
Ready for Screening: Fast Assessable Hydraulic and Anatomical Proxies for Vulnerability to Cavitation of Young Conifer Sapwood


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Harvest
2.2. Reference Values of Hydraulic Vulnerability to Cavitation (P50, P12, P88)
2.3. Relative Water Loss and Calculation of P25W
2.4. Basic Wood Density
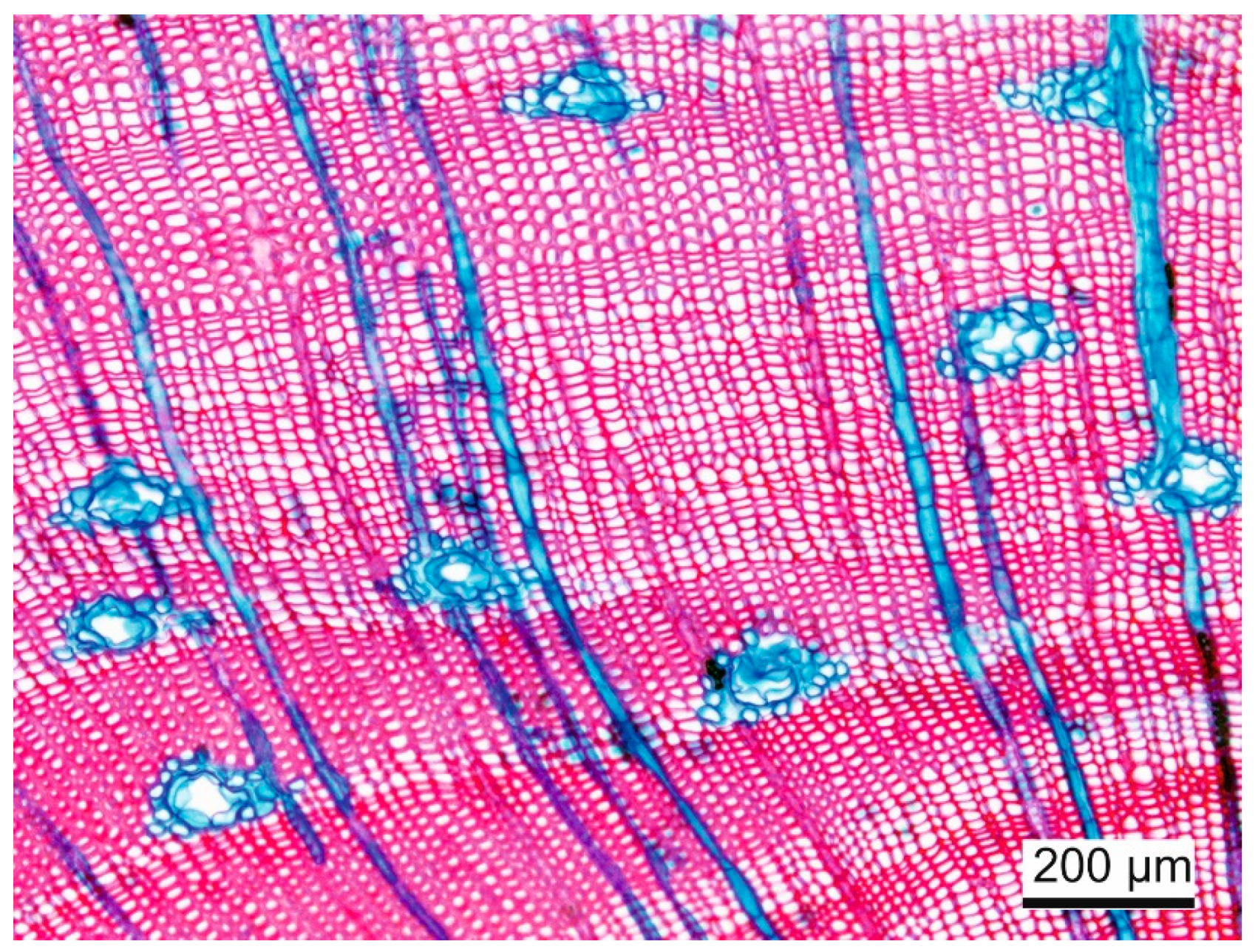
2.5. Wood Anatomy
2.6. Sample Numbers, Data Processing and Statistical Analyses
3. Results
3.1. Range of Hydraulic Vulnerabilities
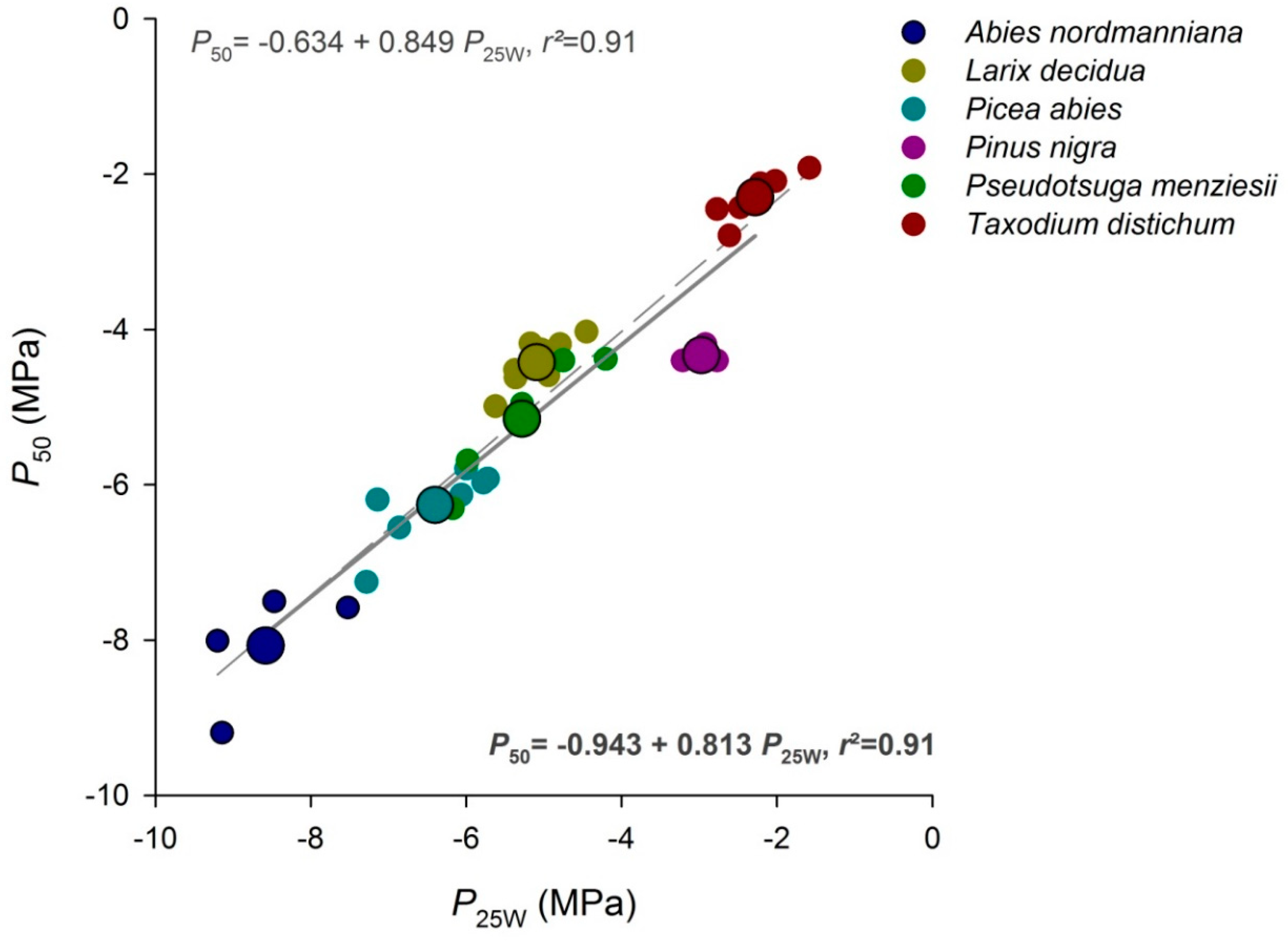
3.2. Relationships between Hydraulic Traits
3.3. Structure–Function Relationships
4. Discussion
4.1. The Hydraulic Capacitance Parameter P25W Is the Best Proxy for Vulnerability to Cavitation
4.2. Tangentially Measured Anatomical Traits Have a Higher Predictive Quality for Hydraulic Failure
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McDowell, N.G.; Allen, C.D. Darcy’s law predicts widespread forest mortality under climate warming. Nat. Clim. Chang. 2015, 5, 669. [Google Scholar] [CrossRef]
- Brodribb, T.; Powers, J.; Cochard, H.; Choat, B. Hanging by a thread? Forests and drought. Science 2020, 368, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Breshears, D.D.; Fontaine, J.B.; Ruthrof, K.X.; Field, J.P.; Feng, X.; Burger, J.R.; Law, D.J.; Kala, J.; Hardy, G.E.S.J. Underappreciated plant vulnerabilities to heat waves. New Phytol. 2021, 231, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.; Cahanovitc, R.; Sprintsin, M.; Herr, N.; Schiller, G. A nation-wide analysis of tree mortality under climate change: Forest loss and its causes in Israel 1948–2017. For. Ecol. Managem. 2019, 432, 840–849. [Google Scholar] [CrossRef]
- Mencuccini, M.; Binks, O. Tall, leafy conifers lose out. Nat. Clim. Chang. 2015, 5, 625–626. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 129. [Google Scholar] [CrossRef]
- Cochard, H. A new mechanism for tree mortality due to drought and heatwaves. Wood Sci. 2020, 1, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Cochard, H.; Delzon, S. Hydraulic failure and repair are not routine in trees. Ann. For. Sci. 2013, 70, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; Lopez, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef]
- Hammond, W.M.; Yu, K.L.; Wilson, L.A.; Will, R.E.; Anderegg, W.R.; Adams, H.D. Dead or dying? Quantifying the point of no return from hydraulic failure in drought-induced tree mortality. New Phytol. 2019, 223, 1834–1843. [Google Scholar] [CrossRef] [Green Version]
- Tyree, M.T.; Sperry, J.S. Do woody plants operate near the point of catastrophic xylem dysfunction caused by dynamic water stress? Answers from a model. Plant Physiol. 1988, 88, 574–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Field, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [Green Version]
- Delzon, S.; Cochard, H. Recent advances in tree hydraulics highlight the ecological significance of the hydraulic safety margin. New Phytol. 2014, 203, 355–358. [Google Scholar] [CrossRef] [PubMed]
- Cochard, H.; Badel, E.; Herbette, S.; Delzon, S.; Choat, B.; Jansen, S. Methods for measuring plant vulnerability to cavitation: A critical review. J. Exp. Bot. 2013, 64, 4779–4791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacke, U.G.; Venturas, M.D.; MacKinnon, E.D.; Jacobsen, A.L.; Sperry, J.S.; Pratt, R.B. The standard centrifuge method accurately measures vulnerability curves of long-vesselled olive stems. New Phytol. 2015, 205, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.M.; McCulloh, K.A.; Woodruff, D.R.; Meinzer, F.C. Hydraulic safety margins and embolism reversal in stems and leaves: Why are conifers and angiosperms so different? Plant Sci. 2012, 135, 4–53. [Google Scholar] [CrossRef]
- McCulloh, K.A.; Johnson, D.M.; Meinzer, F.C.; Woodruff, D.R. The dynamic pipeline: Hydraulic capacitance and xylem hydraulic safety in four tall conifer species. Plant Cell Environ. 2014, 37, 1171–1183. [Google Scholar] [CrossRef] [Green Version]
- Rosner, S.; Heinze, B.; Savi, T.; Dalla-Salda, G. Prediction of hydraulic conductivity loss from relative water loss: New insights into water storage of tree stems and branches. Physiol. Plant 2019, 165, 843–854. [Google Scholar] [CrossRef] [Green Version]
- Rosner, S.; Johnson, D.M.; Voggeneder, K.; Domec, J.-C. The conifer-curve: Fast prediction of hydraulic conductivity loss and vulnerability to cavitation. Ann. For. Sci. 2019, 76, 82. [Google Scholar] [CrossRef] [Green Version]
- Hacke, U.G.; Sperry, J.S.; Pockman, W.T.; Davis, S.D.; McCulloh, K.A. Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 2001, 126, 457–461. [Google Scholar] [CrossRef]
- Hacke, U.G.; Lachenbruch, B.; Pitterman, J.; Mayr, S.; Domec, J.-C.; Schulte, P.J. The Hydraulic Architecture of Conifers. In Functional and Ecological Xylem Anatomy; Hacke, U.G., Ed.; Springer International Publishing: Basel, Switzerland, 2015; pp. 39–75. [Google Scholar]
- Bouche, P.S.; Larter, M.; Domec, J.-C.; Burlett, R.; Gasson, P.; Jansen, S.; Delzon, S. A broad survey of hydraulic and mechanical safety in the xylem of conifers. J. Exp. Bot. 2014, 65, 4419–4431. [Google Scholar] [CrossRef] [Green Version]
- Rosner, S.; Karlsson, B.; Konnerth, J.; Hansmann, C. Shrinkage processes in standard-size Norway spruce wood specimens with different vulnerability to cavitation. Tree Physiol. 2009, 29, 1419–1431. [Google Scholar] [CrossRef]
- Luss, S.; Lundqvist, S.-O.; Evans, R.; Grahn, T.; Olsson, L.; Petit, G.; Rosner, S. Within-ring variability of wood structure and its relationship to drought sensitivity in Norway spruce trunks. IAWA J. 2019, 40, 288–310. [Google Scholar] [CrossRef]
- Domec, J.-C.; Warren, J.M.; Meinzer, F.C.; Lachenbruch, B. Safety for xylem failure by implosion and air-seeding within roots, trunks and branches of young and old conifer trees. IAWA J. 2009, 30, 101–120. [Google Scholar] [CrossRef] [Green Version]
- Hereş, A.-M.; Camarero, J.J.; López, B.C.; Martínez-Vilalta, J. Declining hydraulic performances and low carbon investments in tree rings predate Scots pine drought-induced mortality. Trees 2014, 28, 1737–1750. [Google Scholar] [CrossRef]
- Rosner, S.; Světlík, J.; Andreassen, K.; Børja, I.; Dalsgaard, L.; Evans, R.; Luss, S.; Tveito, O.E.; Solberg, S. Novel hydraulic vulnerability proxies for a boreal conifer species reveal that opportunists may have lower survival prospects under extreme climatic events. Front. Plant Sci. 2016, 7, 831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasani, N.; Pâques, L.E.; Boulanger, G.; Singh, A.P.; Gierlinger, N.; Rosner, S.; Brendel, O. Physiological and anatomical responses to drought stress differ between two larch species and their hybrid. Trees 2021, in press. [Google Scholar] [CrossRef]
- Rosner, S. Wood density as a proxy for vulnerability to cavitation: Size matters. J. Plant Hydraul. 2017, 4, e001. [Google Scholar] [CrossRef]
- Boden, S.; Schinker, M.G.; Duncker, P.; Spiecker, H. Resolution abilities and measuring depth of high-frequency densitometry on wood samples. Measurement 2012, 45, 1913–1921. [Google Scholar] [CrossRef]
- Björklund, J.; von Arx, G.; Nievergelt, D.; Wilson, R.; Van den Bulcke, J.; Günther, B.; Loader, N.J.; Rydval, M.; Fonti, P.; Scharnweber, T.; et al. Scientific Merits and Analytical Challenges of Tree-Ring Densitometry. Rev. Geophys. 2019, 57, 1224–1264. [Google Scholar] [CrossRef] [Green Version]
- Cochard, H.; Cruizat, P.; Tyree, M.T. Use of positive pressures to establish vulnerability curves. Further support for the air-seeding hypothesis and implications for pressure-volume analysis. Plant Physiol. 1992, 100, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Salleo, S.; Hinckley, T.M.; Kikuta, S.B.; Lo Gullo, M.A.; Weilgony, P.; Yoon, T.M.; Richter, H. A method for inducing xylem emboli in situ: Experiments with a field-grown tree. Plant Cell Environ. 1992, 15, 491–497. [Google Scholar] [CrossRef]
- Sergent, A.-S.; Varela, S.A.; Barigah, T.S.; Badel, E.; Cochard, H.; Dalla-Salda, G.; Delzon, S.; Fernández, M.E.; Guillemot, J.; Gyenge, J.; et al. A comparison of five methods to assess embolism resistance in trees. For. Ecol. Manag. 2020, 468, 118175. [Google Scholar] [CrossRef]
- Hietz, P.; Rosner, S.; Sorz, J.; Mayr, S. Comparison of methods to quantify loss of hydraulic conductivity in Norway spruce. Ann. For. Sci. 2008, 65, 502. [Google Scholar] [CrossRef]
- Domec, J.-C.; Gartner, B.L. Age- and position-related changes in hydraulic versus mechanical dysfunction of xylem: Inferring the design criteria for Douglas-fir wood structure. Tree Physiol. 2002, 22, 91–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pammenter, N.W.; VanderWilligen, C. A mathematical and statistical analysis of the curves illustrating vulnerability of xylem to cavitation. Tree Physiol. 1998, 18, 589–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, O.R.; Kursar, T.A.; Cochard, H.; Tyree, M.T. Interspecific variation in xylem vulnerability to cavitation among tropical tree and shrub species. Tree Physiol. 2005, 25, 1553–1562. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Meth. 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Savi, T.; Casolo, V.; Borgo, A.; Rosner, S.; Torboli, V.; Stenni, B.; Bertoncin, P.; Martellos, S.; Pallavicini, A.; Nardini, A. Drought-induced dieback of Pinus nigra: A tale of hydraulic failure and carbon starvation. Conserv. Physiol. 2019, 7, coz012. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Pitterman, J. Analysis of circular bordered pit function II. Gymnosperm tracheids with torus-margo pit membranes. Am. J. Bot. 2004, 91, 386–400. [Google Scholar] [CrossRef] [Green Version]
- Grossoni, P. Pinus nigra. Enzyklopädie der Holzgewächse 2006, 19, 1–14. [Google Scholar]
- LaPasha, C.A.; Wheeler, E.A. Resin canals in Pinus taeda-longitudinal canal lengths and interconnections between longitudinal and radial canals. IAWA J. 1990, 11, 227–238. [Google Scholar] [CrossRef]
- Rosner, S. Hydraulic and biomechanical optimization in Norway spruce trunkwood: A review. IAWA J. 2013, 34, 365–390. [Google Scholar] [CrossRef] [Green Version]
- Piermattei, A.; von Arx, G.; Avanzi, C.; Fonti, P.; Gärtner, H.; Piotti, A.; Urbinati, C.; Vendramin, G.G.; Büntgen, U.; Crivellaro, A. Functional relationships of wood anatomical traits in Norway spruce. Front. Plant Sci. 2020, 11, 683. [Google Scholar] [CrossRef] [PubMed]
- Anfodillo, T.; Petit, G.; Crivellaro, A. Axial conduit widening in woody species: A still neglected anatomical pattern. IAWA J. 2013, 34, 352–364. [Google Scholar] [CrossRef] [Green Version]
- Olson, M.E.; Anfodillo, T.; Gleason, S.M.; McCulloh, K.A. Tip-to-base xylem conduit widening as an adaptation: Causes, consequences, and empirical priorities. New Phytol. 2021, 229, 1877–1893. [Google Scholar] [CrossRef]
- Larson, P.R. The Vascular Cambium—Development and Structure; Springer: Berlin/Heidelberg, Germany, 1994. [Google Scholar]
- Petit, G.; Crivellaro, A. Comparative axial widening of phloem and xylem conduits in small woody plants. Trees 2014, 28, 915–921. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Age | Trees | Organ | Samples | Dataset |
|---|---|---|---|---|---|
| Abies nordmanniana | 4 | 3 | stem | 4 | 27 |
| Larix decidua | 20 | 2 | branch | 8 | 52 |
| Picea abies | 4 | 8 | stem | 8 | 54 |
| Pinus nigra | 20 | 3 | branch | 3 | 20 |
| Pseudotsuga menziesii | 4 | 5 | stem | 5 | 37 |
| Taxodium distichum | 20 | 3 | branch | 6 | 36 |
| Abbreviation | Trait | Unit |
|---|---|---|
| P12 | Water potential resulting in 12% conductivity loss | MPa |
| P50 | Water potential resulting in 50% conductivity loss | MPa |
| P88 | Water potential resulting in 88% conductivity loss | MPa |
| P25W | Water potential resulting in 25% relative water loss | MPa |
| BD | Basic wood density | kg m−3 |
| br | Radial lumen diameter of earlywood tracheids | µm |
| tr | Thickness of the radial double cell wall of earlywood tracheids | µm |
| (tr/br)2 | Radial conduit wall reinforcement (tangential force direction) | dimensionless |
| bt | Tangential lumen diameter of earlywood tracheids | µm |
| tt | Thickness of the tangential double cell wall of earlywood tracheids | µm |
| (tt/bt)2 | Tangential conduit wall reinforcement (radial force direction) | dimensionless |
| b | Mean lumen diameter (br + bt)/2 | µm |
| t | Mean double cell wall thickness (tr + tt)/2 | µm |
| (t/b)2 | Mean conduit wall reinforcement (((tr/br)2 + (tt/bt)2)/2) | Dimensionless |
| CF/mm | Number of radial cell files per tangential distance of 1 mm | n/mm |
| Species | P50 [MPa] | P12 [MPa] | P88 [MPa] | P25W [MPa] |
|---|---|---|---|---|
| Abies nordmanniana | −8.07 ± 0.78 | −4.11 ± 1.62 | −12.24 ± 0.62 | −8.58 ± 0.78 |
| Larix decidua | −4.42 ± 0.31 | −2.70 ± 0.26 | −6.14 ± 0.55 | −5.09 ± 0.37 |
| Picea abies | −6.26 ± 0.46 | −3.70 ± 0.50 | −8.82 ± 0.73 | −6.40 ± 0.62 |
| Pinus nigra | −4.33 ± 0.12 | −3.06 ± 0.35 | −5.60 ± 0.22 | −2.97 ± 0.22 |
| Pseudotsuga menziesii | −5.14 ± 0.84 | −3.47 ± 0.53 | −6.82 ± 1.43 | −5.28 ± 0.83 |
| Taxodium distichum | −2.30 ± 0.31 | −0.71 ± 0.58 | −4.19 ± 0.55 | −2.28 ± 0.44 |
| P50 | P12 | P88 | P25W | BD | br | tr | (tr/br)2 | bt | tt | (tt/bt)2 | b | t | (t/b)2 | CF/mm | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P50 | 0.91 ** | 0.98 *** | 0.95 ** | −0.92 * | 0.53 | −0.54 | −0.74 | 0.88 * | −0.44 | −0.86 * | 0.71 | −0.50 | −0.81 * | −0.85 * | |
| P12 | 0.86 *** | 0.80 | 0.81 * | −0.78 | 0.66 | −0.21 | −0.57 | 1.00 *** | −0.05 | −0.58 | 0.84 * | −0.13 | −0.60 | −0.93 ** | |
| P88 | 0.95 *** | 0.68 *** | 0.96 ** | −0.92 ** | 0.44 | −0.69 | −0.79 | 0.76 | −0.62 | −0.94 ** | 0.60 | −0.66 | −0.88 * | −0.75 | |
| P25W | 0.95 *** | 0.78 *** | 0.93 *** | −0.83 * | 0.29 | −0.59 | −0.64 | 0.76 | −0.54 | −0.86 * | 0.50 | −0.57 | −0.76 | −0.78 | |
| BD | −0.67 *** | −0.41 * | −0.74 *** | −0.63 *** | −0.50 | 0.50 | 0.74 | −0.77 | 0.44 | 0.83 * | −0.65 | 0.47 | 0.80 | 0.74 | |
| br | 0.47 ** | 0.38 * | 0.47 ** | 0.31 | −0.47 ** | −0.25 | −0.72 | 0.68 | 0.00 | −0.38 | 0.96 * | −0.13 | −0.60 | −0.66 | |
| tr | −0.38 * | −0.15 | −0.49 ** | −0.38 * | 0.30 | −0.28 | 0.82 * | −0.14 | 0.96 ** | 0.88 * | −0.23 | 0.99 ** | 0.87 ** | 0.26 | |
| (tr/br)2 | −0.51 *** | −0.32 | −0.58 *** | −0.39 ** | 0.48 *** | −0.82 *** | 0.76 *** | −0.54 | 0.66 | 0.87 * | −0.71 | 0.75 | 0.98 ** | 0.65 | |
| bt | 0.64 *** | 0.59 *** | 0.59 *** | 0.60 *** | −0.48 ** | 0.52 ** | −0.01 | −0.33 | 0.02 | −0.53 | 0.86 * | −0.06 | −0.55 | −0.93 ** | |
| tt | −0.22 | 0.07 | −0.38 | −0.27 | 0.21 | −0.08 | 0.89 *** | 0.57 *** | 0.04 | 0.83 * | 0.01 | 0.99 ** | 0.75 | 0.09 | |
| (tt/bt)2 | −0.54 *** | −0.29 | −0.63 *** | −0.55 *** | 0.44 ** | −0.34 * | 0.69 *** | 0.63 *** | −0.60 *** | 0.76 *** | −0.47 | 0.87 * | 0.95 * | 0.59 | |
| b | 0.61 *** | 0.52 *** | 0.59 *** | 0.49 ** | −0.54 *** | 0.92 *** | −0.20 | −0.72 *** | 0.80 *** | −0.05 | −0.51 ** | −0.12 | −0.63 | −0.82 ** | |
| t | −0.31 | −0.04 | −0.45 | −0.33 | 0.26 | −0.19 | 0.97 *** | 0.69 *** | 0.01 | 0.97 *** | 0.75 *** | −0.13 | 0.82 ** | 0.18 | |
| (t/b)2 | −0.59 *** | −0.35 * | −0.67 *** | −0.52 *** | 0.53 ** | −0.69 *** | 0.79 *** | 0.92 *** | −0.52 ** | 0.71 *** | 0.87 *** | −0.71 *** | 0.77 *** | 0.64 | |
| CF/mm | −0.46 ** | −0.42 * | −0.44 ** | −0.48 ** | 0.33 * | −0.46 ** | −0.06 | 0.23 | −0.84 *** | −0.03 | 0.46 *** | −0.69 *** | −0.05 | 0.38 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosner, S.; Nöbauer, S.; Voggeneder, K. Ready for Screening: Fast Assessable Hydraulic and Anatomical Proxies for Vulnerability to Cavitation of Young Conifer Sapwood. Forests 2021, 12, 1104. https://doi.org/10.3390/f12081104
Rosner S, Nöbauer S, Voggeneder K. Ready for Screening: Fast Assessable Hydraulic and Anatomical Proxies for Vulnerability to Cavitation of Young Conifer Sapwood. Forests. 2021; 12(8):1104. https://doi.org/10.3390/f12081104
Chicago/Turabian StyleRosner, Sabine, Sebastian Nöbauer, and Klara Voggeneder. 2021. "Ready for Screening: Fast Assessable Hydraulic and Anatomical Proxies for Vulnerability to Cavitation of Young Conifer Sapwood" Forests 12, no. 8: 1104. https://doi.org/10.3390/f12081104
APA StyleRosner, S., Nöbauer, S., & Voggeneder, K. (2021). Ready for Screening: Fast Assessable Hydraulic and Anatomical Proxies for Vulnerability to Cavitation of Young Conifer Sapwood. Forests, 12(8), 1104. https://doi.org/10.3390/f12081104