
Resilience of a Fire-Maintained Pinus palustris Woodland to Catastrophic Wind Disturbance: 10 Year Results



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Field Methods
2.3. Analytical Methods
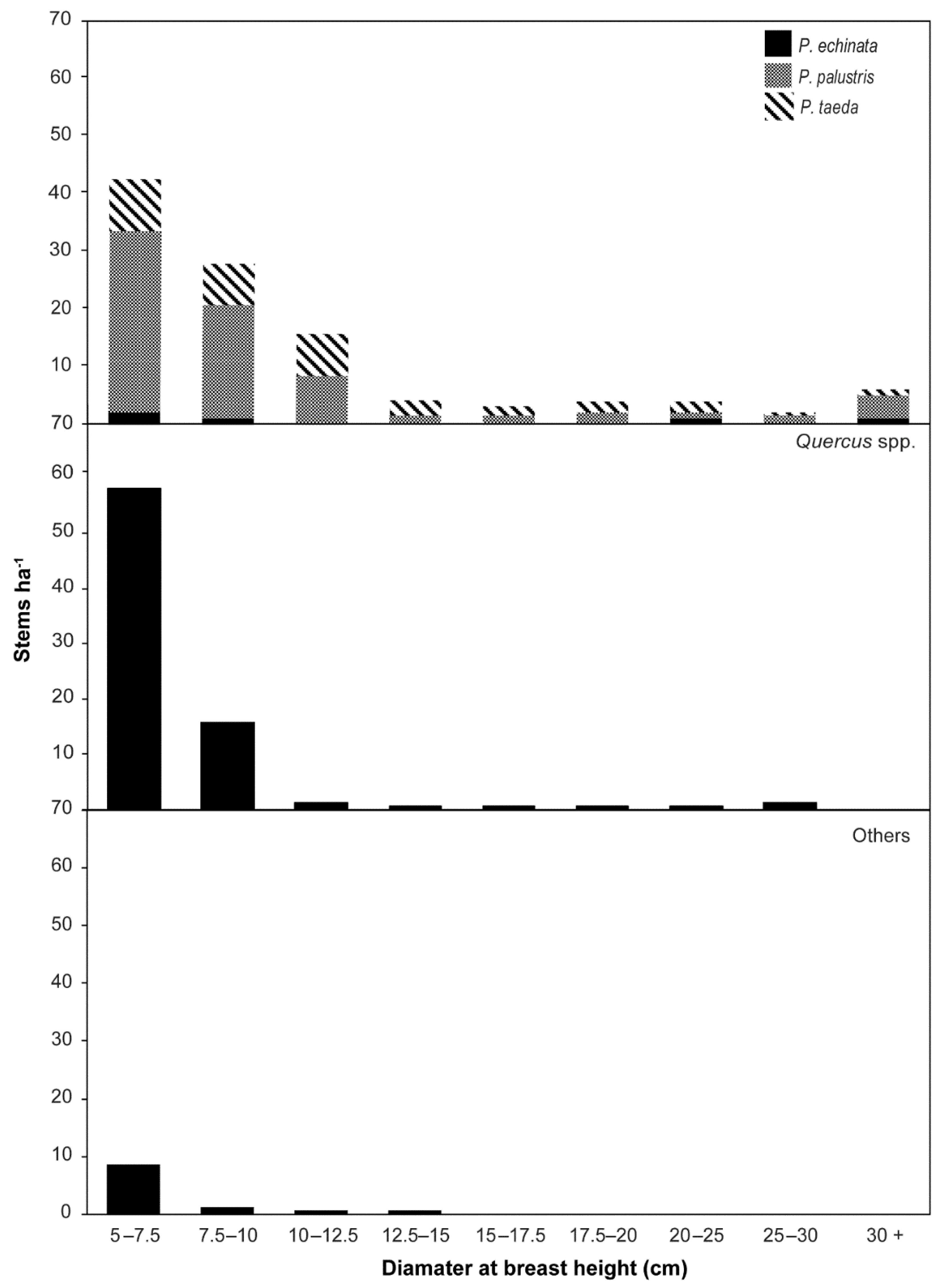
3. Results
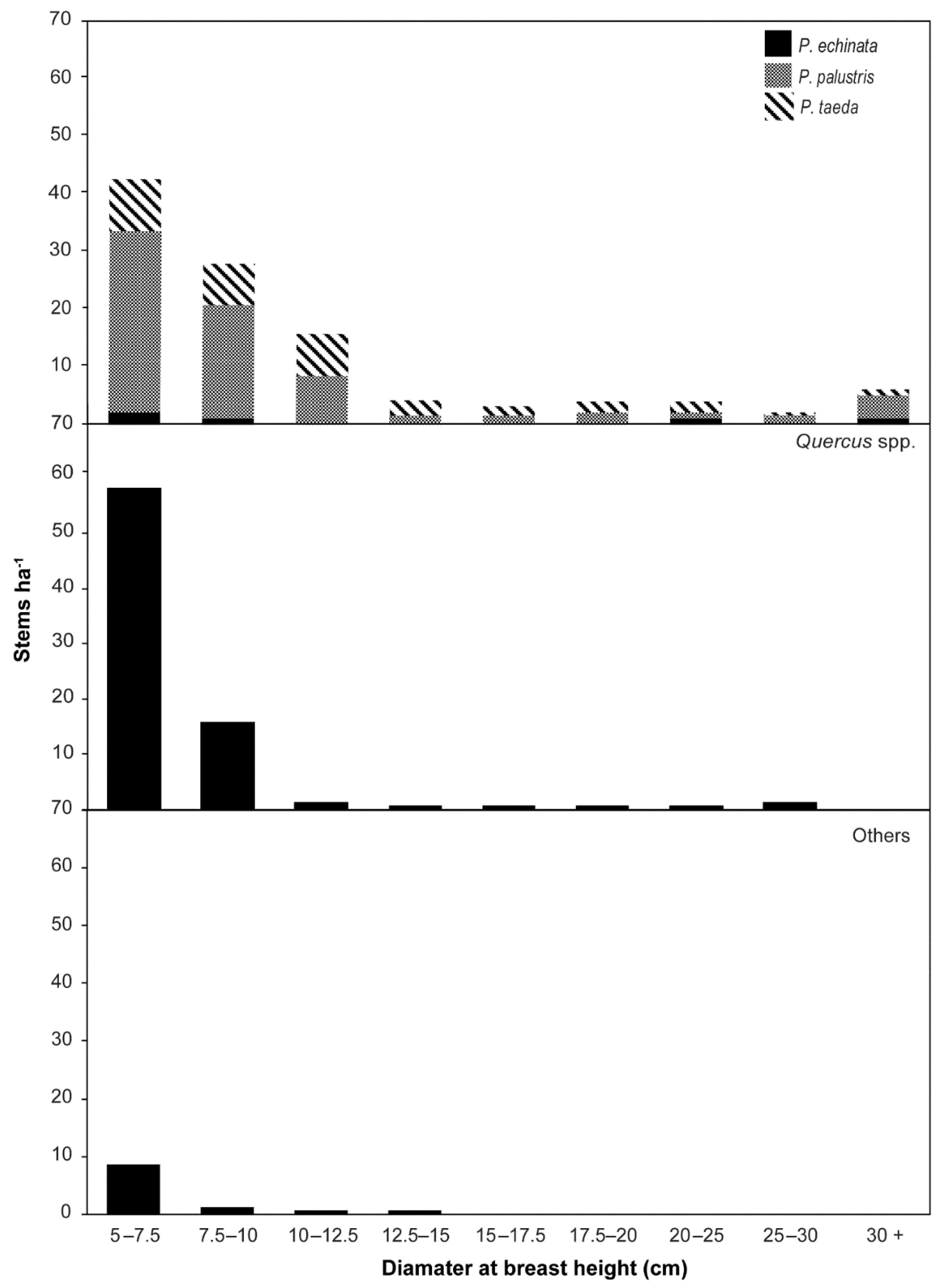
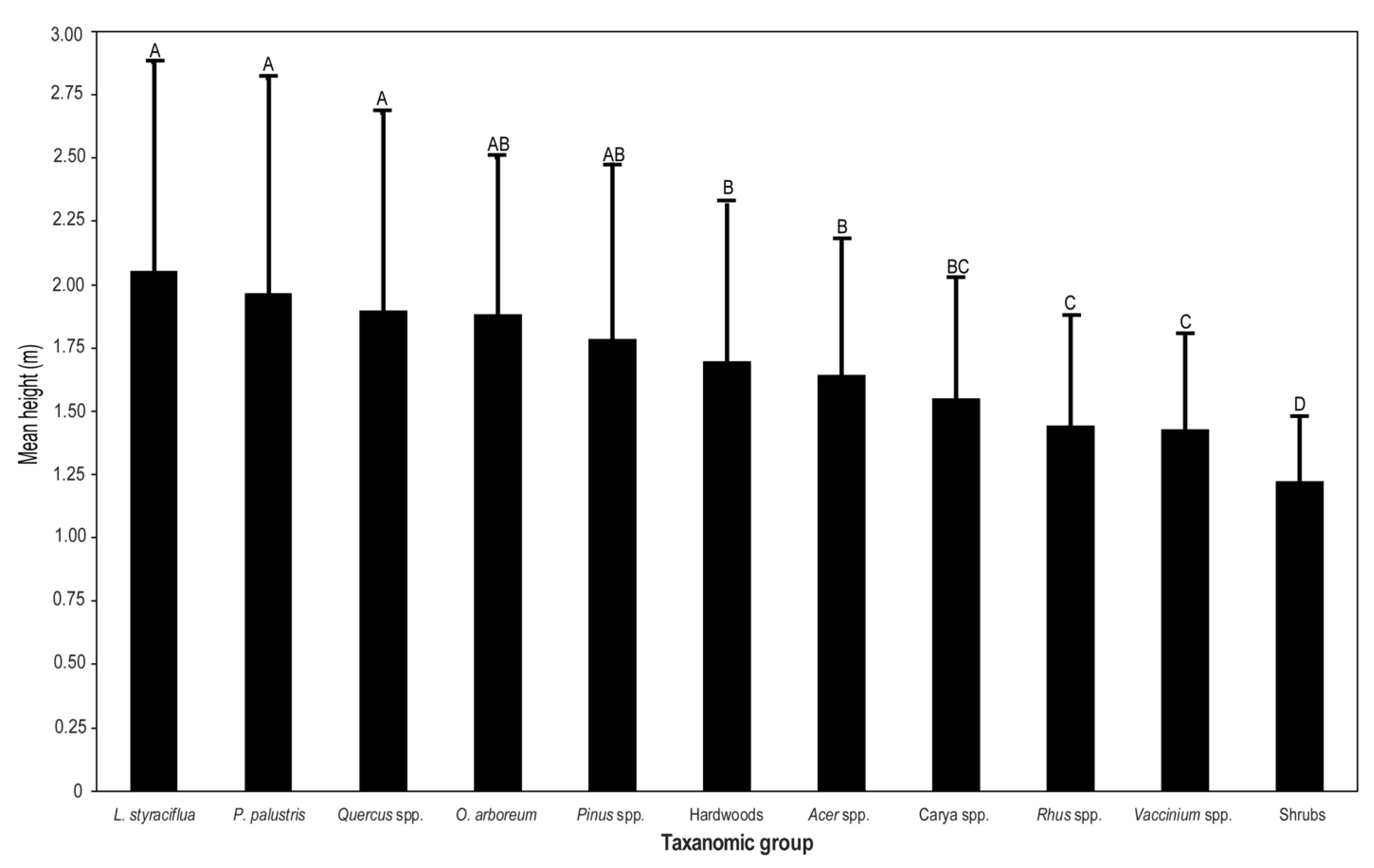
3.1. Woody Plant Composition, Structure, and Height
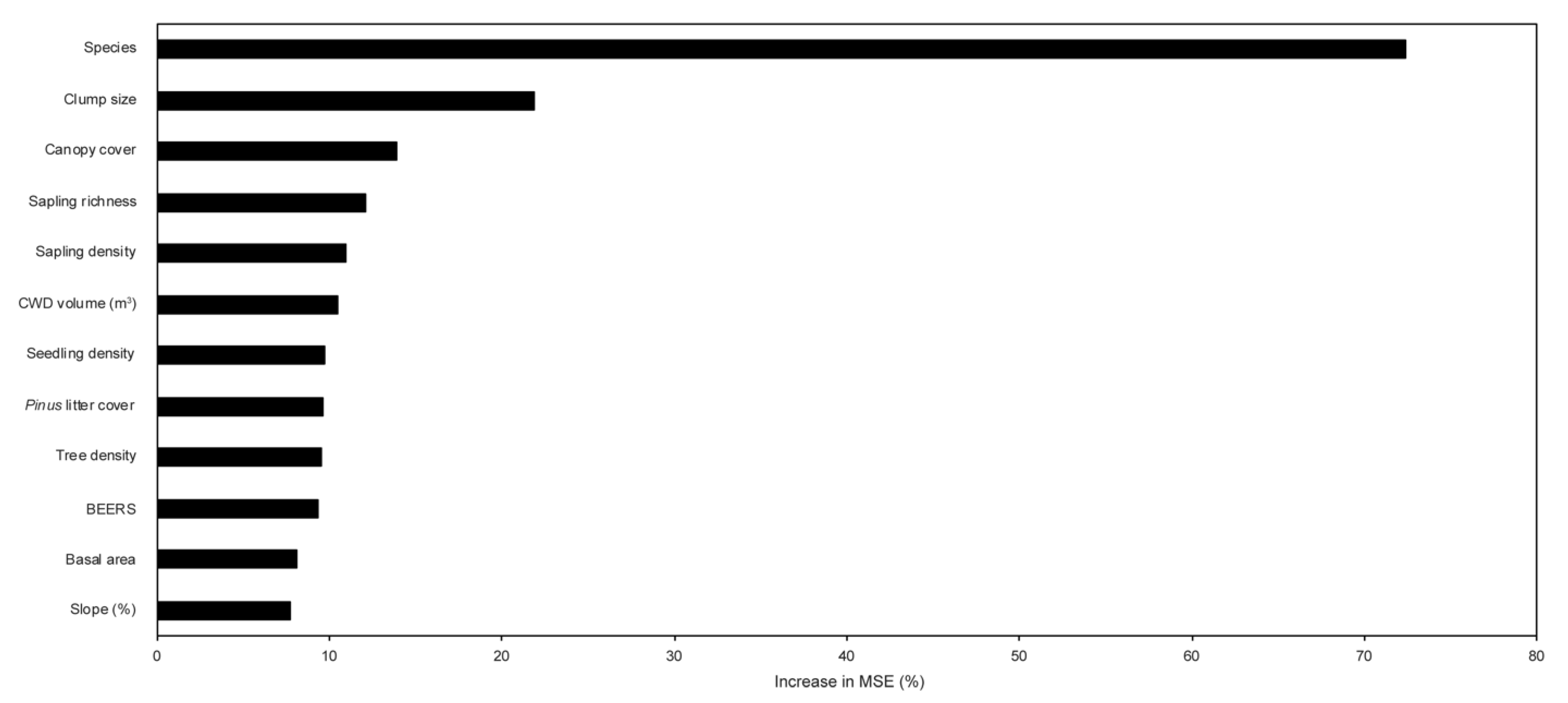
3.2. Biotic and Abiotic Influence on Sapling Height
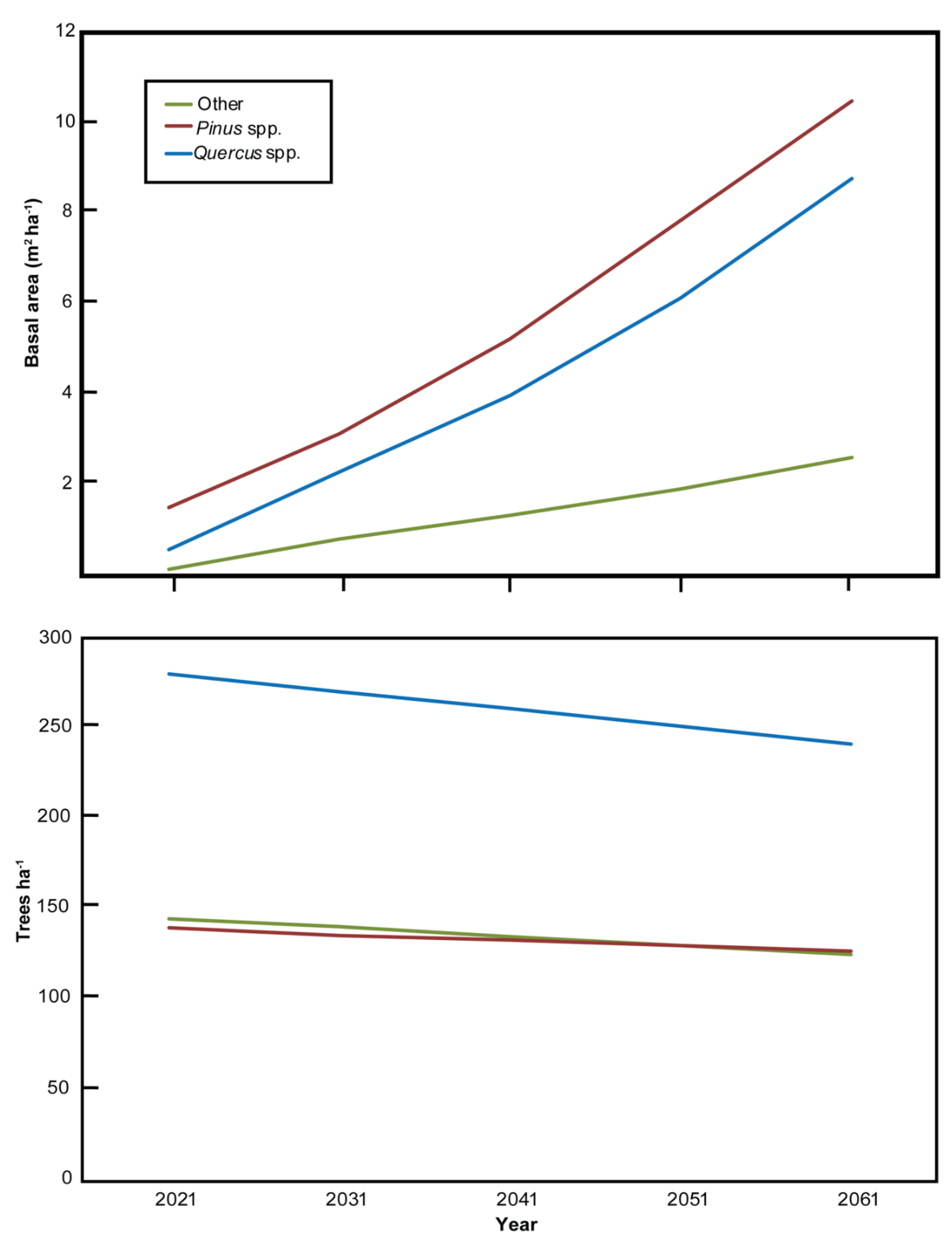
3.3. Stand Successional Trajectory
4. Discussion
4.1. Current Stand Conditions
4.2. Projected Stand Conditions
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, P.S.; Jentsch, A. The search for generality in studies of disturbance and ecosystem dynamics. Prog. Bot. 2001, 62, 399–450. [Google Scholar]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; John Wiley and sons: Hoboken, NJ, USA, 1996. [Google Scholar]
- Foster, D.R.; Knight, D.H.; Franklin, J.F. Landscape patterns and legacies resulting from large, infrequent forest disturbances. Ecosystems 1998, 1, 497–510. [Google Scholar] [CrossRef]
- Webster, C.R.; Dickinson, Y.L.; Burton, J.I.; Frelich, L.E.; Jenkins, M.A.; Kern, C.C.; Raymond, P.; Saunders, M.R.; Walters, M.B.; Willis, J.L. Promoting and maintaining diversity in contemporary hardwood forests: Confronting contemporary drivers of change and the loss of ecological memory. For. Ecol. Manag. 2018, 421, 98–108. [Google Scholar] [CrossRef]
- Johnstone, J.F.; Allen, C.D.; Franklin, J.F.; Frelich, L.E.; Harvey, B.J.; Higuera, P.E.; Mack, M.C.; Meentemeyer, R.K.; Metz, M.R.; Perry, G.L.; et al. Changing disturbance regimes, ecological memory, and forest resilience. Front. Ecol. Environ. 2016, 14, 369–378. [Google Scholar] [CrossRef]
- Franklin, J.F.; Lindenmayer, D.; MacMahon, J.A.; McKee, A.; Magnuson, J.; Perry, D.A.; Waide, R.; Foster, D. Threads of continuity. Conserv. Biol. 2000, 1, 8–16. [Google Scholar] [CrossRef]
- Palik, B.J.; D’Amato, A.W.; Franklin, J.F.; Johnson, K.N. Ecological Silviculture: Foundations and Applications; Waveland Press: Long Grove, IL, USA, 2020. [Google Scholar]
- Holling, C.S. Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Angeler, D.G.; Allen, C.R. Quantifying resilience. J. Appl. Ecol. 2016, 53, 617–624. [Google Scholar] [CrossRef]
- Myers, R.K.; van Lear, D.H. Hurricane-fire interactions in coastal forests of the south: A review and hypothesis. For. Ecol. Manag. 1998, 103, 265–276. [Google Scholar] [CrossRef]
- Busing, R.T.; White, R.D.; Harmon, M.E.; White, P.S. Hurricane disturbance in a temperate deciduous forest: Patch dynamics, tree mortality, and coarse woody detritus. In Forest Ecology; Springer: Dordrecht, The Netherlands, 2008; pp. 351–363. [Google Scholar]
- Cannon, J.B.; O’Brien, J.J.; Loudermilk, E.L.; Dickinson, M.B.; Peterson, C.J. The influence of experimental wind disturbance on forest fuels and fire characteristics. For. Ecol. Manag. 2014, 330, 294–303. [Google Scholar] [CrossRef]
- Mitchell, R.J.; Hiers, J.K.; O’Brien, J.; Starr, G. Ecological forestry in the Southeast: Understanding the ecology of fuels. J. For. 2009, 107, 391–397. [Google Scholar]
- Tepley, A.J.; Thomann, E.; Veblen, T.T.; Perry, G.L.; Holz, A.; Paritsis, J.; Anderson-Teixeira, K.J. Influences of fire–vegetation feedbacks and post-fire recovery rates on forest landscape vulnerability to altered fire regimes. J. Ecol. 2018, 106, 1925–1940. [Google Scholar] [CrossRef] [Green Version]
- Holzmueller, E.J.; Jose, S. Response of the invasive grass Imperata cylindrica to disturbance in the southeastern forests, USA. Forests 2012, 3, 853–863. [Google Scholar] [CrossRef] [Green Version]
- Cannon, J.B.; Peterson, C.J.; O’Brien, J.J.; Brewer, J.S. A review and classification of interactions between forest disturbance from wind and fire. For. Ecol. Manag. 2017, 406, 381–390. [Google Scholar] [CrossRef]
- Emery, R.K.; Kleinman, J.S.; Goode, J.D.; Hart, J.L. Effects of catastrophic wind disturbance, salvage logging, and prescribed fire on fuel loading and composition in a Pinus palustris woodland. For. Ecol. Manag. 2020, 478, 118515. [Google Scholar] [CrossRef]
- O’Brien, J.J.; Hiers, J.K.; Callaham, M.A., Jr.; Mitchell, R.J.; Jack, S. Interactions among overstory structure, seedling life history traits and fire in frequently burned neotropical pine forests. Ambio 2008, 37, 542–547. [Google Scholar] [CrossRef]
- Gensini, V.A.; Brooks, H.E. Spatial trends in United States tornado frequency. NPJ Clim. Atmos. Sci. 2018, 1, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Griffith, G.E.; Omernik, J.M.; Comstock, J.A.; Lawrence, S.; Martin, G.; Goddard, A.; Hulcher, V.J.; Foster, T. Ecoregions of Alabama and Georgia, (Color Poster with Map, Descriptive Text, Summary Tables, and Photographs); U.S Geological Survey: Reston, VA, USA, 2001. [Google Scholar]
- Braun, E.L. Deciduous Forests of Eastern North America; The Blackburn Press: Caldwell, NJ, USA, 1950. [Google Scholar]
- Fenneman, N.M. Physiography of Eastern United States; McGraw-Hill: New York, NY, USA, 1938. [Google Scholar]
- Shankman, D.; Hart, J.L. The fall line: A physiographic forest-vegetation boundary. Geogr. Rev. 2007, 97, 502–519. [Google Scholar] [CrossRef]
- Kleinman, J.S.; Hart, J.L. Vascular flora of longleaf pine woodlands after wind disturbance and salvage harvesting in the Alabama Fall Line Hills. Castanea 2018, 83, 183–195. [Google Scholar] [CrossRef]
- Harper, R.M. Forests of Alabama Geological Survey of Alabama; Monograph 10; Wetumpka Printing Company: Wetumpka, AL, USA, 1943. [Google Scholar]
- Beckett, S.; Golden, M.S. Forest vegetation and vascular flora of reed brake research natural area, Alabama. Castanea 1982, 47, 368–392. [Google Scholar]
- Cox, L.E.; Hart, J.L. Two centuries of forest compositional and structural changes in the Alabama fall line hills. Am. Midl. Nat. 2015, 174, 218–238. [Google Scholar] [CrossRef]
- Szabo, M.W.; Osborne, W.E.; Copeland, C.W., Jr.; Neathery, T.L. Geologic Map of Alabama (1:250,000): Alabama Geological Survey Special Map 220; Geological Survey of Alabama: Uscaloosa, AL, USA, 1988. [Google Scholar]
- USDA NRCS (United States Department of Agriculture, Natural Resources Conservation Service). Web Soil Survey. Available online: https://websoilsurvey.nrcs.usda.gov/ (accessed on 5 June 2021).
- United Stated Department of Agriculture, Natural Resources Conservation Service. Soil Survey of Bibb County, Alabama; 2008. Available online: http://soils.usda.gov/survey/printed_surveys/ (accessed on 5 June 2021).
- Boyer, W.D.; Bledsoe, B.W. Establishment Report: Reed Brake Research Natural Area, National Forest in Alabama, Bibb County, Alabama; USDA Forest Service Southeastern Forest Experiment Station: New Orleans, LA, USA, 1975.
- Thornthwaite, C.W. An approach toward a rational classification of climate. Geogr. Rev. 1948, 38, 55–94. [Google Scholar] [CrossRef]
- PRISM Climate Group. Northwest Alliance for Computational Science and Engineering. Available online: http://www.prism.oregonstate.edu/ (accessed on 8 June 2021).
- NWS (National Weather Service). Sawyerville-Eoline (Greene, Hale and Bibb Counties) EF-3 Tornado 27 April 2011. Available online: https://www.weather.gov/bmx/event_04272011sawyerville (accessed on 25 April 2020).
- Kleinman, J.S.; Ford, S.A.; Hart, J.L. Catastrophic wind and salvage harvesting effects on woodland plants. For. Ecol. Manag. 2017, 403, 112–125. [Google Scholar] [CrossRef]
- Ford, S.A.; Kleinman, J.S.; Hart, J.L. Effects of wind disturbance and salvage harvesting on macrofungal communities in a Pinus woodland. For. Ecol. Manag. 2018, 407, 31–46. [Google Scholar] [CrossRef]
- Kleinman, J.S.; Goode, J.D.; Hart, J.L.; Dey, D.C. Prescribed fire effects on Pinus palustris woodland development after catastrophic wind disturbance and salvage logging. For. Ecol. Manag. 2020, 468, 118173. [Google Scholar] [CrossRef]
- Goode, J.D.; Kleinman, J.S.; Hart, J.L.; Bhuta, A.A. Edge influence on composition and structure of a Pinus palustris woodland following catastrophic wind disturbance. Can. J. For. Res. 2020, 50, 332–341. [Google Scholar] [CrossRef]
- USDA, Forest Service. Forest Inventory and Analysis National Core Field Guide, Volume 1: Field Data Collection Procedures for Phase 2 Plots; Version 7.1; 2016; Available online: https://www.nrs.fs.fed.us/fia/data-collection/field-guides/ver7.1/FG%20NRS%207.1-Complete-Document.pdf (accessed on 6 July 2021).
- Lemmon, P.E. A new instrument for measuring forest overstory density. J. For. 1957, 55, 667–668. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Cutler, D.R.; Edwards, T.C., Jr.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef]
- Beers, T.W.; Dress, P.E.; Wensel, L.C. Aspect transformation in site productivity research. J. For. 1966, 64, 691–692. [Google Scholar]
- Dixon, G.E. Essential FVS: A User’s Guide to the Forest Vegetation Simulator; U.S. Department of Agriculture, F.S. Forest Management Service Center: Fort Collins, CO, USA, 2010.
- Scholl, E.R.; Waldrop, T.A. Photos for Estimating Fuel Loadings before and after Prescribed Burning in the Upper Coastal Plain of the Southeast; Gen. Tech. Rep. SRS-26; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 1999.
- Goode, J.D.; Brager, L.M.; Hart, J.L. Drought-induced growth response of longleaf pine in the Alabama Fall Line Hills. Southeast. Nat. 2019, 18, 99–112. [Google Scholar] [CrossRef]
- Wahlenberg, W.G. Longleaf Pine: Its Use, Ecology, Regeneration, Protection, Growth, and Management; Charles Lathrop Pack Forestry: Washington, DC, USA; Forest Service, U.S. Department of Agriculture: Washington DC, USA, 1976; Volume 429. [Google Scholar]
- Boyer, W.D. Pinus palustris Mill. longleaf pine. Silv. N. Am. 1990, 1, 405–412. [Google Scholar]
- Kirkman, L.K.; Jack, S.B. Ecological Restoration and Management of Longleaf Pine Forests; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Logan, A.T.; Goode, J.D.; Keellings, D.J.; Hart, J.L. Microsite influence on woody plant regeneration in a Pinus palustris woodland following catastrophic disturbance. Forests 2020, 11, 588. [Google Scholar] [CrossRef]
- Williamson, G.B.; Black, E.M. High temperature of forest fires under pines as a selective advantage over oaks. Nature 1981, 293, 643–644. [Google Scholar] [CrossRef]
- Provencher, L.; Herring, B.J.; Gordon, D.R.; Rodgers, H.L.; Tanner, G.W.; Hardesty, J.L.; Brennand, L.A.; Litt, A.R. Longleaf pine and oak responses to hardwood reduction techniques in fire-suppressed sandhills in northwest Florida. For. Ecol. Manag. 2001, 148, 63–77. [Google Scholar] [CrossRef]
- Wang, G.G.; Pile, L.S.; Knapp, B.O.; Hu, H. Longleaf pine adaptation to fire: Is early height growth pattern critical to fire survival? In Proceedings of the 18th Biennial Southern Silvicultural Research Conference, Knoxville, TN, USA, 2–5 March 2015; Knoxville, TN, USA 2–5 March 2015; e-Gen. Tech. Rep. SRS-212; US Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2016; Volume 12, pp. 214–218. [Google Scholar]
- Kabrick, J.M.; Clark, K.L.; D’Amato, A.W.; Dey, D.C.; Kenefic, L.S.; Kern, C.C.; Knapp, B.O.; Maclean, D.A.; Raymond, P.; Waskiewicz, J.D. Managing hardwood-softwood mixtures for future forests in eastern North America: Assessing suitability to projected climate change. J. For. 2017, 115, 190–201. [Google Scholar] [CrossRef] [Green Version]
- Phillips, D.L. Stand Dynamics in a Longleaf Pine Woodland: A Spatial Analysis. Master’s Thesis, University of Alabama, Tuscaloosa, AL, USA, 2021. [Google Scholar]
- Dey, D.C.; Hartman, G. Returning fire to Ozark Highland forest ecosystems: Effects on advance regeneration. For. Ecol. Manag. 2005, 217, 37–53. [Google Scholar] [CrossRef]
- Short, M.F.; Stambaugh, M.C.; Dey, D.C. Prescribed fire effects on oak woodland advance regeneration at the prairie–forest border in Kansas, USA. Can. J. For. Res. 2019, 49, 1570–1579. [Google Scholar] [CrossRef]
- Peters, D.P.; Lugo, A.E.; Chapin, F.S., III; Pickett, S.T.; Duniway, M.; Rocha, A.V.; Swanso, S.J.; Laney, C.; Jones, J. Cross-system comparisons elucidate disturbance complexities and generalities. Ecosphere 2011, 2, 1–26. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 5–7.5 | 7.5–10 | 10–12.5 | 12.5–14.5 | 15–17.5 | 17.5–20 | 20–24.5 | 25–30 | 30+ | Total | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | Density | Rel. Den. | Density | Rel. Den. | Density | Rel. Den. | Density | Rel. Den. | Density | Rel. Den. | Density | Rel. Den. | Density | Rel. Den. | Density | Rel. Den. | Density | Rel. Den. | Density | Rel. Den. |
| Pinus palustris Mill. | 30 | 26 | 21 | 52 | 9 | 50 | 1 | 13 | 1 | 25 | 1 | 40 | 1 | 20 | - | - | 3 | 100 | 65 | 34 |
| Pinus taeda L. | 11 | 9 | 6 | 14 | 7 | 39 | 3 | 63 | 1 | 50 | 1 | 40 | 1 | 40 | 1 | 50 | 1 | 25 | 32 | 17 |
| Quercus stellata Wangenh. | 18 | 16 | 3 | 8 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 21 | 11 |
| Quercus laevis Walter | 14 | 13 | 3 | 6 | 1 | 4 | - | - | - | - | - | - | - | - | 1 | 25 | - | - | 18 | 9 |
| Quercus coccinea Munchh. | 9 | 8 | 2 | 5 | 1 | 4 | - | - | - | - | - | - | - | - | - | - | - | - | 12 | 6 |
| Quercus marilandica Munchh. | 7 | 6 | 1 | 3 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 8 | 4 |
| Quercus nigra L. | 5 | 4 | 1 | 3 | - | - | - | - | 1 | 25 | 1 | 20 | 1 | 20 | - | - | - | - | 8 | 4 |
| Liquidambar styraciflua L. | 5 | 4 | - | - | 1 | 4 | - | - | - | - | - | - | - | - | - | - | - | - | 6 | 3 |
| Quercus alba L. | 4 | 4 | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 25 | - | - | 5 | 3 |
| Quercus margaretta (Ashe) Small | 3 | 2 | 1 | 3 | - | - | 1 | 13 | - | - | - | - | - | - | - | - | - | - | 4 | 2 |
| Pinus echinata Mill. | 1 | 1 | 1 | 2 | - | - | - | - | - | - | - | - | 1 | 20 | - | - | 1 | 40 | 3 | 1 |
| Magnolia macrophylla Michx. | 2 | 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | 1 |
| Quercus velutina Lam. | 1 | 1 | 1 | 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | 1 |
| Vaccinium arboreum Marshall | 1 | 1 | 1 | 2 | - | - | 1 | 13 | - | - | - | - | - | - | - | - | - | - | 2 | 1 |
| Acer rubrum L. | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 0 |
| Carya glabra (Mill.) Sweet | - | - | 1 | 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 0 |
| Cornus florida L. | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 0 |
| Nyssa sylvatica Marshall | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 0 |
| Quercus falcata Michx. | 1 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 0 |
| Total | 114 | 100 | 40 | 100 | 18 | 100 | 5 | 100 | 3 | 100 | 3 | 100 | 3 | 100 | 3 | 100 | 4 | 165 | 191 | 100 |
| Species | Density (Stems ha−1) | Relative Density (%) | Mean Height | SD |
|---|---|---|---|---|
| Rhus copallinum L. | 1207.5 | 23.6 | 1.4 | 0.4 |
| Vaccinium arboreum Marshall | 1095.0 | 21.4 | 1.4 | 0.4 |
| Quercus falcata Michx. | 396.3 | 7.7 | 1.9 | 0.8 |
| Liquidambar styraciflua L. | 348.8 | 6.8 | 2.1 | 0.8 |
| Quercus nigra L. | 277.5 | 5.4 | 1.8 | 0.7 |
| Pinus taeda L. | 237.5 | 4.6 | 1.8 | 0.7 |
| Quercus coccinea Munchh. | 173.1 | 3.4 | 2.0 | 0.9 |
| Diospyros virginiana L. a | 161.9 | 3.2 | 1.7 | 0.6 |
| Quercus alba L. | 155.6 | 3.0 | 1.9 | 0.7 |
| Quercus velutina Lam. | 116.3 | 2.3 | 1.8 | 0.8 |
| Carya tomentosa (Lam.) Nutt. | 106.9 | 2.1 | 1.5 | 0.4 |
| Quercus marilandica Munchh. | 102.5 | 2.0 | 2.2 | 0.8 |
| Oxydendrum arboreum (L.) DC. | 85.0 | 1.7 | 1.9 | 0.6 |
| Quercus stellata Wangenh. | 79.4 | 1.6 | 1.9 | 0.8 |
| Pinus palustris Mill. | 73.8 | 1.4 | 2.0 | 0.9 |
| Acer rubrum L. | 71.3 | 1.4 | 1.6 | 0.5 |
| Styrax grandifolius Aiton b | 65.0 | 1.3 | 1.1 | 0.2 |
| Symplocos tinctoria (L.) L’Hér. b | 55.0 | 1.1 | 1.3 | 0.3 |
| Nyssa sylvatica Marshall a | 44.4 | 0.9 | 1.6 | 0.6 |
| Carya glabra (Mill.) Sweet | 35.0 | 0.7 | 1.6 | 0.5 |
| Quercus hemisphaerica W. Bartram ex Willld. | 33.1 | 0.6 | 1.4 | 0.5 |
| Quercia margaretta (Ashe) Small | 31.3 | 0.6 | 1.9 | 0.6 |
| Quercus laevis Walter | 28.1 | 0.5 | 2.3 | 0.9 |
| Quercus montana Willd. | 22.5 | 0.4 | 1.7 | 0.5 |
| Callicarpa americana L. | 20.0 | 0.4 | 1.2 | 0.2 |
| Rhus glabra L. | 16.9 | 0.3 | 1.4 | 0.2 |
| Vaccinium stamineum L. | 15.6 | 0.3 | 1.3 | 0.3 |
| Acer floridium (Chapm.) Pax | 13.8 | 0.3 | 1.6 | 0.5 |
| Sassafra albidum (Nutt.) Nees a | 13.8 | 0.3 | 1.4 | 0.5 |
| Quercus incana W. Bartram | 5.6 | 0.1 | 1.7 | 0.8 |
| Cornus florida L. a | 5.6 | 0.1 | 1.5 | 0.4 |
| Prunus umbellata Elliott a | 5.0 | 0.1 | 2.1 | 0.6 |
| Liriodendron tulipifera L. a | 5.0 | 0.1 | 1.7 | 0.5 |
| Magnolia macrophylla Michx. a | 2.5 | 0.0 | 2.7 | 1.0 |
| Pinus echinata Mill. | 1.9 | 0.0 | 2.1 | 0.9 |
| Vaccinium pallidum Aiton | 1.9 | 0.0 | 1.7 | 0.3 |
| Hypericum hypericoides (L.) Crantz b | 1.3 | 0.0 | 1.5 | 0.0 |
| Vaccinium elliottii Chapm. | 1.3 | 0.0 | 1.5 | 0.7 |
| Acer saccharum Marshall | 1.3 | 0.0 | 1.3 | 0.4 |
| Magnolia virginiana L. a | 0.6 | 0.0 | 2.5 | 0.0 |
| Ilex vomitoria Aiton | 0.6 | 0.0 | 2.0 | 0.0 |
| Asimina parviflora Michx. Dunal b | 0.6 | 0.0 | 1.6 | 0.0 |
| Aesculus pavia L. b | 0.6 | 0.0 | 1.5 | 0.0 |
| Quercus rubra L. | 0.6 | 0.0 | 1.3 | 0.0 |
| Aralia spinosa L. b | 0.6 | 0.0 | 1.2 | 0.0 |
| Castanea Pumila (L.) Mill. a | 0.6 | 0.0 | 1.2 | 0.0 |
| Ilex opaca Aiton a | 0.6 | 0.0 | 1.2 | 0.0 |
| Hamamelis virginiana L. b | 0.6 | 0.0 | 1.1 | 0.0 |
| TOTAL | 5119.4 | 100.0 | – | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goode, J.D.; Kleinman, J.S.; Hart, J.L. Resilience of a Fire-Maintained Pinus palustris Woodland to Catastrophic Wind Disturbance: 10 Year Results. Forests 2021, 12, 1051. https://doi.org/10.3390/f12081051
Goode JD, Kleinman JS, Hart JL. Resilience of a Fire-Maintained Pinus palustris Woodland to Catastrophic Wind Disturbance: 10 Year Results. Forests. 2021; 12(8):1051. https://doi.org/10.3390/f12081051
Chicago/Turabian StyleGoode, J. Davis, Jonathan S. Kleinman, and Justin L. Hart. 2021. "Resilience of a Fire-Maintained Pinus palustris Woodland to Catastrophic Wind Disturbance: 10 Year Results" Forests 12, no. 8: 1051. https://doi.org/10.3390/f12081051
APA StyleGoode, J. D., Kleinman, J. S., & Hart, J. L. (2021). Resilience of a Fire-Maintained Pinus palustris Woodland to Catastrophic Wind Disturbance: 10 Year Results. Forests, 12(8), 1051. https://doi.org/10.3390/f12081051