
Genetic Diversity Analysis and Potential Distribution Prediction of Sophora moorcroftiana Endemic to Qinghai–Tibet Plateau, China



Abstract
:1. Introduction
2. Materials and Methods
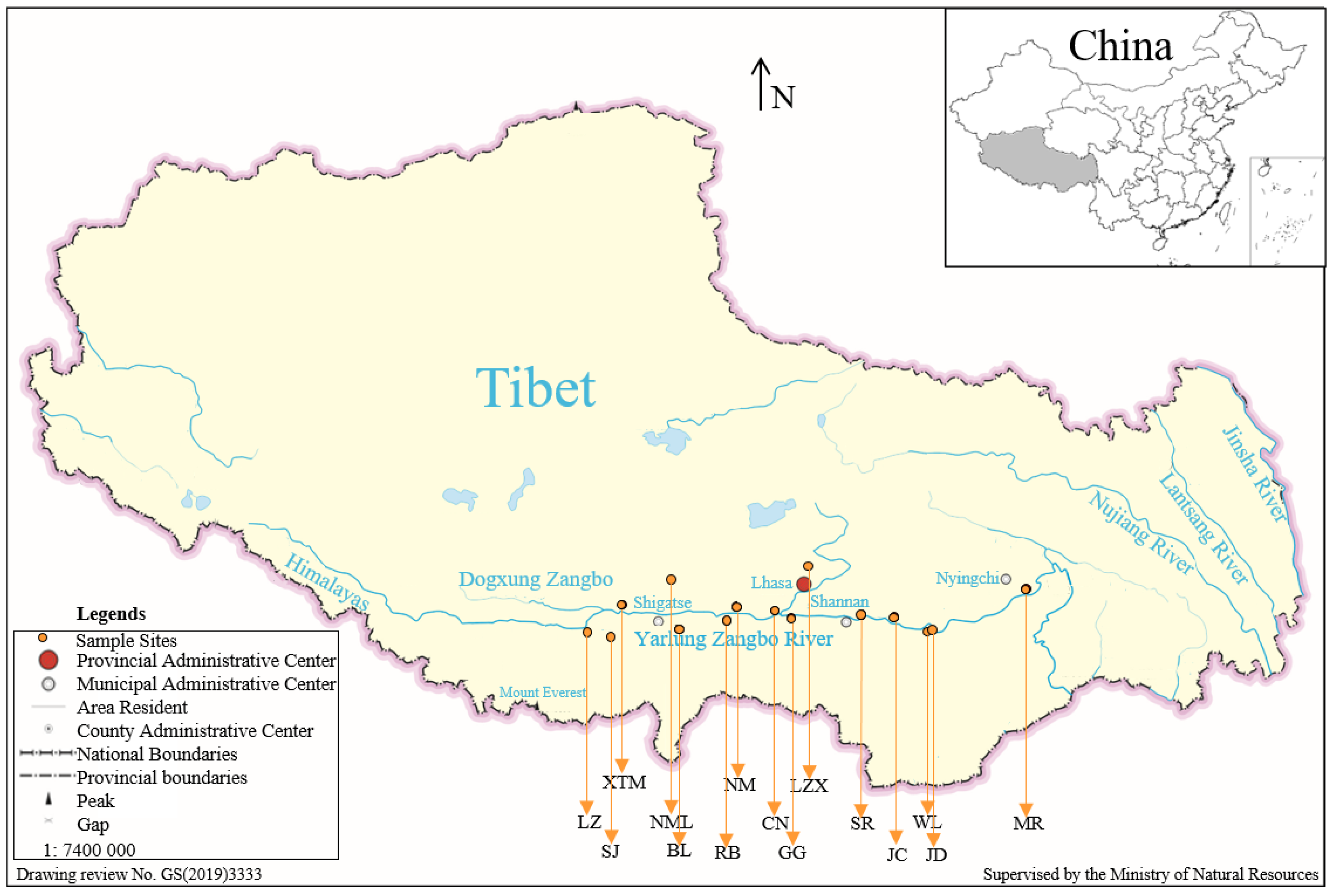
2.1. Materials and Data Sources
2.2. DNA Extraction, Primer Screening, and PCR Amplification
2.3. Genetic Diversity Analysis
2.4. Niche Modelling for Past Two Periods, the Present, and Four Future Periods
3. Results
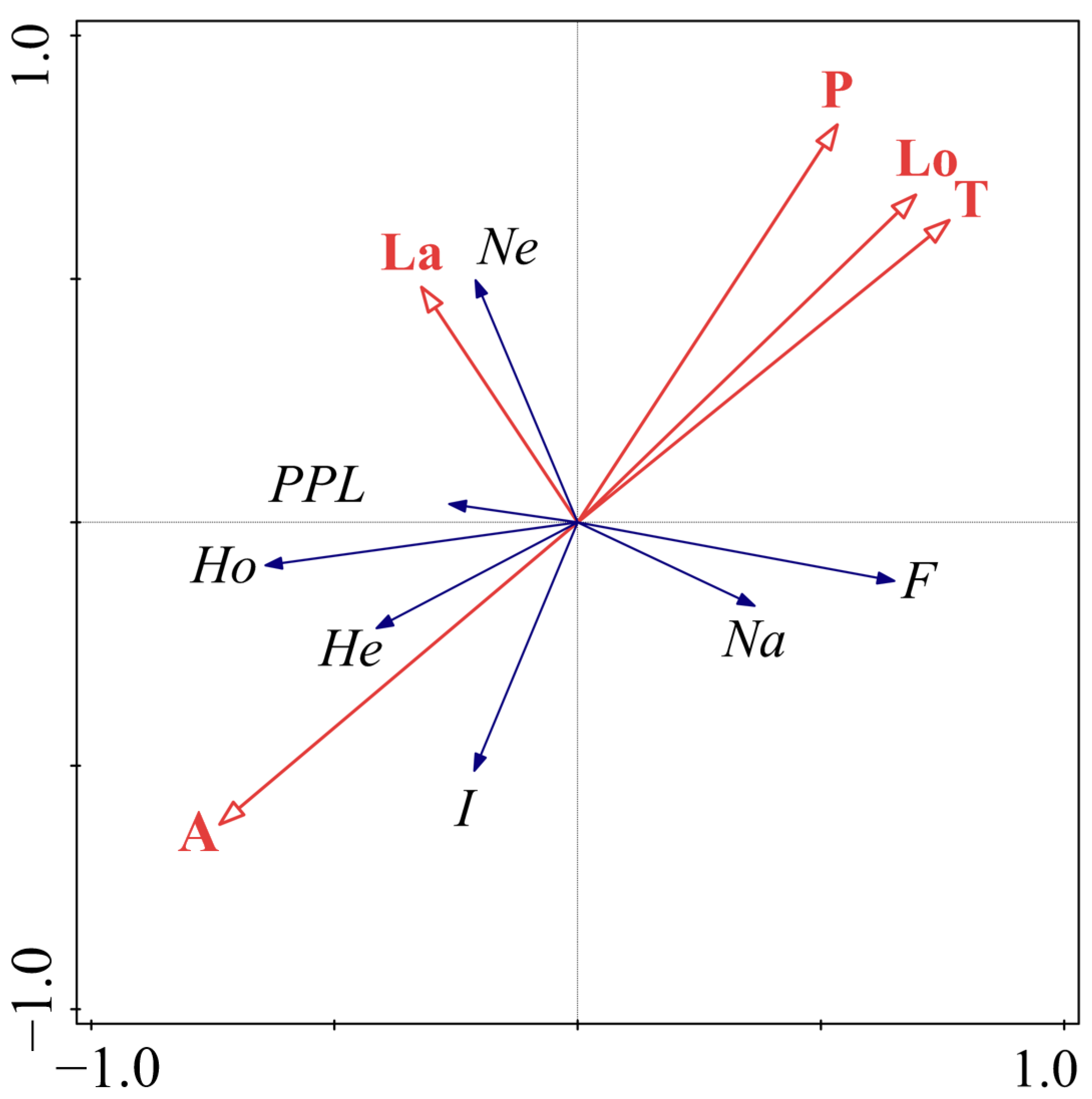
3.1. Genetic Diversity Analysis
3.1.1. Genetic Diversity Analysis of SSR Loci
3.1.2. Genetic Diversity of S. moorcroftiana Population
3.1.3. Genetic Differentiation and Gene Flow among Populations of S. moorcroftiana
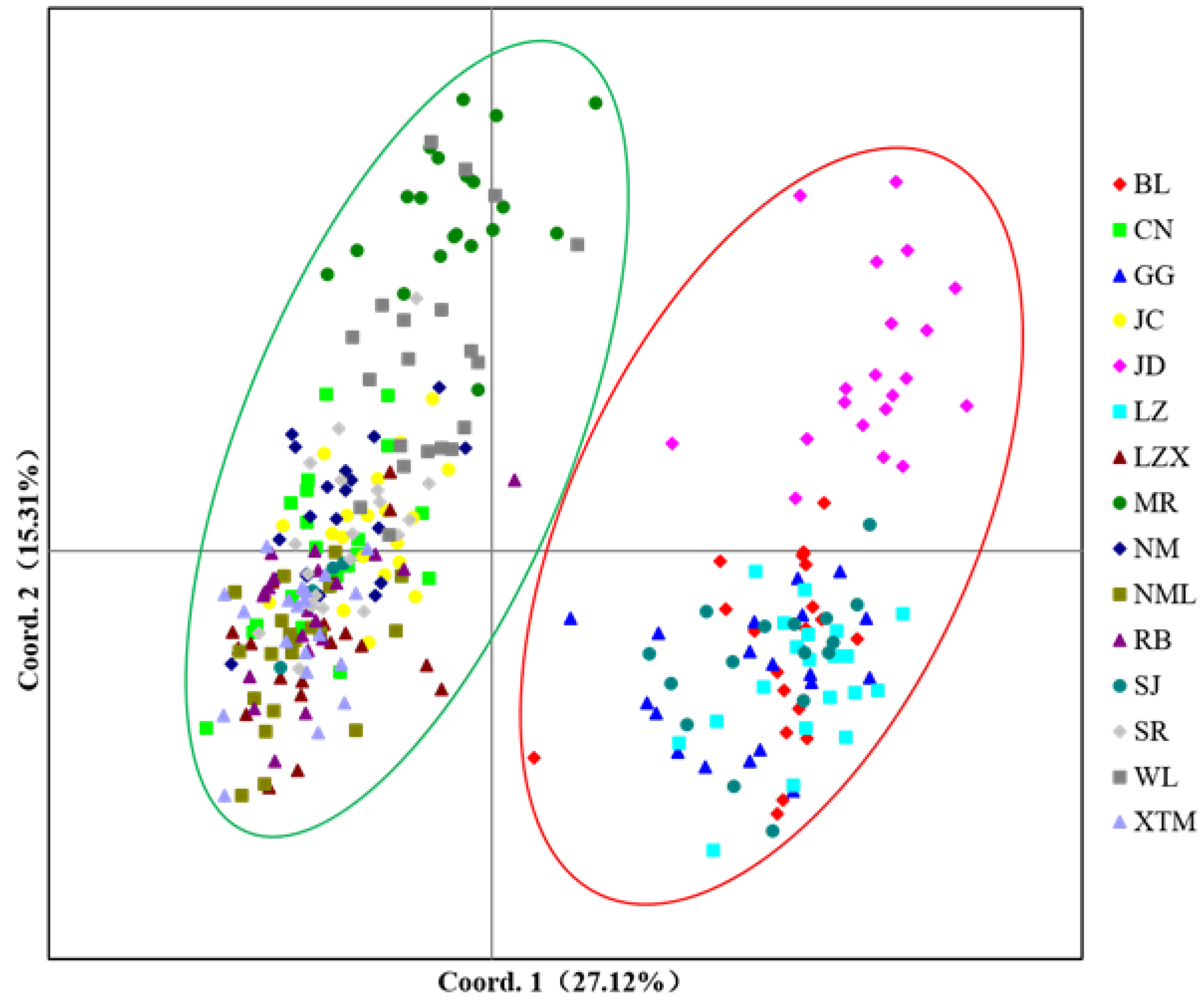
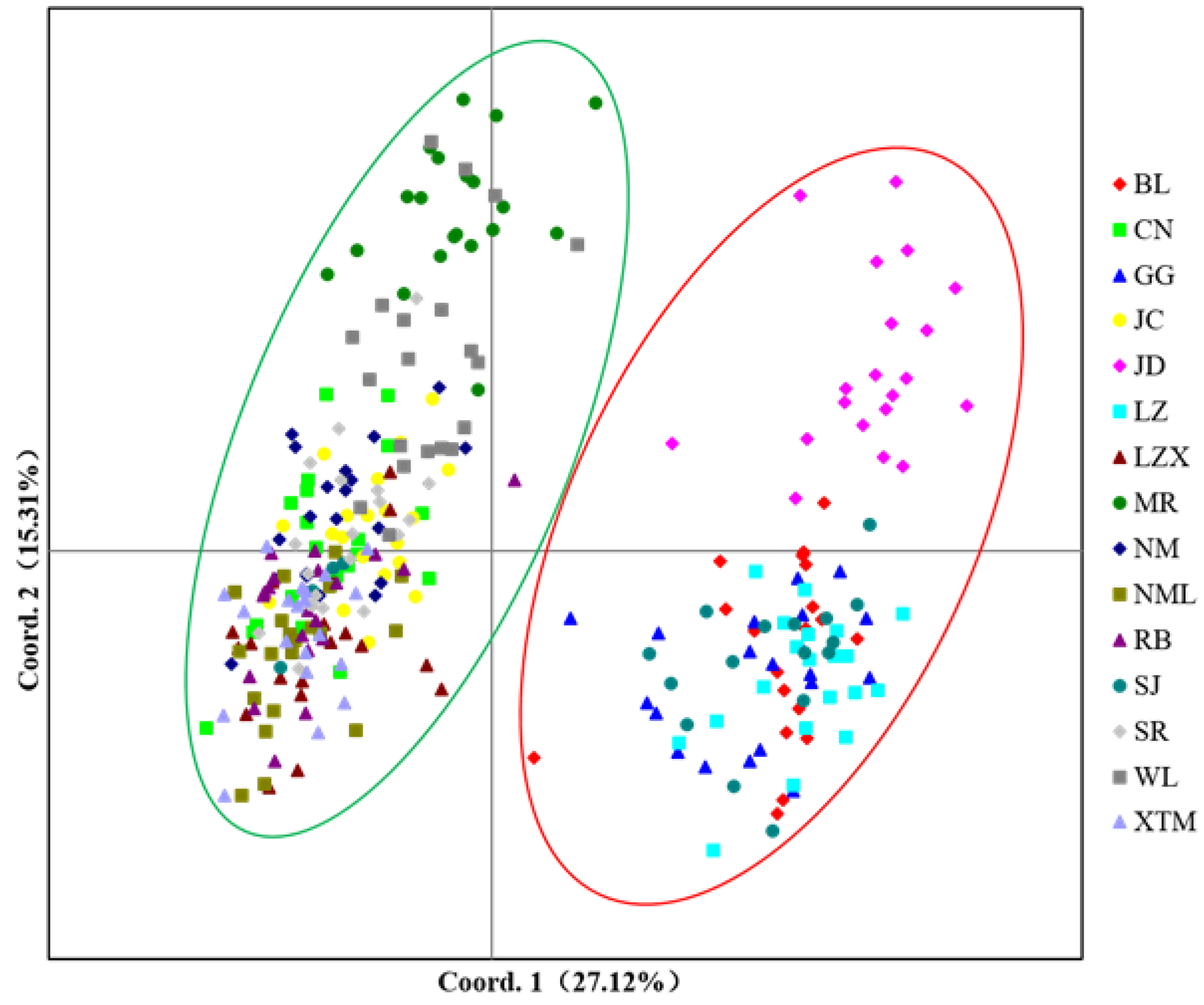
3.1.4. Genetic Distance and Clustering Analysis among Populations of S. moorcroftiana
3.1.5. Bayesian-Inference-Based Genetic Structure Analysis
3.2. Niche Modelling and Analysis
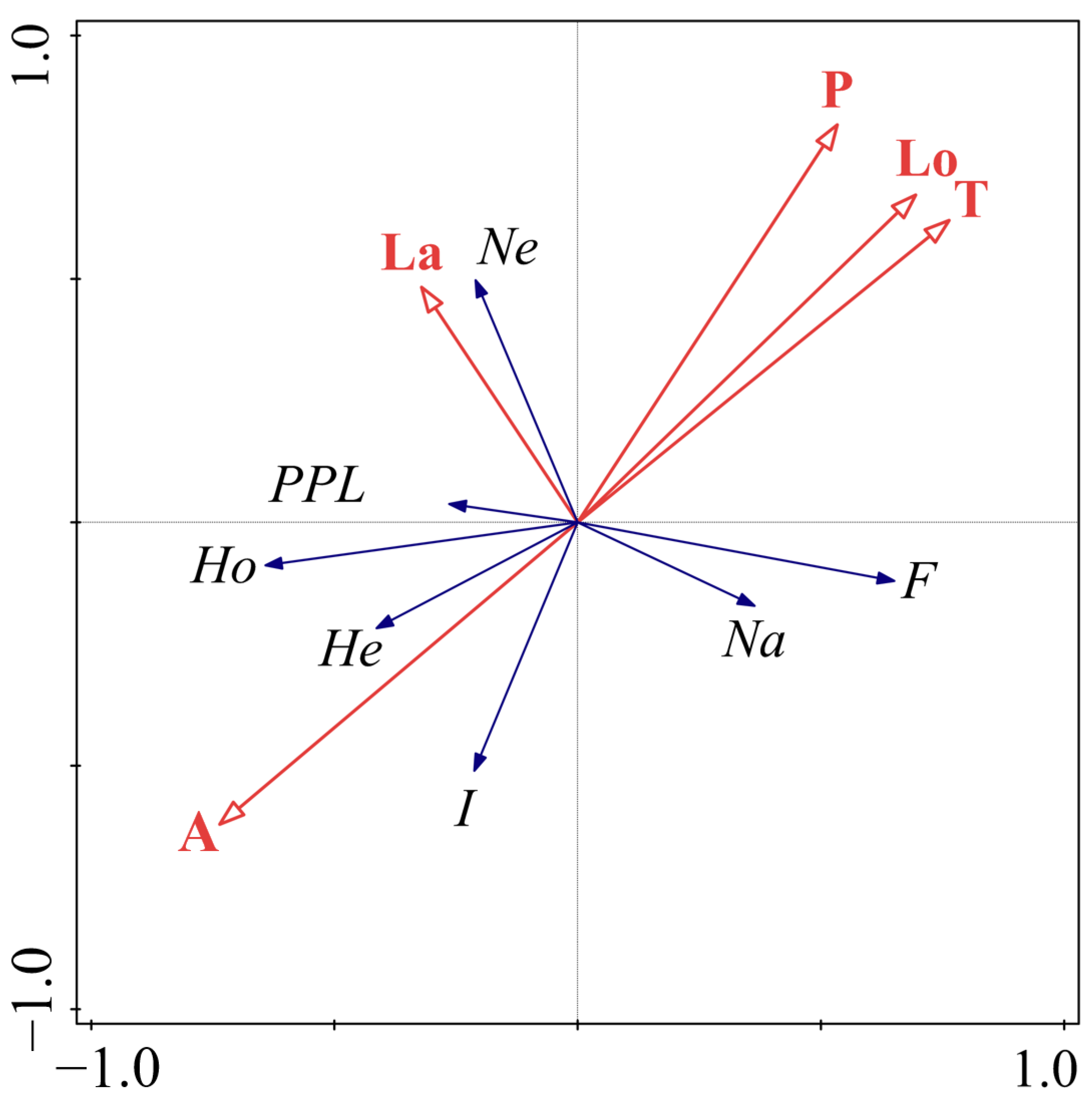
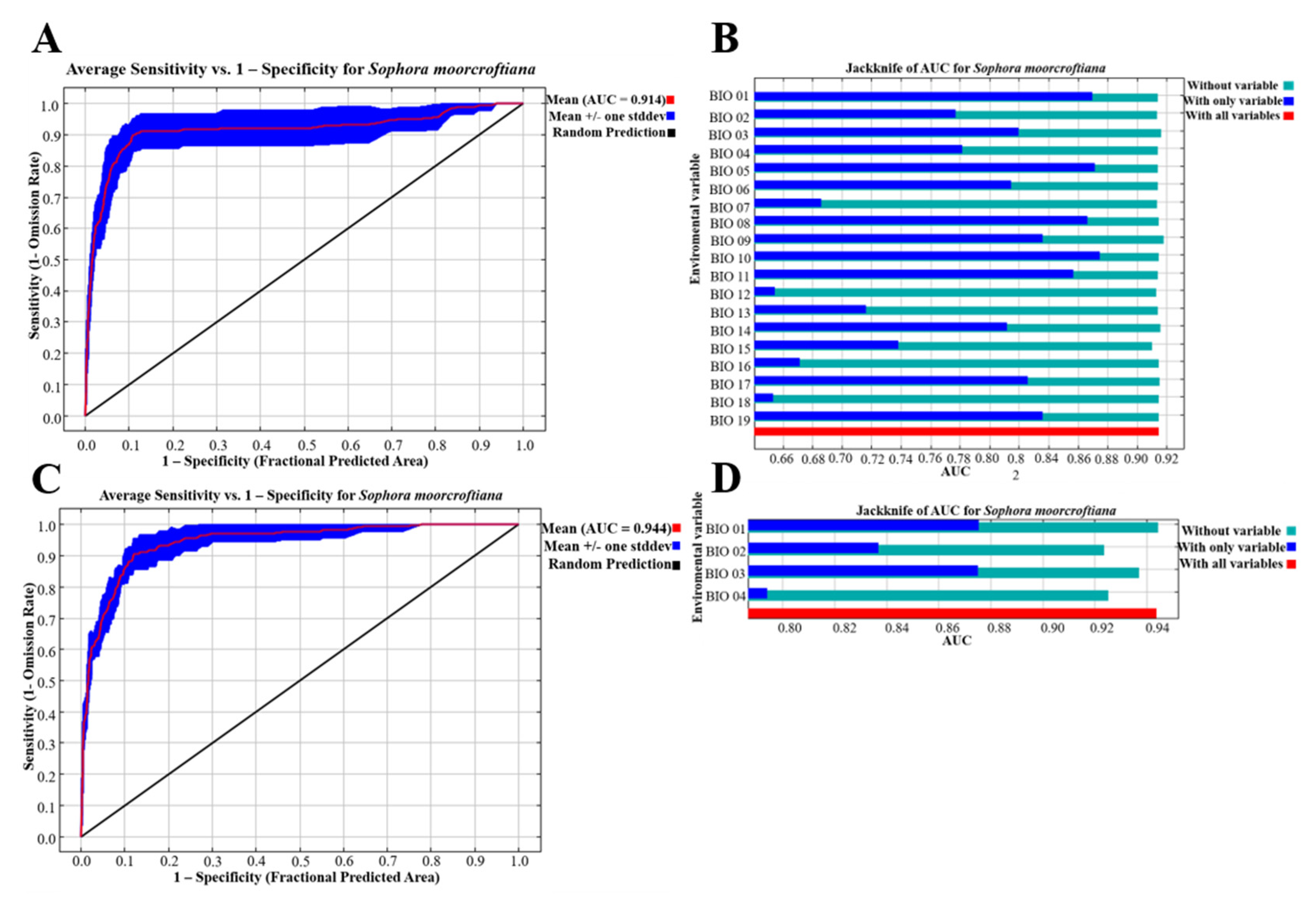
3.2.1. Screening of Critical Environmental Variables
3.2.2. MaxEnt Model Prediction
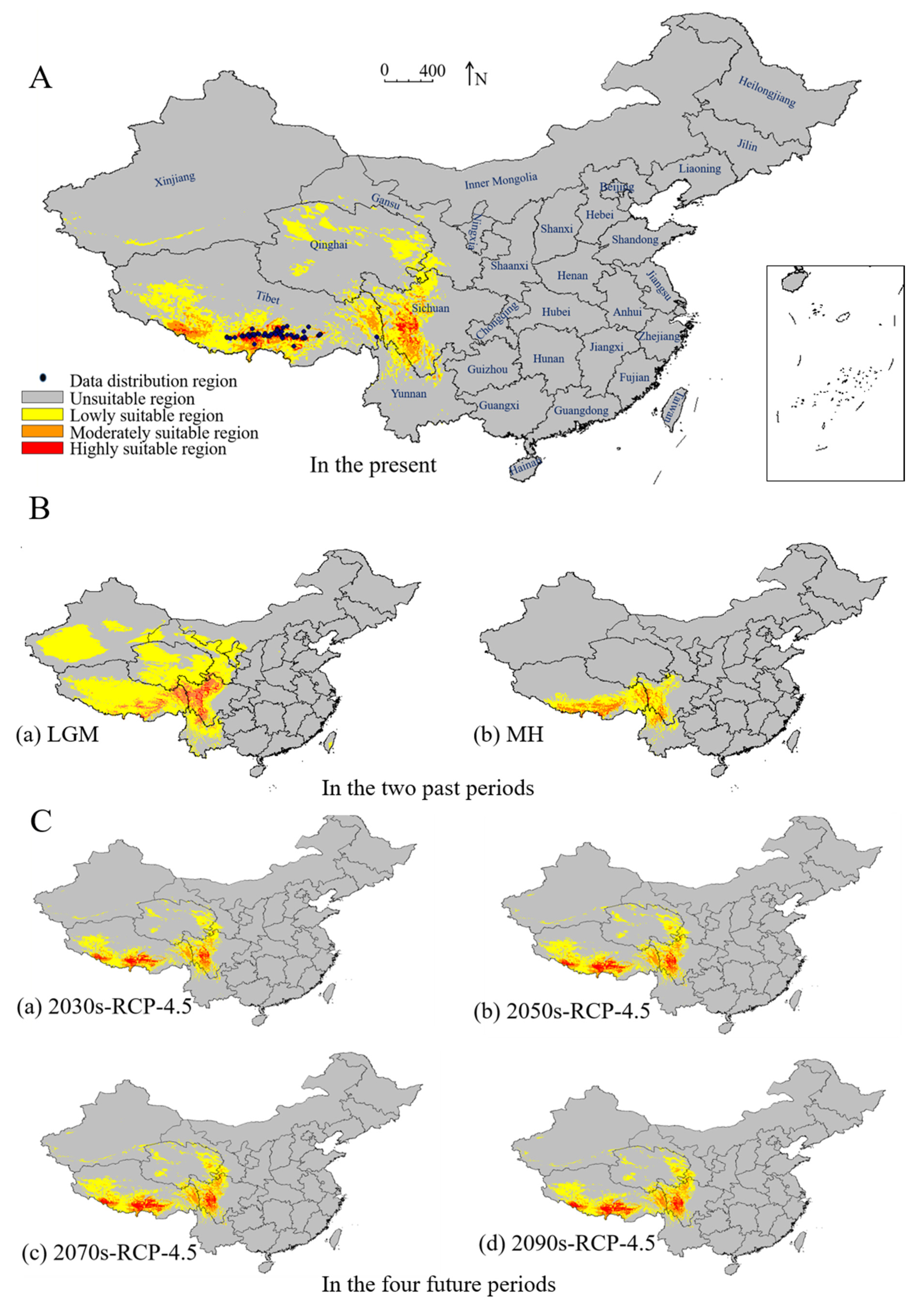
3.2.3. Distribution Trend of S. moorcroftiana during two Past Periods, the Present, and Four Future Periods
3.2.4. Prediction of the Potentially Suitable Areas of S. moorcroftiana in Four Future Periods
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, H.; Cheng, S.; Zhang, Y. The growth of two species of subalpine co-nifer saplings in response to soil warming and inter-competition in Mt. Gongga on the south-eastern fringe of the Qinghai-Tibetan Plateau, China. World J. Eng. Technol. 2016, 4, 398–412. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Shen, X.; Tong, L.; Lei, F.; Mu, X.; Zhang, X. Impact of past and future climate change on the potential distribution of an endangered montane shrub Lonicera oblata and its conservation implications. Forests 2021, 12, 125. [Google Scholar] [CrossRef]
- Ouyang, L.; Arnold, R.J.; Chen, S.; Xie, Y.; He, S.; Liu, X.; Zhang, W. Prediction of the suitable distribution of Eucalyptus grandis in China and its responses to climate change. New For. 2021. (prepublish). [Google Scholar] [CrossRef]
- Kirpotin, S.N.; Callaghan, T.V.; Peregon, A.M.; Babenko, A.S.; Berman, D.I.; Bulakhova, N.A.; Byzaakay, A.A.; Chernykh, T.M.; Chursin, V.; Interesova, E.A. Impacts of environmental change on biodiversity and vegetation dynamics in Siberia. Ambio 2021. Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Gianni, B.; Catherine, P. Effects of climate change on grassland biodiversity and productivity. Agronomy 2021, 11, 1047. [Google Scholar]
- Tang, Z.; Chen, L.; Chen, Z.; Fu, Y.; Sun, X.; Wang, B.; Xia, T. Climatic factors determine the yield and quality of Honghe flue-cured tobacco. Sci. Rep. 2020, 10, 19868. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, T.; Abbo, S.; Sherman, A.; Ophir, R. Prospects for genetic diversity of crop wild relatives under climate change: The case of the wild pea Pisum fulvum. Plant Sci. 2021, 310, 110957. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Guo, L.; He, B.; Lyu, Y.; Li, T. The stability of Qinghai-Tibet Plateau ecosystem to climate change. Phys. Chem. Earth 2019, 115, 102827. [Google Scholar] [CrossRef]
- Hirota, M.; Tang, Y.; Hu, Q.; Kato, T.; Hirata, S.; Mo, W.; Cao, G.; Mariko, S. The potential importance of grazing to the fluxes of carbon dioxide and methane in an alpine wetland on the Qinghai-Tibetan Plateau. Atmosp Environ. 2005, 39, 5255–5259. [Google Scholar] [CrossRef]
- Li, H.; Yang, L.; Li, Q.; Li, J. Completed plastome sequence of Sophora moorcroftiana, an endemic shrub to the Qinghai-Tibet Plateau, China. Mitochond. DNA Part B Res. 2019, 4, 4182–4183. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yi, F.; Yang, G.; Wang, Y.; Pubu, C.; He, R.; Xiao, Y.; Wang, J.; Liu, N.; Wang, J.; et al. Geographic population genetic structure and diversity of Sophora moorcroftiana based on genotyping-by-sequencing (GBS). Peerj 2020, 8, e9609. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Fu, Y.; Zhang, Y.; Li, H. Cloning of four DREB genes from Tibetan Sophora moorcroftiana and analysis of their expression during abiotic stress. North. For. Univ. 2016, 27, 675–683. [Google Scholar] [CrossRef]
- Li, H.; Yao, W.; Fu, Y.; Li, S.; Guo, Q. De novo assembly and discovery of genes that are involved in drought tolerance in Tibetan Sophora moorcroftiana. PLoS ONE 2015, 10, e111054. [Google Scholar]
- Qin, A.; Jin, K.; Batsaikhan, M.-E.; Nyamjav, J.; Li, G.; Li, J.; Xue, Y.; Sun, G.; Wu, L.; Indree, T.; et al. Predicting the current and future suitable habitats of the main dietary plants of the Gobi Bear using MaxEnt modeling. Glob. Ecol. Conserv. 2020, 22, e01032. [Google Scholar] [CrossRef]
- Hsia, A.P.; Chen, H.D.; Ohtsu, K.; Schnable, P.S. DNA extraction from freeze-dried plant tissue with CTAB in a 96-Well format. Cold Spring Harb. Protoc. 2010, 2010, pdb.prot5516. [Google Scholar] [CrossRef] [PubMed]
- You, Q.; Pan, Y.; Xu, L.; Gao, S.; Wang, Q.; Su, Y.; Yang, Y.; Wu, Q.; Zhou, D.; Que, Y. Genetic diversity analysis of sugarcane germplasm based on fluorescence-labeled simple Sequence repeat markers and a capillary electrophoresis-based genotyping platform. Sugar Tech. 2016, 18, 380–390. [Google Scholar] [CrossRef]
- Ollvelra, T.; Perelra, A.; Coppede, J.; França, S.; Ming, L.; Bertonl, B. Genetic diversity analysis of Croton antisyphiliticus Mart. using AFLP molecular markers. Gen. Mol. Res. GMR 2016, 15, gmr.15017461. [Google Scholar]
- Marlon, L.; Massimo, G.; Lorenzo, B.; Erick, O.; Erick, D.; Osman, G.; Alberto, M.; Edgardo, G.; Stefano, B.; Enrico, P. Genetic Purity of cacao criollo from Honduras is revealed by SSR molecular markers. Agronomy 2021, 11, 225. [Google Scholar]
- Reetika, S.; Pratap, K.S.; Nishi, K.; Major, S. Regeneration of soapnut tree through somatic embryogenesis and assessment of genetic fidelity through ISSR and RAPD markers. Physiol. Mol. Biol. Plants Int. J. Funct. Plant Biol. 2016, 22, 381–389. [Google Scholar]
- Qi, Y.; Zhao, W.; Huang, Y.; Wang, X.; Zhao, Y. Correlation between climatic factors and genetic diversity of Phrynocephalus forsythii. Asian Herpetol. Res. 2019, 10, 270–275. [Google Scholar]
- Nyabera, L.; Nzuki, I.; Runo, S.; Amwayi, P. Assessment of genetic diversity of pumpkins (Cucurbita spp.) from western Kenya using SSR molecular markers. Mol. Biol. Rep. 2021, 48, 2253–2260. [Google Scholar] [CrossRef]
- Warren, D.; Matzke, N.; Cardillo, M.; Baumgartner, J.; Beaumont, L.; Turelli, M.; Glor, R.; Huron, N.; SimÕes, M.; Lglesias, T.; et al. ENMTools 1.0: An R package for comparative ecological biogeography. Ecography 2021, 44, 504–511. [Google Scholar] [CrossRef]
- Sivaraj, N.; Elangovan, M.; Kamala, V.; Pandravada, S.; Pranusha, P.; Chakrabarty, S. Maximum entropy (Maxent) approach to Sorghum Landraces distribution modelling. Ind. J. Plant Gen. Res. 2016, 29, 16–21. [Google Scholar] [CrossRef]
- Cobos, M.; Peterson, A.; Barve, N.; Olvera, L. Kuenm: An R package for detailed development of ecological niche models using Maxent. Peerj 2019, 7, e6281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruthi, R.; Kumar, A.; Choudhary, S.; Sharma, H.; Mitra, J. DIVA-GIS based insight into geographical distribution and diversity spectrum of indian sunnhemp (Crotalaria juncea L.) accessions suitable for diversified applications. Leg. Res. Int. J. 2020, 43, 749–756. [Google Scholar] [CrossRef]
- Swets, J. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, G.; Zhang, F.; Gao, Q.; Fu, P.-C.; Xing, R.; Wang, J.; Liu, H.; Chen, S. Molecular phylogeography and intraspecific divergence of Spiraea alpina (Rosaceae) distributed in the Qinghai-Tibetan Plateau and adjacent regions inferred from nrDNA. Biochem. Syst. Ecol. 2014, 57, 278–286. [Google Scholar] [CrossRef]
- Liu, W.; Zhao, Y.; Qi, D.; You, J.; Zhou, Z.; Song, Z. The Tanggula Mountains enhance population divergence in Carex moorcroftii: A dominant sedge on the Qinghai-Tibetan Plateau. Sci. Rep. 2018, 8, 2741. [Google Scholar] [CrossRef]
- Cheng, S.; Qiong, L.; Lu, F.; Yonezawa, T.; Yin, G.; Song, Z.; Wang, Y.; Yang, J.; Zhang, W. Phylogeography of Sophora moorcroftiana supports Wu’s hypothesis on the origin of Tibetan Alpine Flora. J. Heredity 2017, 108, 405–414. [Google Scholar] [CrossRef]
- Zhang, M.; Xiang, X.; Xue, J.; Sanderson, S.; Firtsch, P. Himalayan uplift shaped biomes in Miocene temperate Asia: Evidence from leguminous Caragana. Sci. Rep. 2016, 6, 36528. [Google Scholar] [CrossRef]
- Zhao, Z.; Kang, D.; Guo, W.; Zhao, L.; Li, C.; Li, J. Climate sensitivity of purple cone spruce (Picea purpurea) across an altitudinal gradient on the eastern Tibetan Plateau. Dendrochronologia 2019, 56, 125586. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population Name | Population Code | Longitude | Latitude | Altitude (m) | Annual Average Temperature (°C) | Annual Average Precipitation (mm) |
|---|---|---|---|---|---|---|
| Jiding Township, Sajia County, Shigatse | SJ | 87°38′13″ | 29°05′12″ | 4087 | 3.6 | 242 |
| Chawu Township, Lazi County, Shigatse | LZX | 87°34′80″ | 29°06′56″ | 4010 | 3.9 | 251 |
| Dongga Township, Xietongmeng County, Shigatse | XTM | 88°50′90″ | 29°21′26″ | 3855 | 4.1 | 269 |
| Bazha Township, Bailang County, Shigatse | BL | 89°02′20″ | 29°09′40″ | 3845 | 4.3 | 274 |
| Aima Township, Nanmulin County, Shigatse | NML | 89°06′43″ | 29°18′49″ | 3819 | 5.1 | 279 |
| Qiewa Township, Renbu County, Shigatse | RB | 89°37′30″ | 29°20′30″ | 3782 | 5.8 | 282 |
| Nimu Township, Nimu County, Lhasa | NM | 90°10′40″ | 29°19′27″ | 3738 | 5.9 | 344 |
| Kazi Township, Linzhou County, Lhasa | LZ | 91°17′34″ | 29°41′22″ | 3709 | 6.7 | 374 |
| Caina Township, Lhasa | CN | 91°06′52″ | 29°44′59″ | 3622 | 6.9 | 412 |
| Jiazhulin Township, Gongga County, Shannan | GG | 90°54′12″ | 29°17′28″ | 3590 | 7.3 | 396 |
| Rong Township, Sangri County, Shannan | SR | 91°52′28″ | 29°15′53″ | 3582 | 7.4 | 377 |
| Jiacha Township, Jiacha County, Shannan | JC | 92°35′52″ | 29°08′39″ | 3251 | 7.7 | 364 |
| Wolong Township, Nyingchi | WL | 93°15′35″ | 29°00′14″ | 3157 | 7.9 | 492 |
| Jindong Township, Nyingchi | JD | 93°19′55″ | 29°01′43″ | 3048 | 8.6 | 567 |
| Mirui Township, Nyingchi | MR | 94°31′00″ | 29°27′40″ | 2931 | 8.8 | 591 |
| Pop | N | Na | Ne | I | Ho | He | PPL (%) | F |
|---|---|---|---|---|---|---|---|---|
| BL | 20 | 4.65 | 2.808 | 0.986 | 0.563 | 0.491 | 85.00% | −0.104 |
| CN | 20 | 4.05 | 2.485 | 0.868 | 0.543 | 0.445 | 90.00% | −0.128 |
| GG | 20 | 5.45 | 3.240 | 1.116 | 0.639 | 0.534 | 90.00% | −0.167 |
| JC | 20 | 4.50 | 2.727 | 0.967 | 0.648 | 0.497 | 90.00% | −0.276 |
| JD | 20 | 4.45 | 2.606 | 0.944 | 0.523 | 0.477 | 85.00% | −0.043 |
| LZ | 20 | 4.40 | 2.769 | 0.944 | 0.541 | 0.471 | 90.00% | −0.107 |
| LZX | 20 | 5.35 | 3.196 | 1.062 | 0.573 | 0.500 | 80.00% | −0.146 |
| MR | 20 | 3.70 | 2.266 | 0.816 | 0.516 | 0.437 | 90.00% | −0.132 |
| NM | 20 | 4.50 | 2.702 | 0.907 | 0.551 | 0.448 | 90.00% | −0.186 |
| NML | 20 | 4.90 | 2.719 | 0.969 | 0.560 | 0.469 | 95.00% | −0.154 |
| RB | 20 | 5.55 | 3.250 | 1.095 | 0.585 | 0.512 | 90.00% | −0.087 |
| SJ | 20 | 5.50 | 3.043 | 1.118 | 0.576 | 0.539 | 95.00% | 0.003 |
| SR | 20 | 5.25 | 3.214 | 1.076 | 0.570 | 0.513 | 90.00% | −0.074 |
| WL | 20 | 4.75 | 2.867 | 1.014 | 0.568 | 0.502 | 90.00% | −0.100 |
| XTM | 20 | 5.05 | 3.279 | 1.024 | 0.566 | 0.493 | 90.00% | −0.092 |
| mean | 20 | 4.80 | 2.878 | 0.994 | 0.568 | 0.489 | 89.33% | −0.120 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Li, H.; Li, Q.; Guo, Q.; Li, J. Genetic Diversity Analysis and Potential Distribution Prediction of Sophora moorcroftiana Endemic to Qinghai–Tibet Plateau, China. Forests 2021, 12, 1106. https://doi.org/10.3390/f12081106
Yang L, Li H, Li Q, Guo Q, Li J. Genetic Diversity Analysis and Potential Distribution Prediction of Sophora moorcroftiana Endemic to Qinghai–Tibet Plateau, China. Forests. 2021; 12(8):1106. https://doi.org/10.3390/f12081106
Chicago/Turabian StyleYang, Lan, Huie Li, Qian Li, Qiqiang Guo, and Jiangrong Li. 2021. "Genetic Diversity Analysis and Potential Distribution Prediction of Sophora moorcroftiana Endemic to Qinghai–Tibet Plateau, China" Forests 12, no. 8: 1106. https://doi.org/10.3390/f12081106
APA StyleYang, L., Li, H., Li, Q., Guo, Q., & Li, J. (2021). Genetic Diversity Analysis and Potential Distribution Prediction of Sophora moorcroftiana Endemic to Qinghai–Tibet Plateau, China. Forests, 12(8), 1106. https://doi.org/10.3390/f12081106