashi, K.H.; Tuno, N.; Kagaya, T. The relative importance of spatial aggregation and resource partitioning on the coexistence of mycophagous insects. Oikos 2005, 109, 125–134. [Google Scholar] [CrossRef]
- Kadowaki, K. Species coexistence patterns in a mycophagous insect community inhabiting the wood-decaying bracket fungus Cryptoporus volvatus. Eur. J. Entomol. 2010, 107, 89–99. [Google Scholar] [CrossRef]
- Komonen, A.; Penttila, R.; Lindgren, M.; Hanski, I. Forest fragmentation truncates a food chain based on an old-growth forest bracket fungus. Oikos 2000, 90, 119–126. [Google Scholar] [CrossRef]
- Komonen, A.; Kouki, J. Occurrence and abundance of fungus-dwelling beetles (Ciidae) in boreal forests and clearcuts: Habitat associations at two spatial scales. Anim. Biodiv. Conserv. 2005, 28, 137–147. [Google Scholar]
- Joseph, R.; Keyhani, N.O. Fungal mutualisms and pathosystems: Life and death in the ambrosia beetle mycangia. Appl. Microbiol. Biotechnol. 2021, 105, 3393–3410. [Google Scholar] [CrossRef]
- Pažoutová, S.; Srutka, P.; Holusa, J.; Chudickova, M.; Kolarik, M. Diversity of xylariaceous symbionts in Xiphydria woodwasps: Role of vector and a host tree. Fung. Ecol. 2010, 3, 392–401. [Google Scholar] [CrossRef]
- Visser, A.A.; Ros, V.I.D.; de Beer, Z.W.; Debets, A.J.M.; Hartog, E.; Kuyper, T.W.; Læssøe, T.; Slippers, B.; Aanen, D.K. Level of specificity of Xylaria species associated with fungus-growing termites: A phylogenetic approach. Mol. Ecol. 2009, 18, 553–567. [Google Scholar] [CrossRef]
- Xiao, G.; Ying, S.H.; Zheng, P.; Wang, Z.L.; Zhang, S.; Xie, X.Q.; Shang, Y.; St Leger, R.J.; Zhao, G.P.; Wang, C.; et al. Genomic perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [Google Scholar] [CrossRef] [PubMed]
- Stone, L.B.L.; Bidochka, M.J. The multifunctional lifestyles of Metarhizium: Evolution and applications. Appl. Microbiol. Biotechnol. 2020, 104, 9935–9945. [Google Scholar] [CrossRef] [PubMed]
- Nikoh, N.; Fukatsu, T. Interkingdom host jumping underground: Phylogenetic analysis of entomoparasitic fungi of the genus Cordyceps. Mol. Biol. Evol. 2000, 17, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Okada, T.; Tomaru, N.; Nishimura, N.; Nakagawa, M. Forest dynamics of stands damaged by Japanese oak wilt in the Kaisho forest, Aichi Prefecture. J. Jap. For. Soc. 2016, 98, 273–278. [Google Scholar] [CrossRef]
- Nishikawa, S.; Kubo, M.; Ozaki, Y. Mass mortality due to the spread of oak wilt disease in a 19-ha secondary Quercus serrata forest within Sambe Forest at Shimane University. Bull Fac. Life Env. Sci. Shimane Univ. 2018, 23, 21–26. [Google Scholar]
- Endoh, R.; Suzuki, M.; Okada, G.; Takeuchi, Y.; Futai, K. Fungus symbionts colonizing the galleries of the ambrosia beetle Platypus quercivorus. Microb. Ecol. 2011, 62, 106–120. [Google Scholar] [CrossRef]
- Masuya, H.; Manabe, R.; Ohkuma, M.; Endoh, R. Draft genome sequence of Raffaelea quercivora JCM 11526, a Japanese oak wilt pathogen associated with the Platypodid beetle, Platypus quecivorus. Genome Announc. 2016, 4, e00755-16. [Google Scholar] [CrossRef]
- Kobayashi, T.; Oguro, M.; Akiba, M.; Taki, H.; Kitajima, H.; Ishihara, H. Mushroom yield of cultivated shiitake (Lentinula edodes) and fungal communities in logs. J. Res. 2020, 25, 269–275. [Google Scholar] [CrossRef]
- Tokimoto, K.; Komatsu, M.; Fukumasa-Nakai, Y. Establishing the selection method for the Lentinula edodes strains resistant to Trichoderma spp. Proc. Jpn. Acad 1994, 70, 112–116. [Google Scholar] [CrossRef][Green Version]
- Kim, J.Y.; Kwon, H.W.; Yun, Y.H.; Kim, S.H. Identification and characterization of Trichoderma species damaging Shiitake mushroom bed-logs infested by Camptomyia pest. J. Microbiol Biotechnol 2016, 26, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Kurosawa, Y.; Hisamatsu, S.; Sasaji, H. The coleoptera of Japan in color Vol.III; Hoikusha: Tokyo, Japan, 1998. (In Japanese) [Google Scholar]
- Ueno, S.; Kurosawa, Y.; Sato, M. The coleoptera of Japan in color Vol.II; Hoikusha: Tokyo, Japan, 1999. (In Japanese) [Google Scholar]
- Hayashi, M.; Morimoto, K.; Kimoto, S. The coleoptera of Japan in color Vol.IV; Hoikusha: Tokyo, Japan, 2002. (In Japanese) [Google Scholar]
- Ishikawa, T.; Takai, M.; Yasunaga, T. A Field Guide to Japanese Bugs: Terrestrial Heteropterans; Zenkoku Noson Kyoiku Kyokai: Tokyo, Japan, 2012; Volume 3. (In Japanese) [Google Scholar]
- Terayama, M.; Kubota, S.; Eguchi, K. Encyclopedia of Japanese Ants; Asakura Shoten: Tokyo, Japan, 2014. (In Japanese) [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; The R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package; R Package Version 2.5-7; 2020; Available online: https://cran.r-project.org/web/packages/vegan/index.html.
- Anderson, M.J. A new method for non parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Ulyshen, M.D. Saproxylic Insects: Diversity, Ecology and Conservation; Springer: Cham, Switzerland, 2019. [Google Scholar]
- McKeever, S. The biology of the Golden-mantled ground squirrel, Citellus lateralis. Ecol. Monogr. 1964, 34, 383–401. [Google Scholar] [CrossRef]
- Maser, C.; Maser, Z. Interactions among squirrels, mycorrhizal fungi, and coniferous forests in Oregon. Great Basin Nat. 1988, 48, 358–369. [Google Scholar]
- Stadler, M. Importance of secondary metabolites in the Xylariaceae as parameters for assessment of their taxonomy, phylogeny, and functional biodiversity. Curr. Res. Environ. Appl. Mycol. 2011, 1, 75–133. [Google Scholar] [CrossRef]
- Cork, S.J.; Kenagy, G.J. Nutritional value of hypogeous fungus for a forest-dwelling ground squirrel. Ecology 1989, 70, 577–586. [Google Scholar] [CrossRef]
- Crowson, R.A. The associations of Coleoptera with Ascomycetes. In Fungus-Insect Relationships; Wheeler, Q., Blackwell, M., Eds.; Columbia University Press: New York, NY, USA, 1984; pp. 257–285. [Google Scholar]
- Katanoda, I.; Sato, Y.; Morita, S.; Okubo, H. Adult emergence and oviposition preference of Mesosa longipennis for Chinese Quercus acutissima. Kyushu J. Res. 2005, 58, 105–108. (In Japanese) [Google Scholar]
- Gossner, M.M.; Damken, C. Diversity and ecology of saproxylic Hemiptera. In Saproxylic Insects: Diversity, Ecology and Conservation (Zoological Monographs 1); Ulyshen, M.D., Ed.; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Powell, J.A.; Mitter, C.; Farrell, B. Evolution of larval food preferences in Lepidoptera. In Lepidoptera, Moths and Butterflies Volume 1: Evolution, Systematics, and Biogeography; Kristensen, N.P., Ed.; de Gruyter: Berlin, Germany, 1998; pp. 403–422. [Google Scholar]
- Powell, J.A. Apomyelois bistriatella: A moth which feeds in an ascomycete fungus (Lepidoptera: Pyralidae). J. N. Y. Entomol. Soc. 1967, 75, 190–194. [Google Scholar]
- Lee, Y.-B.; An, S.J.; Park, C.G.; Kim, J.; Han, S.; Kwak, Y.-S. Oak tree canker disease supports arthropod diversity in a natural ecosystem. Plant Path. J. 2014, 30, 43–50. [Google Scholar] [CrossRef][Green Version]
- Jacobsen, R.M.; Birkemore, T.; Sverdrup-Thygeson, A. Priority effects of early successional insects influence late successional fungi in dead wood. Ecol. Evol. 2015, 5, 4896–4905. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, J.; Kakutani, T.; Nishida, T. Influence of resource abundance on the structure of the insect community attracted to fermented tree sap. Ecol. Res. 2005, 20, 405–414. [Google Scholar] [CrossRef]
- Park, M.S.; Fong, J.J.; Lee, H.; Shin, S.; Lee, S.; Lee, N.; Lim, Y.W. Determination of coleopteran insects associated with spore dispersal of Cryptoporus volvatus. J. Asia-Pac. Entomol 2014, 17, 647–651. [Google Scholar] [CrossRef]
- Lawrence, J.F. Coleoptera associated with an Hypoxylon species (Ascomycetes: Xylariaceae) on oak. Coleopt. Bull. 1977, 31, 309–312. [Google Scholar]
- Seibold, S.; Bässler, C.; Baldrian, P.; Thorn, S.; Müller, J.; Gossner, M.M. Wood resource and not fungi attract early-successional saproxylic species of Heteroptera—An experimental approach. Insent. Conserv. Divers. 2014, 7, 533–542. [Google Scholar] [CrossRef]
- Nardi, J.B.; Bee, C.M.; Miller, L.A.; Taylor, S.J. Distinctive features of the alimentary canal of a fungus-feeding hemipteran, Mezira granulata (Heteroptera: Aradidae). Arthropod Struct. Dev. 2009, 38, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Wikars, L.-O. The wood-decaying fungus Daldinia loculata (Xylariaceae) as an indicator of fire-dependent insects. Ecol. Bull. 2001, 49, 263–268. [Google Scholar]
- Vogel, S.; Alvarez, B.; Bässler, C.; Müller, J.; Thorn, S. The red-belted bracket (Fomitopsis pinicola) colonizes spruce trees early after bark beetle attack and persists. Fung. Ecol. 2017, 27, 182–188. [Google Scholar] [CrossRef]
- FFPRI. Naragare Boujo No Shintenkai; Forestry and Forest Products Research Institute: Tsukuba, Japan, 2015; ISBN 978-4-905304-48-7. (In Japanese) [Google Scholar]
- Fukasawa, Y.; Matsukura, K.; Stephan, J.; Makoto, K.; Suzuki, S.N.; Kominami, Y.; Takagi, M.; Tanaka, N.; Takemoto, S.; Kinuura, H.; et al. Patterns of community composition and diversity in latent fungi of living Quercus serrata trunks across a range of oak wilt prevalence and climate variables in Japan. Fung. Ecol. 2021, in press. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Estimate |
|---|---|
| Diameter | –0.03 ** |
| Position (bottom) | –0.34 ** |
| Teleomorph | 0.02 * |
| Anamorph | – |
| d.f. (null) | 444 |
| Null deviance | 814.12 |
| d.f. (residual) | 441 |
| Residual deviance | 770.35 |
| AIC | 1517.1 |
| Variable | Laemophloeus | Librodor | Neuroctenus |
|---|---|---|---|
| Diameter | 0.18 | 0.32 * | 0.20 ** |
| Anamorph area | 0.97 * | 1.17 | – |
| Teleomorph area | 0.08 * | 0.24 * | 0.51 ** |
| d.f. (null) | 63 | 63 | 63 |
| Null deviance | 51.98 | 71.98 | 86.46 |
| d.f. (residual) | 60 | 60 | 61 |
| Residual deviance | 24.47 | 22.51 | 41.33 |
| AIC | 32.47 | 30.51 | 47.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukasawa, Y. Invertebrate Assemblages on Biscogniauxia Sporocarps on Oak Dead Wood: An Observation Aided by Squirrels. Forests 2021, 12, 1124. https://doi.org/10.3390/f12081124
Fukasawa Y. Invertebrate Assemblages on Biscogniauxia Sporocarps on Oak Dead Wood: An Observation Aided by Squirrels. Forests. 2021; 12(8):1124. https://doi.org/10.3390/f12081124
Chicago/Turabian StyleFukasawa, Yu. 2021. "Invertebrate Assemblages on Biscogniauxia Sporocarps on Oak Dead Wood: An Observation Aided by Squirrels" Forests 12, no. 8: 1124. https://doi.org/10.3390/f12081124
APA StyleFukasawa, Y. (2021). Invertebrate Assemblages on Biscogniauxia Sporocarps on Oak Dead Wood: An Observation Aided by Squirrels. Forests, 12(8), 1124. https://doi.org/10.3390/f12081124