
Genetic Diversity and Structure of Pinus densiflora Siebold & Zucc. Populations in Republic of Korea Based on Microsatellite Markers


Abstract
1. Introduction
2. Materials and Methods
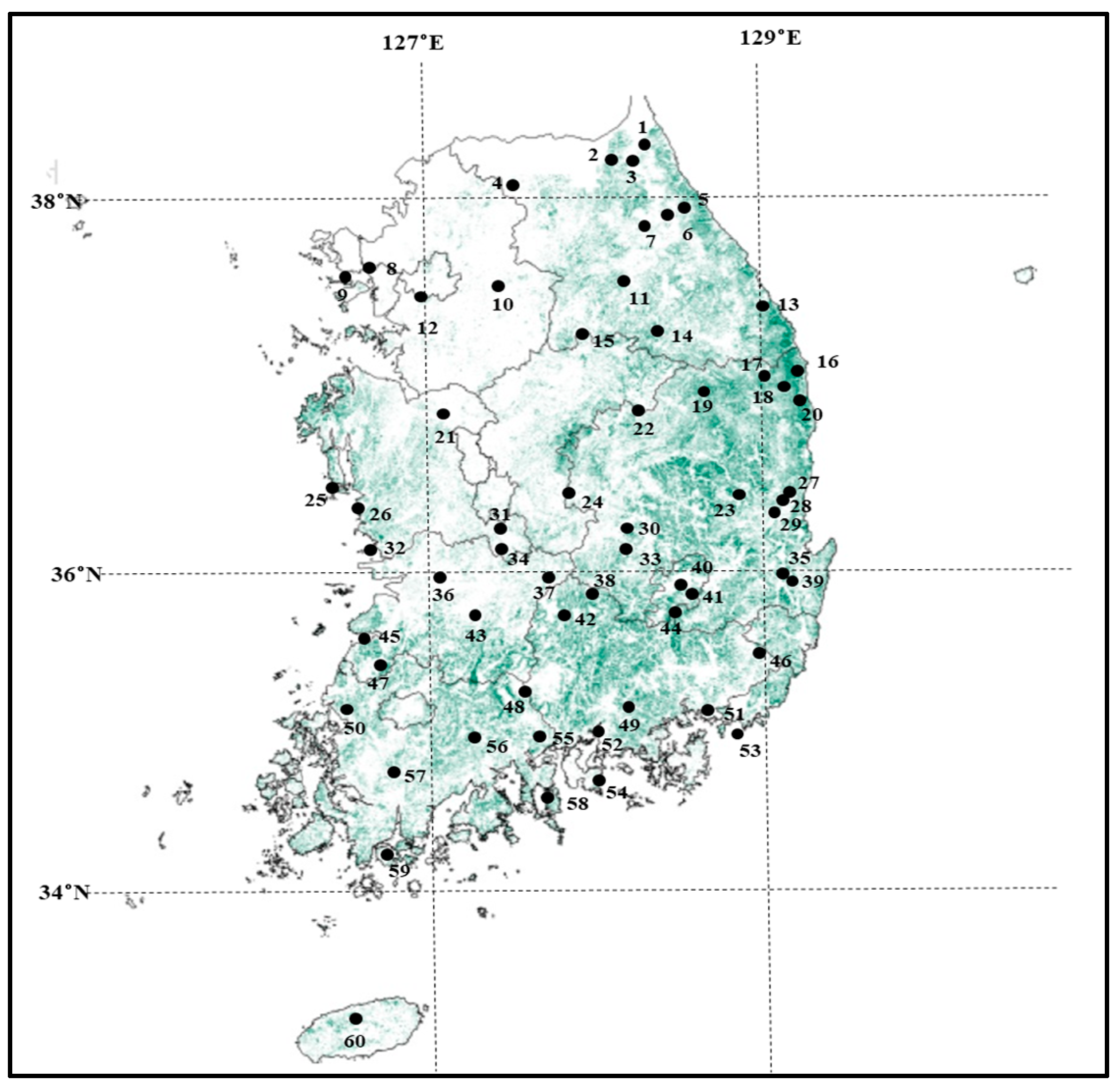
2.1. Sample Collection and DNA Extraction

2.2. Microsatellite Analysis
2.3. Data Analysis
3. Results
3.1. Genetic Diversity within a Population
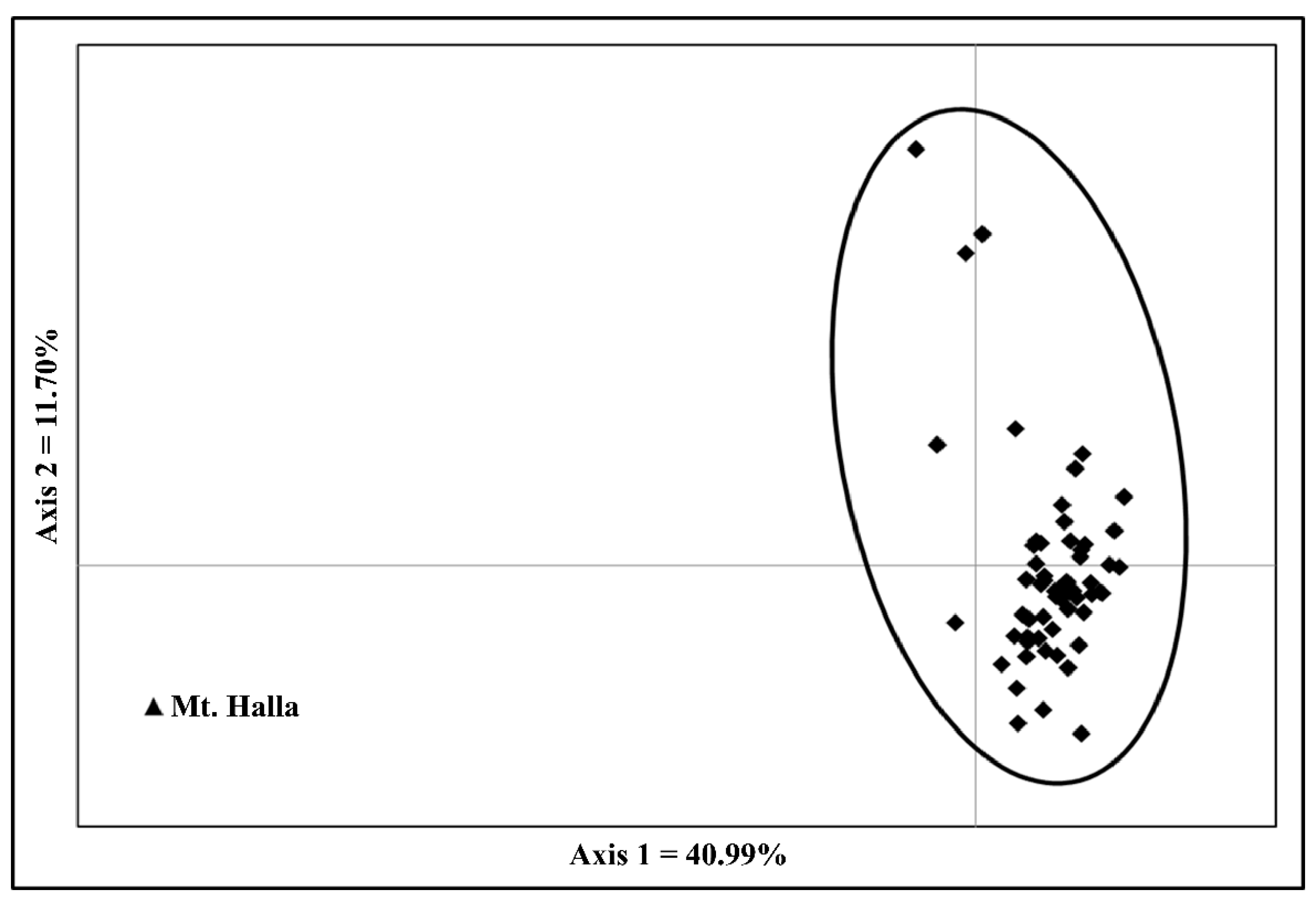
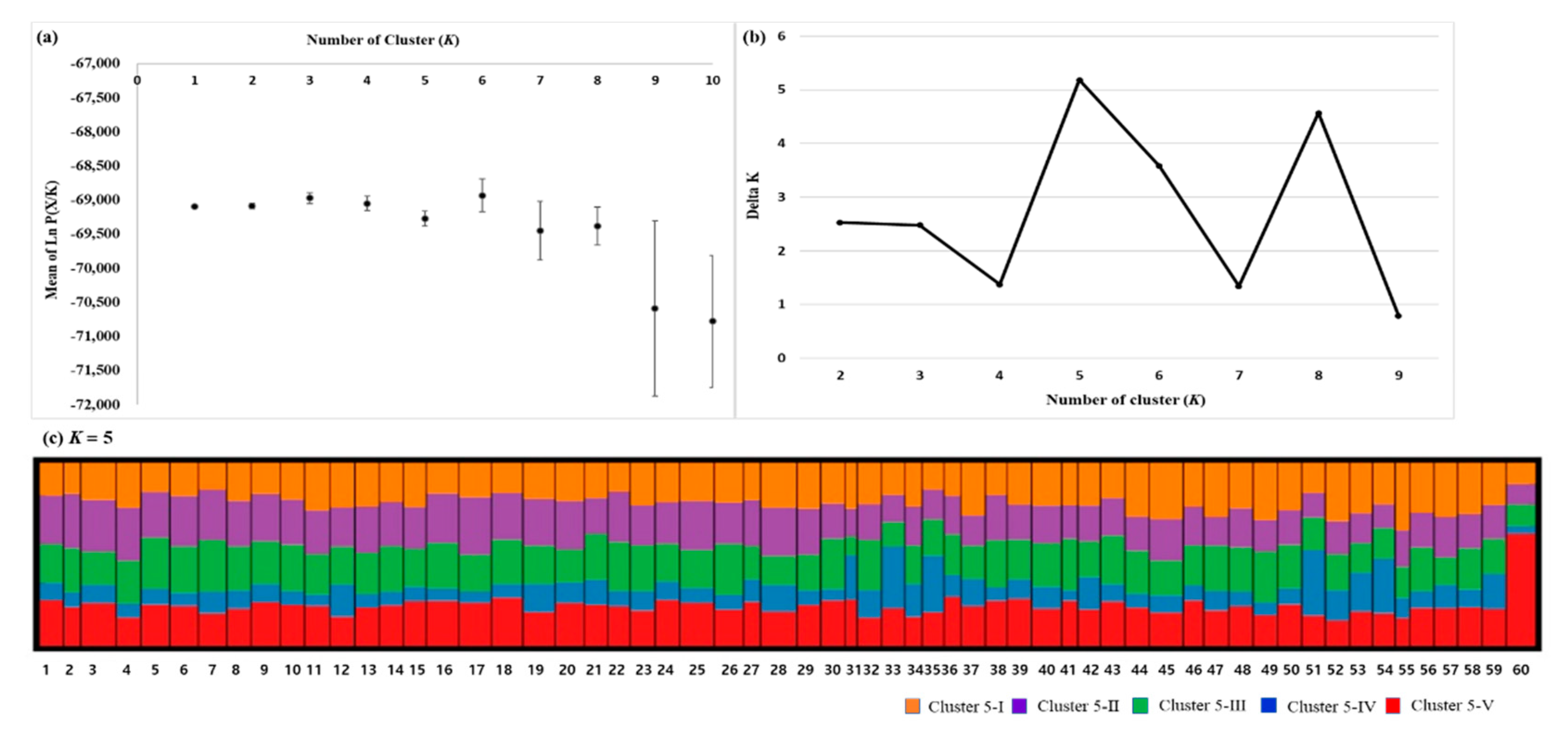
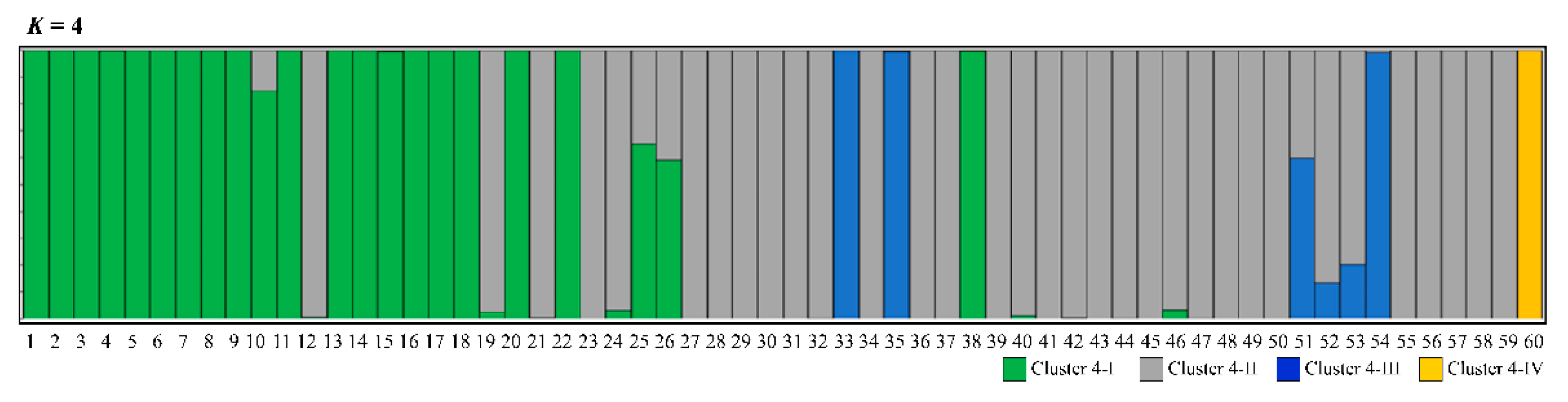
3.2. Genetic Differentiation and Population Structure
4. Discussion
4.1. Genetic Diversity within a Population
4.2. Genetic Differentiation among Populations
4.3. Population Structure
4.4. Implication for Forest Conservation and Management of P. densiflora
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Park, J.H. Chapter II Forest Resource-Forest area and growing stock by key tree species. In Statistical Yearbook of Forestry, 50th ed.; Lim, S.S., Ed.; Korea Forest Service: Dajeon, Korea, 2020; p. 162. [Google Scholar]
- Kong, W.S.; Lee, S.G.; Park, H.N.; Lee, Y.M.; Oh, S.H. Time-spatial distribution of Pinus in the Korean peninsula. Quat. Int. 2014, 344, 43–52. [Google Scholar] [CrossRef]
- IUCN Red list Pinus densiflora. Available online: https://dx.doi.org/10.2305/IUCN.UK.2013-1.RLTS.T42355A2974820.en (accessed on 15 February 2020).
- Szmidt, A.E.; Wang, X.R. Molecular systematics and genetic differentiation of Pinus sylvestris (L.) and P. densiflora (Sieb. et Zucc.). Theor. Appl. Genet. 1993, 86, 159–165. [Google Scholar] [CrossRef]
- Kim, Z.S. Science of Pinus densiflora: From DNA to Management; Korea University Press: Seoul, Korea, 2014; p. 404. [Google Scholar]
- Cho, H.J.; Lee, C.B. Vegetation types and diversity patterns of Pinus densiflora forests in South Korea. J. Korean For. Soc. 2011, 100, 118–123. [Google Scholar]
- Hong, Y.P.; Kwon, H.Y.; Kim, I.S. I-SSR markers revealed inconsistent phylogeographic patterns among populations of Japanese red pine in Korea. Silvae Genet. 2007, 56, 22–26. [Google Scholar] [CrossRef][Green Version]
- Kamada, M.; Nakagoshi, N. Pine forest structure in a human-dominated landscape system in Korea. Ecol. Res. 1993, 8, 35–46. [Google Scholar] [CrossRef]
- Bae, J.S. A study on the system and the development of Pine Policy in the Late Chosun era. Korean J. For. Econ. 2002, 10, 22–50. [Google Scholar]
- Kim, J.W. What is the problem about Pinus densiflora in Korea? Through Pine Wilt Disease (Bursaphelenchus xylophilus) and Wildfire of East coast. Korean J. Environ. Ecol. 2005, 28, 113–120. [Google Scholar] [CrossRef]
- Lee, S.Y.; Jun, K.W.; Lee, M.W.; Chun, K.W. Mortality in Pine stand and vegetation recovery after Forest Fire. Korean Soc. Hazard Mitig. 2008, 8, 71–79. [Google Scholar]
- Kim, E.S.; Lee, B.R.; Kim, J.B.; Cho, N.H.; Lim, J.H. Risk assessment of Pine Tree Dieback in Sogwang-Ri, Uljin. J. Korean For. Soc. 2020, 109, 259–270. [Google Scholar]
- Uyeki, H. On the Physiognomy of Pinus densiflora Growing in Korea and Silvicultural Treatment for its Improvement. In Bulletin of the Agricultural and Forestry College, Suigen, Chosen; Suwon College of Agriculture and Forestry: Suwon, Korea, 1928; p. 263. [Google Scholar]
- Lee, S.W.; Jang, S.S.; Jang, K.H.; Kim, C.S. Estimation of mating system parameters in the natural population of Pinus densiflora of Anmyun Island, Korea using allozyme markers. J. Korean For. Soc. 2003, 92, 121–128. [Google Scholar]
- Yim, K.B.; Kim, Z.S. The variation of natural population of Pinus densiflora S. et Z. in Korea (I)-Characteristics of needle and wood of Chuwang-san, An-Myeon-Do and Odae-san populations. J. Korean For. Soc. 1975, 28, 1–20. [Google Scholar]
- Yi, J.S.; Song, J.H.; Han, S.S.; Park, W.G. The variation of natural populations of Pinus densiflora S. et Z. in Kangwondo-The morphological characteristics of needle and cone. J. For. Environ. Sci. 1999, 15, 107–116. [Google Scholar]
- Lee, C.H.; Shin, C.H.; Kim, K.S. Geographic variation in needle characteristics of Pinus densiflora in Korea. Korean J. Plant. Res. 2007, 20, 336–341. [Google Scholar]
- Kim, I.S.; Ryu, K.O.; Song, J.H.; Kim, T.S. Geographic variation in survival rate and height growth of Pinus densiflora S. et Z. in Korea. J. Korean For. Soc. 2005, 94, 73–81. [Google Scholar]
- Kim, Z.S.; Lee, S.W. Genetic Diversity of Three Native Pinus Species in Korea. In Population Genetics and Genetic Conservation of Forest Trees; Baradat, P., Adams, W.T., Müller-Starck, G., Eds.; SPB Publishing: Amsterdam, The Netherland, 1994; pp. 211–218. [Google Scholar]
- Kim, Y.Y.; Hyun, J.O.; Hong, K.N.; Choi, T.B.; Kim, K.S. Genetic variation of natural populations of Pinus densiflora in Korea based on RAPD marker analysis. Korean J. Breed. Sci. 1995, 27, 23–48. [Google Scholar]
- Balloux, F.; Lugon-Moulin, N. The estimation of population differentiation with microsatellite markers. Mol. Ecol. 2002, 11, 155–165. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M.J.W.; Gonzales, E. Conservation of genetic diversity in old-growth forest communities of the southeastern United States. Appl. Veg. Sci. 2006, 9, 51–58. [Google Scholar] [CrossRef]
- Porth, I.; El-Kassaby, Y.A. Assessment of the genetic diversity in forest tree populations using molecular markers. Diversity 2014, 6, 283–295. [Google Scholar] [CrossRef]
- Liewlaksaneeyanawin, C.; Ritland, C.E.; El-Kassaby, Y.A.; Ritland, K. Single-copy, species-transferable microsatellite markers developed from loblolly pine ESTs. Theor. Appl. Genet. 2004, 109, 361–369. [Google Scholar] [CrossRef]
- Ahn, J.Y.; Lee, J.W.; Lim, H.Y.; Hong, K.N. Genetic diversity and structure of Prunus padus populations in South Korea based on AFLP markers. For. Sci. Technol. 2020, 16, 171–179. [Google Scholar] [CrossRef]
- Watanabe, A.; Iwaizumi, M.G.; Ubukata, M.; Kondo, T.; Lian, C.; Hogetsu, T. Isolation of microsatellite markers from Pinus densiflora Sieb. et Zucc. using a dual PCR technique. Mol. Ecol. Notes 2006, 6, 80–82. [Google Scholar] [CrossRef]
- Lian, C.; Miwa, M.; Hogetsu, T. Isolation and characterization of microsatellite loci from the Japanese red pine, Pinus densiflora. Mol. Ecol. 2000, 9, 1186–1188. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Lee, J.B.; Gil, J.S.; Um, Y.R.; Kim, J.H.; Hwang, M.Y.; Kim, H.B.; Hong, C.P.; Park, S.G.; Shim, D.H.; et al. Development of polymorphic SSR markers from Pinus densiflora (Pinaceae) natural population in Korea. Plant. Breed. Biotechnol. 2019, 7, 67–71. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P.F. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Marshall, T.C.; Slate, J.; Kruuk, L.; Pemberton, J.M. Statistical confidence for likelihood-based paternity inference in natural populations. Mol. Ecol. 1998, 6, 639–655. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENEALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT Program to Estimate and Test Gene Diversities and Fixation Indices (Version 2.9.3.2). Available online: http://www2.unil.ch/popgen/softwares/fstat.html (accessed on 30 June 2020).
- Piry, S.; Luikart, G.; Cornuet, J.M. BOTTLENECK: A program for detecting recent effective population size reductions from allele data frequencies. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Iwaizumi, M.G.; Tsuda, Y.; Ohtani, M.; Tsumura, Y.; Takahashi, M. Recent distribution changes affect geographic clines in genetic diversity and structure of Pinus densiflora natural populations in Japan. For. Ecol. Manag. 2013, 304, 407–416. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Reanaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, N.A. Distruct: A program for the graphical display of population structure. Mol. Ecol. Notes 2003, 4, 137–138. [Google Scholar] [CrossRef]
- Guillot, G.; Mortier, F.; Estoup, A. GENLAND: A computer package for landscape genetics. Mol. Ecol. Notes 2005, 5, 712–715. [Google Scholar] [CrossRef]
- Moriguchi, Y.; Kang, K.S.; Lee, K.Y.; Lee, S.W.; Kim, Y.Y. Genetic variation of Picea jezoensis populations in South Korea revealed by chloroplast, mitochondrial and nuclear DNA markers. J. Plant. Res. 2009, 122, 153–160. [Google Scholar] [CrossRef]
- Hong, Y.P.; Ahn, J.Y.; Kim, Y.M.; Yang, B.H.; Song, J.H. Genetic variation of nSSR markers in natural populations of Abies koreana and Abies nephrolepis in South Korea. J. Korean For. Soc. 2011, 100, 577–584. [Google Scholar]
- Nybom, H. Comparison of different nuclear DNA markers for estimating intraspecific genetic diversity in Plants. Mol. Ecol. 2004, 13, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, H.; Watanabe, A.; Uchiyama, K.; Saito, Y.; Ide, Y. Influence of long-distance seed dispersal on the genetic diversity of seed rain in fragmented Pinus densiflora populations relative to pollen-mediated gene flow. J. Hered. 2013, 104, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Lian, C.; Miwa, M.; Hogetsu, T. Outcrossing and paternity analysis of Pinus densiflora (Japanese red pine) by microsatellite polymorphism. Heredity 2001, 87, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Belletti, P.; Ferrazzini, D.; Piotti, A.; Monteleone, I.; Ducci, F. Genetic variation and divergence in Scots Pine (Pinus sylvestris L.) within its natural range in Italy. Eur. J. For. Res. 2012, 131, 1127–1138. [Google Scholar] [CrossRef]
- Tong, Y.W.; Lewis, B.J.; Zhou, W.M.; Mao, C.R.; Wang, Y.; Zhou, L.; Yu, D.P.; Dai, L.M.; Qi, L. Genetic diversity and population structure of natural Pinus koraiensis populations. Forests 2020, 11, 39. [Google Scholar] [CrossRef]
- Whitney, T.D.; Gandhi, K.J.K.; Hamrick, J.L.; Lucardi, R.D. Extant population genetic variation and structure of eastern white pine (Pinus strobus L.) in the Southern Appalachians. Tree Genet. Genomes 2019, 15, 1–19. [Google Scholar] [CrossRef]
- Tóth, E.G.; Tremblay, F.; Housset, J.M.; Bergeron, Y.; Carcaillet, C. Geographic isolation and climatic variability contribute to genetic differentiation in fragmented populations of the long-lived subalpine conifer Pinus cembra L. in the western Alps. BMC Evol. Biol. 2019, 19, 1–17. [Google Scholar] [CrossRef]
- Vogl, C.; Karhu, A.; Morgan, G.; Savolainen, O. High resolution analysis of mating systems: Inbreeding in natural populations of Pinus radiata. J. Evol. Biol. 2002, 15, 433–439. [Google Scholar] [CrossRef]
- Thomas, B.R.; Macdonald, S.E.; Hicks, M.; Adams, D.L.; Hodgetts, R.B. Effects of reforestation methods on genetic diversity of lodgepole pine: An assessment using microsatellite and randomly amplified polymorphic DNA markers. Theor. Appl. Genet. 1999, 98, 793–801. [Google Scholar] [CrossRef]
- Potter, K.M.; Jetton, R.M.; Dvorak, W.S.; Hipkins, V.D.; Rhea, R.; Whittier, W.A. Widespread inbreeding and unexpected geographic patterns of genetic variation in eastern hemlock (Tsuga canadensis), an imperiled North American conifer. Conserv. Genet. 2012, 13, 475–498. [Google Scholar] [CrossRef][Green Version]
- Han, S.D.; Hing, Y.P.; Yang, B.H.; Lee, S.W.; Kim, C.S. Estimation of mating system parameters in the natural population of Pinus densiflora of Mt. Juwang. Proc. J. Korean For. Soc. 2004, 315–316. [Google Scholar]
- Kim, Y.M.; Hong, K.N.; Park, Y.J.; Hong, Y.P.; Park, J.I. Estimating the parameters of pollen flow and mating system in Pinus densiflora population in Buan, South Korea, using microsatellite markers. Korean J. Plant. Res. 2015, 28, 101–110. [Google Scholar] [CrossRef][Green Version]
- Myking, T. Evaluating genetic resources of forest trees by means of life history traits-a Norwegian example. Biodivers. Conserv. 2002, 11, 1681–1696. [Google Scholar] [CrossRef]
- Iwaizumi, M.G.; Takahashi, M.; Watanabe, A.; Ubukata, M. Simultaneous evaluation of paternal and maternal immigrant gene flow and the implications for the overall genetic composition of Pinus densiflora dispersed seeds. J. Hered. 2010, 101, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Dolezal, J.; Altman, J.; Kopecky, M.; Cerny, T.; Janecek, S.; Bartos, M.; Petrik, P.; Srutek, M.; Leps, J.; Song, J.S. Plant diversity changes during the postglacial in East Asia: Insights from forest refugia on Halla volcano, Jeju Island. PLoS ONE 2012, 7, e33065. [Google Scholar]
- Karhu, A.; Vogl, C.; Moran, G.F.; Bell, J.C.; Savolainen, O. Analysis of microsatellite variation in Pinus radiata reveals effects of genetic drift but no recent bottlenecks. J. Evol. Biol. 2006, 19, 167–175. [Google Scholar] [CrossRef]
- Manel, S.; Schwartz, M.K.; Luikart, G.; Taberlet, P. Landscape genetics: Combining landscape ecology and population genetics. Trends Ecol. Evol. 2003, 18, 189–197. [Google Scholar] [CrossRef]
- Frantz, A.C.; Cellina, S.; Krier, A.; Schley, L.; Burke, T. Using spatial Bayesian methods to determine the genetic structure of a continuously distributed population: Clusters or isolation by distance? J. Appl. Ecol. 2009, 46, 493–505. [Google Scholar] [CrossRef]
- Sork, V.L.; Smouse, P.E. Genetic analysis of landscape connectivity in tree populations. Landsc. Ecol. 2006, 21, 821–836. [Google Scholar] [CrossRef]
- Che, S.H.; Kim, W. Comparison of Plant community structures in cut and uncut areas at burned area of Mt. Gemo-san. J. Korean For. Soc. 1997, 86, 509–520. [Google Scholar]
- Kim, H.S.; Park, G.S.; Lee, S.M.; Lee, J.K. Analysis on the Correlation Between Vegetation Structure and Environment Factors of the Geumosanseong-inside in Geumosan Provincial Park, in Korea. J. Korean Soc. Environ. Restor. Technol. 2020, 23, 49–67. [Google Scholar]
- Ahn, S.H.; Jeon, M.J.; Eom, Y.G.; Oh, S.C.; Lee, M.R. Wood Anatomical Characteristics of Domestic Red Pine (Pinus densiflora) Infested by Pine wood Nematode (Bursaphelenchus xylophilus). J. Korean Wood Sci. Technol. 2011, 39, 15–20. [Google Scholar] [CrossRef]
- Cho, N.H.; Kim, E.S.; Lee, B.R.; Lim, J.H.; Kang, S.K. Predicting the potential distribution of Pinus densiflora and analyzing the Relationship with Environmental variable using MaxEnt model. Korean J. Agric. For. Meteorol. 2020, 22, 47–56. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Population | Latitude (N°) | Longitude (E°) | N | A | Ae | AR | Ho | He | FIS |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Mt. Seorak | 38.13 | 128.29 | 30 | 8.5 | 4.3 | 6.9 | 0.612 | 0.661 | 0.091 *** |
| 2 | Yanggu | 38.09 | 128.03 | 21 | 7.7 | 4.1 | 6.7 | 0.707 | 0.685 | −0.007 |
| 3 | Inje | 38.06 | 128.34 | 44 | 9.3 | 4.4 | 6.7 | 0.621 | 0.689 | 0.111 *** |
| 4 | Mt. Hwaak | 38.03 | 127.52 | 30 | 9.4 | 4.2 | 7.0 | 0.654 | 0.672 | 0.044 |
| 5 | Gangneung | 37.83 | 128.72 | 36 | 9.5 | 4.7 | 7.0 | 0.664 | 0.684 | 0.044 |
| 6 | Mt. Odae | 37.81 | 128.57 | 35 | 8.3 | 4.3 | 6.4 | 0.645 | 0.659 | 0.037 |
| 7 | Hongcheon | 37.74 | 128.34 | 35 | 9.2 | 4.3 | 6.8 | 0.645 | 0.684 | 0.071 ** |
| 8 | Kimpo | 37.74 | 126.54 | 30 | 8.4 | 3.9 | 6.6 | 0.589 | 0.637 | 0.093 ** |
| 9 | Kanghwado | 37.63 | 126.42 | 36 | 9.0 | 4.7 | 6.9 | 0.675 | 0.681 | 0.024 |
| 10 | Yangpyeong | 37.55 | 127.58 | 30 | 9.5 | 4.4 | 7.0 | 0.675 | 0.661 | −0.004 |
| 11 | Hoengseong | 37.52 | 128.27 | 31 | 8.7 | 4.7 | 6.9 | 0.642 | 0.666 | 0.052 * |
| 12 | Mt. Gwanak | 37.43 | 126.97 | 31 | 9.2 | 4.5 | 7.0 | 0.712 | 0.676 | −0.037 |
| 13 | Samcheok | 37.29 | 129.32 | 30 | 8.5 | 4.2 | 6.6 | 0.636 | 0.645 | 0.031 |
| 14 | Pyeongchang | 37.28 | 128.56 | 30 | 8.6 | 4.9 | 6.9 | 0.660 | 0.672 | 0.035 |
| 15 | Wonju | 37.27 | 127.94 | 28 | 8.4 | 4.4 | 6.8 | 0.637 | 0.658 | 0.049 * |
| 16 | Ujin Deoguri | 37.07 | 129.28 | 40 | 9.4 | 4.6 | 6.7 | 0.664 | 0.678 | 0.033 |
| 17 | Bonghwa | 37.05 | 128.98 | 40 | 9.4 | 4.5 | 6.9 | 0.639 | 0.677 | 0.069 ** |
| 18 | Uljin Sokwangri | 37.01 | 129.20 | 39 | 9.3 | 4.6 | 6.8 | 0.643 | 0.671 | 0.055 ** |
| 19 | Youngju | 37.00 | 128.69 | 40 | 10.0 | 4.3 | 7.0 | 0.675 | 0.687 | 0.030 |
| 20 | Uljin Sugokri | 36.95 | 129.32 | 36 | 9.5 | 5.1 | 7.2 | 0.666 | 0.676 | 0.029 |
| 21 | Chunan | 36.84 | 127.20 | 29 | 8.9 | 4.5 | 7.0 | 0.670 | 0.688 | 0.044 |
| 22 | Goesan | 36.63 | 127.90 | 27 | 8.4 | 4.3 | 6.8 | 0.594 | 0.651 | 0.106 *** |
| 23 | Andong | 36.52 | 128.88 | 31 | 8.4 | 4.8 | 6.6 | 0.628 | 0.641 | 0.037 |
| 24 | Boeun | 36.52 | 127.82 | 30 | 8.7 | 4.7 | 6.9 | 0.612 | 0.646 | 0.069 ** |
| 25 | Anmyeondo | 36.49 | 126.36 | 43 | 9.5 | 4.5 | 6.7 | 0.630 | 0.667 | 0.067 ** |
| 26 | Boryeong | 36.42 | 126.61 | 36 | 9.3 | 4.5 | 6.9 | 0.670 | 0.665 | 0.006 |
| 27 | YoungYang | 36.55 | 129.19 | 21 | 7.6 | 4.3 | 6.7 | 0.663 | 0.667 | 0.030 |
| 28 | Cheongsong | 36.41 | 129.18 | 45 | 9.7 | 4.5 | 7.0 | 0.697 | 0.690 | 0.001 |
| 29 | Mt. Juwang | 36.39 | 129.15 | 29 | 8.7 | 4.4 | 6.9 | 0.624 | 0.660 | 0.072 ** |
| 30 | Gumi | 36.28 | 128.28 | 30 | 8.1 | 4.3 | 6.5 | 0.681 | 0.684 | 0.022 |
| 31 | Daejeon | 36.28 | 127.45 | 15 | 7.7 | 4.7 | 7.6 | 0.642 | 0.693 | 0.107 ** |
| 32 | Seocheon | 36.12 | 126.67 | 30 | 9.8 | 4.7 | 7.4 | 0.662 | 0.687 | 0.053 * |
| 33 | Mt. Geumo | 36.10 | 128.32 | 29 | 10.0 | 5.3 | 7.9 | 0.647 | 0.728 | 0.128 *** |
| 34 | Geumsan | 36.11 | 127.37 | 21 | 8.1 | 4.3 | 7.1 | 0.631 | 0.675 | 0.090 ** |
| 35 | Angang | 35.97 | 129.20 | 27 | 10.0 | 5.2 | 8.1 | 0.599 | 0.725 | 0.193 *** |
| 36 | Mt. Weebong | 35.91 | 127.26 | 21 | 7.8 | 4.4 | 6.9 | 0.658 | 0.657 | 0.023 |
| 37 | Mt. Deogyu | 35.84 | 127.71 | 30 | 9.5 | 4.7 | 7.3 | 0.667 | 0.688 | 0.047 |
| 38 | Mt. Gaya | 35.82 | 128.12 | 27 | 8.6 | 5.0 | 7.1 | 0.690 | 0.683 | 0.009 |
| 39 | Gyeongju | 35.81 | 129.23 | 30 | 8.8 | 4.5 | 6.8 | 0.608 | 0.633 | 0.057 * |
| 40 | Mt. Daeguap | 35.81 | 128.58 | 37 | 10.1 | 4.8 | 7.2 | 0.685 | 0.660 | −0.024 |
| 41 | Daegu | 35.79 | 128.65 | 20 | 7.5 | 4.8 | 6.7 | 0.645 | 0.645 | 0.025 |
| 42 | Geochang | 35.76 | 127.83 | 28 | 9.2 | 4.8 | 7.2 | 0.646 | 0.666 | 0.048 |
| 43 | Jinan | 35.76 | 127.42 | 30 | 8.9 | 4.9 | 7.0 | 0.685 | 0.681 | 0.011 |
| 44 | Mt. Biseul | 35.72 | 128.51 | 30 | 9.3 | 4.8 | 7.3 | 0.611 | 0.688 | 0.129 *** |
| 45 | Buan | 35.67 | 126.63 | 42 | 9.9 | 5.0 | 7.1 | 0.688 | 0.695 | 0.023 |
| 46 | Ulju | 35.55 | 129.02 | 25 | 8.5 | 4.9 | 7.0 | 0.638 | 0.666 | 0.054 * |
| 47 | Mt. Naejang | 35.50 | 126.90 | 31 | 9.4 | 4.5 | 7.0 | 0.625 | 0.658 | 0.066 ** |
| 48 | Mt. Jiri | 35.37 | 127.57 | 30 | 9.4 | 4.5 | 7.2 | 0.694 | 0.684 | 0.002 |
| 49 | Uiryeong | 35.31 | 128.27 | 30 | 8.8 | 4.7 | 7.0 | 0.709 | 0.695 | −0.003 |
| 50 | Yeonggwang | 35.20 | 126.54 | 30 | 9.1 | 4.7 | 7.0 | 0.629 | 0.668 | 0.075 ** |
| 51 | Kimhae | 35.19 | 128.75 | 29 | 10.0 | 5.5 | 8.0 | 0.697 | 0.733 | 0.066 ** |
| 52 | Hadong | 35.09 | 127.77 | 30 | 9.9 | 5.0 | 7.5 | 0.645 | 0.700 | 0.095 ** |
| 53 | Gadeokdo | 35.02 | 128.83 | 28 | 9.5 | 4.6 | 7.3 | 0.651 | 0.684 | 0.067 * |
| 54 | Goseong | 34.98 | 128.21 | 28 | 10.0 | 5.1 | 8.0 | 0.660 | 0.715 | 0.096 *** |
| 55 | Suncheon | 34.97 | 127.22 | 18 | 7.3 | 4.4 | 6.6 | 0.683 | 0.648 | −0.024 |
| 56 | Hwasun | 34.90 | 126.92 | 30 | 8.1 | 4.4 | 6.6 | 0.642 | 0.676 | 0.067 * |
| 57 | Yeongam | 34.75 | 126.67 | 29 | 8.5 | 4.3 | 6.9 | 0.655 | 0.647 | 0.005 |
| 58 | Goheung | 34.52 | 127.13 | 30 | 9.5 | 4.8 | 7.3 | 0.703 | 0.688 | −0.005 |
| 59 | Wando | 34.34 | 126.64 | 30 | 9.6 | 4.4 | 7.2 | 0.618 | 0.661 | 0.081 ** |
| 60 | Mt. Halla | 33.41 | 126.54 | 36 | 6.5 | 3.3 | 5.0 | 0.606 | 0.609 | 0.019 |
| Mean | 8.9 | 4.6 | 7.0 | 0.652 | 0.673 | 0.048 |
| Locus | Primer Sequence (5′→3′) | Repeat Motif | Range (bp) | Ta (°C) | Na | PIC | Reference |
|---|---|---|---|---|---|---|---|
| Pdms009 | F: FAM-CAATGAGTAGAAGATCATGGTGG | (CT)31(CA)20 | 130–198 | 52 | 34 | 0.888 | Watanabe et al. 2006 |
| R: CTAGGGAGCCGCATTTACAC | |||||||
| Pdms030 | F: FAM-GATCACTGTAGGAAGGCTGG | (CA)12CT(CA)5 | 98–122 | 52 | 13 | 0.502 | Watanabe et al. 2006 |
| R: TGGGAAGAGGACAACCTGAG | |||||||
| Pdms065 | F: FAM-GTCAGAAGCCTTATACTGTG | (TG)3TATAN10(TG)9(AG)10 | 149–193 | 52 | 21 | 0.352 | Watanabe et al. 2006 |
| R: TTGTAAATTCAAATGTAGCC | |||||||
| Pdms221 | F: FAM-GAGAGTTGTATGACGGAAATAC | (GA)9G3(GA)5 | 169–183 | 52 | 8 | 0.549 | Watanabe et al. 2006 |
| R: CCCACACAAAAGTGTACTTC | |||||||
| Pde14 | F: FAM-TCATAGGTACAAAGTCATTACACC | (TC)18(AC)14 | 179–261 | 52 | 30 | 0.881 | Lian et al. 2000 |
| R: CTTCCCCACTTGACTTGAAGT | |||||||
| Lop5 | F: FAM-AGCCGTAAAAGCTATCTTGTG | (TA)33 | 162–190 | 45 | 13 | 0.314 | Liewlaksaneeyanawin et al. 2004 |
| R: GGCATACTTACATTTTAATAA | |||||||
| CPDE0039 | F: FAM-TTCCAAGAACTCCTGGCTCT | (AT)15 | 163–257 | 56 | 40 | 0.912 | Chung et al. 2019 |
| R: GGGAACAGGTCCTCATTTCT | |||||||
| CPDE0058 | F: FAM-CAGTGGTCCACCACACTAACT | (TA)10 | 167–193 | 56 | 12 | 0.742 | Chung et al. 2019 |
| R: GTGTGGACCATGTAAGGTATGC | |||||||
| CPDE0060 | F: FAM-ATTGATGCATGGCACCTG | (GT)16 | 136–162 | 56 | 14 | 0.710 | Chung et al. 2019 |
| R: ACAGGAGTTCCGATGAGGTT | |||||||
| CPDE0076 | F: CTCAACTGGCCACTGTAGAACT | (GA)9 | 182–212 | 56 | 12 | 0.656 | Chung et al. 2019 |
| R: AAGGTTCAGGTTGGCATC | |||||||
| CPDE0106 | F: FAM-CAGATGTTAATCTGGTAGCCCC | (AT)11 | 156–186 | 56 | 16 | 0.799 | Chung et al. 2019 |
| R: CACCTAAGTTGCCACAATGC |
| Analysis | Source of Variation | Df | Sum of Squares | Variance Component | Percentage of Variation (%) |
|---|---|---|---|---|---|
| Hierarchical AMOVA (including Mt. Halla) | Between cluster | 3 | 151.672 | 0.117 | 1.4 ** |
| Among populations | 56 | 540.749 | 0.051 | 0.6 | |
| Within populations | 1784 | 14,419.034 | 8.082 | 98.0 | |
| Total | 1843 | 15,111.456 | 8.251 | 100 | |
| Hierarchical AMOVA (except Mt. Halla) | Between cluster | 2 | 79.801 | 0.061 | 0.74 ** |
| Among populations | 56 | 540.749 | 0.051 | 0.62 | |
| Within populations | 1749 | 14,176.839 | 8.106 | 98.64 | |
| Total | 1843 | 14,797.389 | 8.217 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, J.-Y.; Lee, J.-W.; Hong, K.-N. Genetic Diversity and Structure of Pinus densiflora Siebold & Zucc. Populations in Republic of Korea Based on Microsatellite Markers. Forests 2021, 12, 750. https://doi.org/10.3390/f12060750
Ahn J-Y, Lee J-W, Hong K-N. Genetic Diversity and Structure of Pinus densiflora Siebold & Zucc. Populations in Republic of Korea Based on Microsatellite Markers. Forests. 2021; 12(6):750. https://doi.org/10.3390/f12060750
Chicago/Turabian StyleAhn, Ji-Young, Jei-Wan Lee, and Kyung-Nak Hong. 2021. "Genetic Diversity and Structure of Pinus densiflora Siebold & Zucc. Populations in Republic of Korea Based on Microsatellite Markers" Forests 12, no. 6: 750. https://doi.org/10.3390/f12060750
APA StyleAhn, J.-Y., Lee, J.-W., & Hong, K.-N. (2021). Genetic Diversity and Structure of Pinus densiflora Siebold & Zucc. Populations in Republic of Korea Based on Microsatellite Markers. Forests, 12(6), 750. https://doi.org/10.3390/f12060750