
Generic Additive Allometric Models and Biomass Allocation for Two Natural Oak Species in Northeastern China



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area Description
2.2. Biomass Data
2.3. Data Analysis
2.3.1. Allometric Models Specification
2.3.2. Model Evaluation and Selection
2.3.3. Comparison to Existing Biomass Models
3. Results
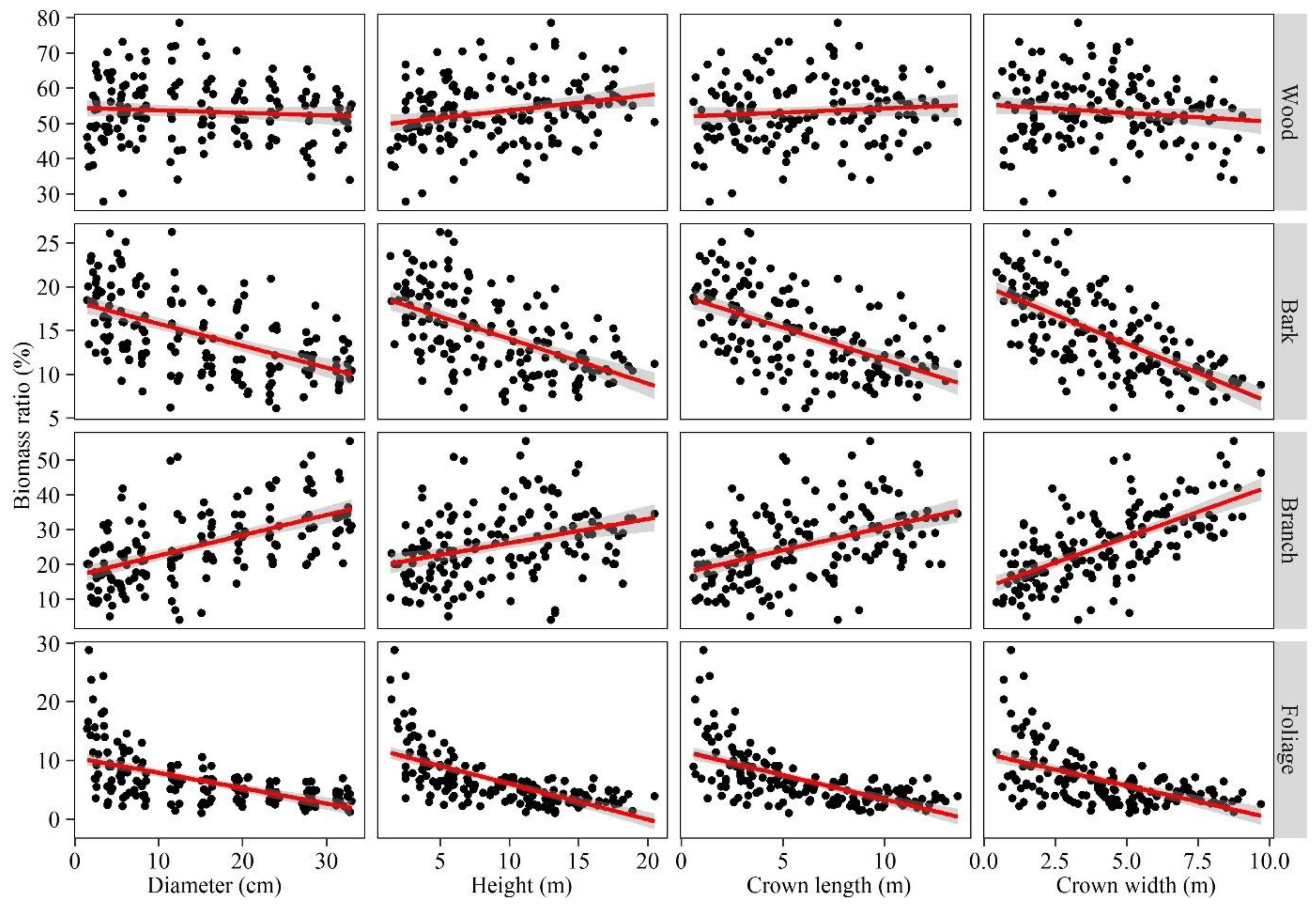
3.1. Aboveground Biomass Partitioning
3.2. Root-to-Shoot Ratio
3.3. Biomass Models
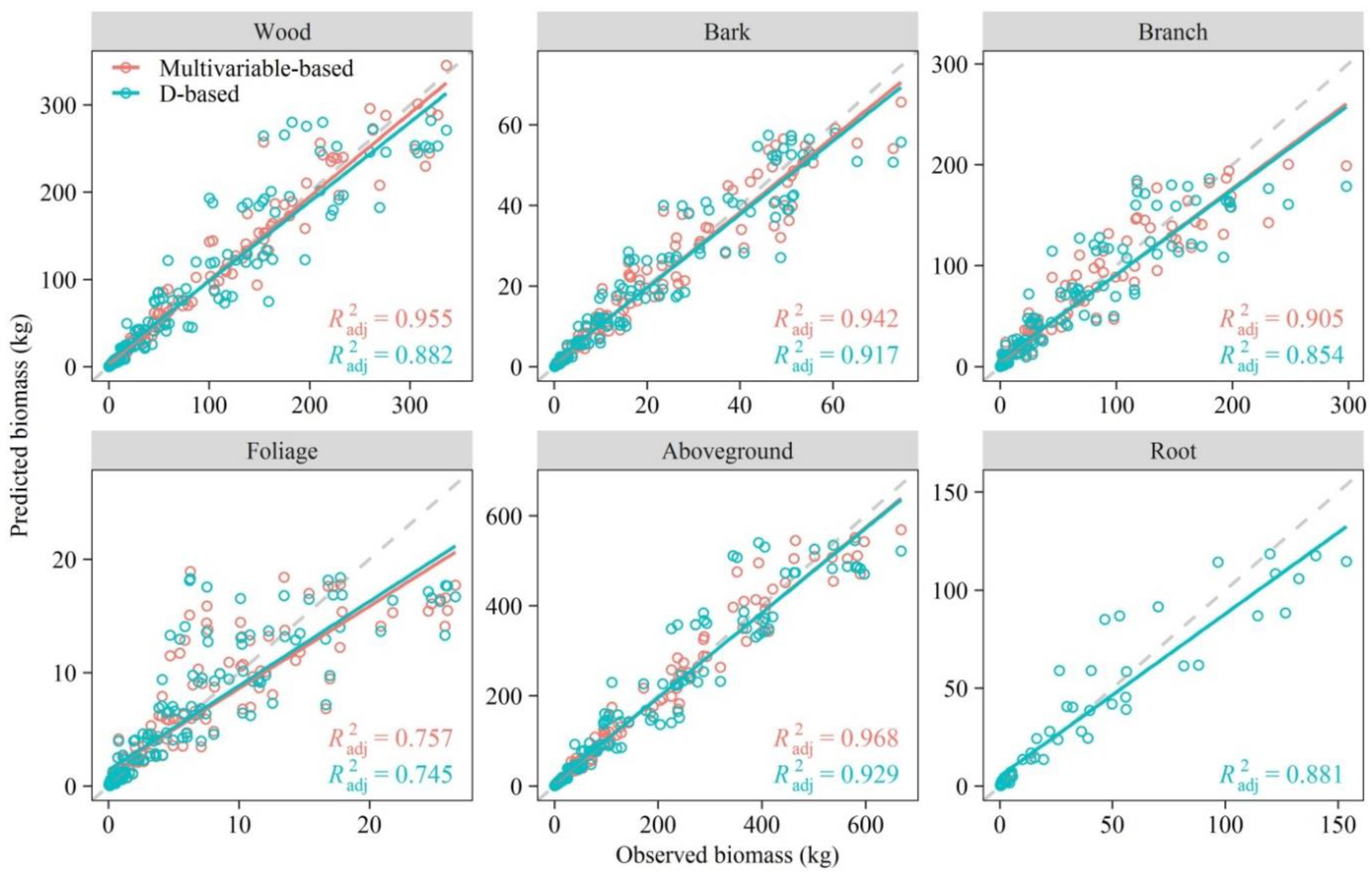
3.4. Model Cross-Validation
4. Discussion
4.1. Biomass Partitioning
4.2. Root–Aboveground Biomass Link
4.3. Allometric Equations
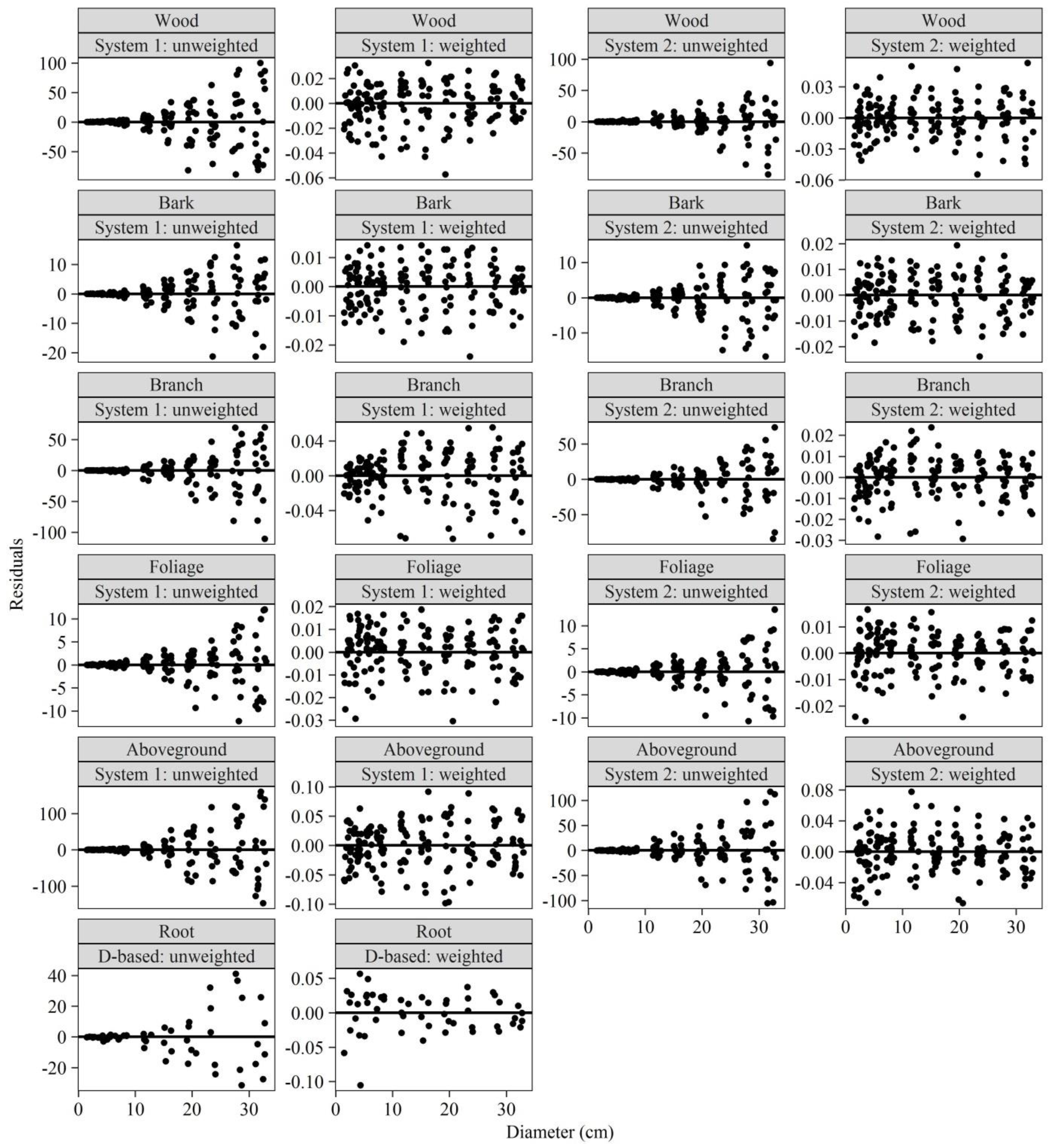
4.4. Heteroscedasticity Elimination
4.5. Performance of Existing Equations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Yin, X.; Zhou, G.; Sui, X.; He, Q.; Li, R. Dominant climatic factors of Quercus mongolica geographical distribution and their thresholds. Acta Ecol. Sin. 2013, 33, 103–109. (In Chinese) [Google Scholar]
- Mugasha, W.A.; Eid, T.; Bollandsas, O.M.; Malimbwi, R.E.; Chamshama, S.A.O.; Zahabu, E.; Katani, J.Z. Allometric models for prediction of above- and belowground biomass of trees in the miombo woodlands of Tanzania. For. Ecol. Manag. 2013, 310, 87–101. [Google Scholar] [CrossRef]
- He, J.S. Carbon cycling of Chinese forests: From carbon storage, dynamics to models. Sci. China Life Sci. 2012, 55, 188–190. [Google Scholar] [CrossRef]
- Alongi, D.M.; Clough, B.F.; Dixon, P.; Tirendi, F. Nutrient partitioning and storage in arid-zone forests of the mangroves Rhizophora stylosa and Avicennia marina. Trees 2003, 17, 51–60. [Google Scholar] [CrossRef]
- Dimobe, K.; Mensah, S.; Goetze, D.; Ouédraogo, A.; Kuyah, S.; Porembski, S.; Thiombiano, A. Aboveground biomass partitioning and additive models for Combretum glutinosum and Terminalia laxiflora in West Africa. Biomass Bioenergy 2018, 115, 151–159. [Google Scholar] [CrossRef]
- Sanaei, A.; Ali, A.; Ahmadaali, K.; Jahantab, E. Generalized and species-specific prediction models for aboveground biomass in semi-steppe rangelands. J. Plant Ecol. 2019, 12, 428–437. [Google Scholar] [CrossRef]
- Chave, J.; Rejou-Mechain, M.; Burquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Dong, L.H.; Zhang, L.J.; Li, F.R. Developing additive systems of biomass equations for nine hardwood species in Northeast China. Trees 2015, 29, 1149–1163. [Google Scholar] [CrossRef]
- He, H.J.; Zhang, C.Y.; Zhao, X.H.; Fousseni, F.; Wang, J.S.; Dai, H.J.; Yang, S.; Zuo, Q. Allometric biomass equations for 12 tree species in coniferous and broadleaved mixed forests, Northeastern China. PLoS ONE 2018, 13, e0186226. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.K. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. For. Ecol. Manag. 2006, 222, 9–16. [Google Scholar] [CrossRef]
- Eker, M.; Poudel, K.P.; Ozcelik, R. Aboveground biomass equations for small trees of brutian pine in turkey to facilitate harvesting and management. Forests 2017, 8, 477. [Google Scholar] [CrossRef]
- Parresol, B.R. Assessing tree and stand biomass: A review with examples and critical comparisons. For. Sci. 1999, 45, 573–593. [Google Scholar]
- Blujdea, V.N.B.; Pilli, R.; Dutca, I.; Ciuvat, L.; Abrudan, I.V. Allometric biomass equations for young broadleaved trees in plantations in Romania. For. Ecol. Manag. 2012, 264, 172–184. [Google Scholar] [CrossRef]
- Kozak, A. Methods for ensuring additivity of biomass components by regression analysis. For. Chron. 1970, 46, 402–405. [Google Scholar] [CrossRef]
- Parresol, B.R. Additivity of nonlinear biomass equations. Can. J. For. Res. 2001, 31, 865–878. [Google Scholar] [CrossRef]
- Tang, S.Z.; Zhang, H.R.; Xu, H. Study on establish and estimate method of compatible biomass model. Sci. Silvae Sin. 1999, 36, 19–27. (In Chinese) [Google Scholar]
- Chiyenda, S.S.; Kozak, A. Additivity of component biomass regression equations when the underlying model is linear. Can. J. For. Res. 1984, 14, 441–446. [Google Scholar] [CrossRef]
- Cao, L.; Li, H.K. Analysis of error structure for additive biomass equations on the use of multivariate likelihood function. Forests 2019, 10, 298. [Google Scholar] [CrossRef]
- Canga, E.; Dieguez-Aranda, I.; Afif-Khouri, E.; Camara-Obregon, A. Above-ground biomass equations for Pinus radiata D. Don in Asturias. For. Syst. 2013, 22, 408–415. [Google Scholar] [CrossRef]
- Zhao, D.H.; Westfall, J.; Coulston, J.W.; Lynch, T.B.; Bullock, B.P.; Montes, C.R. Additive biomass equations for slash pine trees: Comparing three modeling approaches. Can. J. For. Res. 2019, 49, 27–40. [Google Scholar] [CrossRef]
- Helmisaari, H.-S.; Derome, J.; Nöjd, P.; Kukkola, M. Fine root biomass in relation to site and stand characteristics in Norway spruce and Scots pine stands. Tree Physiol. 2007, 27, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Djomo, A.N.; Chimi, C.D. Tree allometric equations for estimation of above, below and total biomass in a tropical moist forest: Case study with application to remote sensing. For. Ecol. Manag. 2017, 391, 184–193. [Google Scholar] [CrossRef]
- Xing, D.L.; Bergeron, J.A.C.; Solarik, K.A.; Tomm, B.; Macdonald, S.E.; Spence, J.R.; He, F.L. Challenges in estimating forest biomass: Use of allometric equations for three boreal tree species. Can. J. For. Res. 2019, 49, 1613–1622. [Google Scholar] [CrossRef]
- Kachamba, D.J.; Eid, T.; Gobakken, T. Above- and belowground biomass models for trees in the miombo woodlands of Malawi. Forests 2016, 7, 38. [Google Scholar] [CrossRef]
- Mokany, K.; Raison, R.J.; Prokushkin, A.S. Critical analysis of root:shoot ratios in terrestrial biomes. Glob. Chang. Biol. 2006, 12, 84–96. [Google Scholar] [CrossRef]
- Mensah, S.; Veldtman, R.; Du Toit, B.; Glèlè Kakaï, R.; Seifert, T. Aboveground biomass and carbon in a south African mistbelt forest and the relationships with tree species diversity and forest structures. Forests 2016, 7, 79. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Folster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Xu, Y.Z.; Zhang, J.X.; Franklin, S.B.; Liang, J.Y.; Ding, P.; Luo, Y.Q.; Lu, Z.J.; Bao, D.C.; Jiang, M.X. Improving allometry models to estimate the above- and belowground biomass of subtropical forest, China. Ecosphere 2015, 6, 289. [Google Scholar] [CrossRef]
- Alvarez, E.; Duque, A.; Saldarriaga, J.; Cabrera, K.; de Las Salas, G.; del Valle, I.; Lema, A.; Moreno, F.; Orrego, S.; Rodriguez, L. Tree above-ground biomass allometries for carbon stocks estimation in the natural forests of Colombia. For. Ecol. Manag. 2012, 267, 297–308. [Google Scholar] [CrossRef]
- Zeng, W.S.; Zhang, L.J.; Chen, X.Y.; Cheng, Z.C.; Ma, K.X.; Li, Z.H. Construction of compatible and additive individual-tree biomass models for Pinus tabulaeformis in China. Can. J. For. Res. 2017, 47, 467–475. [Google Scholar] [CrossRef]
- Zeng, W.S.; Duo, H.R.; Lei, X.D.; Chen, X.Y.; Wang, X.J.; Pu, Y.; Zou, W.T. Individual tree biomass equations and growth models sensitive to climate variables for Larix spp. in China. Eur. J. For. Res. 2017, 136, 233–249. [Google Scholar] [CrossRef]
- De-Miguel, S.; Pukkala, T.; Assaf, N.; Shater, Z. Intra-specific differences in allometric equations for aboveground biomass of eastern Mediterranean Pinus brutia. Ann. For. Sci. 2014, 71, 101–112. [Google Scholar] [CrossRef]
- Mensah, S.; Veldtman, R.; Seifert, T. Allometric models for height and aboveground biomass of dominant tree species in South African Mistbelt forests. South. Forests 2016, 79, 19–30. [Google Scholar] [CrossRef]
- Bi, H.Q.; Murphy, S.; Volkova, L.; Weston, C.; Fairman, T.; Li, Y.; Law, R.; Norris, J.; Lei, X.D.; Caccamo, G. Additive biomass equations based on complete weighing of sample trees for open eucalypt forest species in south-eastern Australia. For. Ecol. Manag. 2015, 349, 106–121. [Google Scholar] [CrossRef]
- Mensah, S.; Kakaï, R.G.; Seifert, T. Patterns of biomass allocation between foliage and woody structure: The effects of tree size and specific functional traits. Ann. For. Res. 2016, 59, 49–60. [Google Scholar] [CrossRef]
- Pajtik, J.; Konopka, B.; Lukac, M. Individual biomass factors for beech, oak and pine in Slovakia: A comparative study in young naturally regenerated stands. Trees 2011, 25, 277–288. [Google Scholar] [CrossRef]
- Cairns, M.A.; Brown, S.; Baumgardner, H.G.A. Root biomass allocation in the world’s upland forests. Oecologia 1997, 111, 1–11. [Google Scholar] [CrossRef]
- Zeng, W.S. Developing one-variable individual tree biomass models based on wood density for 34 tree species in China. For. Resour. Manag. 2017, 41–46. (In Chinese) [Google Scholar]
- George, G. Prediction bias and response surface curvature. For. Sci. 1991, 37, 755–765. [Google Scholar]
- Riofrio, J.; Herrero, C.; Grijalva, J.; Bravo, F. Aboveground tree additive biomass models in Ecuadorian highland agroforestry systems. Biomass Bioenergy 2015, 80, 252–259. [Google Scholar] [CrossRef]
- Bi, H.Q.; Turner, J.; Lambert, M.J. Additive biomass equations for native eucalypt forest trees of temperate Australia. Trees 2004, 18, 467–479. [Google Scholar] [CrossRef]
- Meng, S.W.; Jia, Q.Q.; Liu, Q.J.; Zhou, G.; Wang, H.M.; Yu, J. Aboveground biomass allocation and additive allometric models for natural larix gmelinii in the western Daxing’anling mountains, northeastern China. Forests 2019, 10, 150. [Google Scholar] [CrossRef]
- Kuyah, S.; Sileshi, G.W.; Rosenstock, T.S. Allometric models based on bayesian frameworks give better estimates of aboveground biomass in the miombo woodlands. Forests 2016, 7, 13. [Google Scholar] [CrossRef]
- Abich, A.; Mucheye, T.; Tebikew, M.; Gebremariam, Y.; Alemu, A. Species-specific allometric equations for improving aboveground biomass estimates of dry deciduous woodland ecosystems. J. For. Res. 2019, 30, 1619–1632. [Google Scholar] [CrossRef]
- Tetemke, B.A.; Birhane, E.; Rannestad, M.M.; Eid, T. Allometric models for predicting aboveground biomass of trees in the dry afromontane forests of northern Ethiopia. Forests 2019, 10, 1114. [Google Scholar] [CrossRef]
- Rutishauser, E.; Noor’An, F.; Laumonier, Y.; Halperin, J.; Rufi’ie; Hergoualc’H, K.; Verchot, L. Generic allometric models including height best estimate forest biomass and carbon stocks in Indonesia. For. Ecol. Manag. 2013, 307, 219–225. [Google Scholar] [CrossRef]
- Lambert, M.C.; Ung, C.H.; Raulier, F. Canadian national tree aboveground biomass equations. Can. J. For. Res. 2005, 35, 1996–2018. [Google Scholar] [CrossRef]
- Meng, S.W.; Liu, Q.J.; Zhou, G.; Jia, Q.Q.; Zhuang, H.X.; Zhou, H. Aboveground tree additive biomass equations for two dominant deciduous tree species in Daxing’anling, northernmost China. J. For. Res. 2017, 22, 233–240. [Google Scholar] [CrossRef]
- Wagner, R.G.; Ter-Mikaelian, M.T. Comparison of biomass component equations for four species of northern coniferous tree seedlings. Ann. For. Sci. 1999, 56, 193–199. [Google Scholar] [CrossRef]
- Goodman, R.C.; Phillips, O.L.; Baker, T.R. The importance of crown dimensions to improve tropical tree biomass estimates. Ecol. Appl. 2016, 24, 680–698. [Google Scholar] [CrossRef]
- Xiao, X.; White, E.P.; Hooten, M.B.; Durham, S.L. On the use of log-transformation vs. nonlinear regression for analyzing biological power laws. Ecology 2011, 92, 1887–1894. [Google Scholar] [CrossRef] [PubMed]
- Packard, G.C. On the use of logarithmic transformations in allometric analyses. J. Theor. Biol. 2009, 257, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Packard, G.C. On the use of log-transformation versus nonlinear regression for analyzing biological power laws. Biol. J. Linn. Soc. 2014, 113, 1167–1178. [Google Scholar] [CrossRef]
- Sprugel, D.G. Correcting for bias in log-transformed allometric equations. Ecology 1983, 64, 209–210. [Google Scholar] [CrossRef]
- Zhao, D.H.; Kane, M.; Markewitz, D.; Teskey, R.; Clutter, M. Additive tree biomass equations for Midrotation loblolly pine plantations. For. Sci. 2015, 61, 613–623. [Google Scholar] [CrossRef]
- Wang, X.W.; Zhao, D.H.; Liu, G.F.; Yang, C.J.; Teskey, R.O. Additive tree biomass equations for Betula platyphylla Suk. plantations in Northeast China. Ann. For. Sci. 2018, 75, 60. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Statistics | n | Mean | Min | Max | SD |
|---|---|---|---|---|---|
| D | 159 | 15.1 | 1.5 | 33 | 10.0 |
| H | 159 | 9.11 | 1.4 | 20.5 | 4.94 |
| CW | 159 | 4.22 | 0.45 | 9.70 | 3.51 |
| CL | 159 | 6.13 | 0.60 | 13.61 | 3.51 |
| Wood | 159 | 73.55 | 0.14 | 336.27 | 90.67 |
| Bark | 159 | 16.26 | 0.06 | 74.66 | 19.13 |
| Branch | 159 | 45.19 | 0.03 | 298.32 | 62.55 |
| Foliage | 159 | 5.59 | 0.03 | 26.57 | 6.75 |
| Aboveground | 159 | 140.59 | 0.33 | 668.12 | 173.45 |
| Root | 53 | 37.66 | 0.23 | 153.71 | 4.08 |
| Study | Model Type | Predictor | D Range (cm) | Sample Size | Site | n |
|---|---|---|---|---|---|---|
| Dong et al. [8] | Type 1 | D | 4.2–37.1 | 64 | HLJ, JL | 100 |
| Type 2 | D, H | |||||
| Wang [10] | Type 3 | D | 4.3–57.1 | 10 | HLJ | 99 |
| He et al. [9] | Type 4 | D | 4.2–41.2 | 10 | JL | 100 |
| Component | Model Predictor | Regression Coefficients | Fitting Statistics | Weight Function | |||||
|---|---|---|---|---|---|---|---|---|---|
| β0 | β1 | β2 | β3 | β4 | RMSE (kg) | ||||
| Wood | D | 0.050 *** | 2.473 *** | 30.602 | 0.886 | D4.9241 | |||
| D, H | 0.033 *** | 1.761 *** | 1.043 *** | 19.138 | 0.955 | D4.2932 | |||
| D, CW | 0.061 *** | 2.252 *** | 0.271 *** | 30.605 | 0.886 | D3.3857 | |||
| D, CL | 0.059 *** | 1.845 *** | 0.826 *** | 25.121 | 0.923 | D3.6802 | |||
| D, H, CW | 0.034 *** | 1.689 *** | 1.031 *** | 0.121 * | 18.833 | 0.957 | D4.3493 | ||
| D, H, CL | 0.030 *** | 1.834 *** | 1.191 *** | −0.235 *** | 18.968 | 0.956 | D5.1990 | ||
| D, H, CW, CL | 0.034 *** | 1.723 *** | 1.173 *** | 0.174 *** | −0.252 *** | 18.548 | 0.958 | D4.3589 | |
| Bark | D | 0.022 *** | 2.249 *** | 5.397 | 0.920 | D4.2996 | |||
| D, H | 0.018 *** | 1.916 *** | 0.491 *** | 4.571 | 0.943 | D4.1216 | |||
| D, CW | 0.022 *** | 2.291 *** | −0.076 ns | 5.354 | 0.922 | D4.2709 | |||
| D, CL | 0.024 *** | 1.975 *** | 0.367 *** | 4.835 | 0.936 | D4.0130 | |||
| D, H, CW | 0.017 *** | 2.022 *** | 0.467 *** | −0.146 * | 4.466 | 0.946 | D4.1201 | ||
| D, H, CL | 0.018 *** | 1.939 *** | 0.565 *** | −0.107 ns | 4.623 | 0.942 | D4.1430 | ||
| D, H, CW, CL | 0.017 *** | 2.003 *** | 0.560 *** | −0.090 ns | −0.109 ns | 4.561 | 0.943 | D4.1967 | |
| Branch | D | 0.011 *** | 2.772 *** | 23.320 | 0.861 | D4.2891 | |||
| D, H | 0.012 *** | 2.716 *** | 0.035 ns | 23.440 | 0.860 | D4.3426 | |||
| D, CW | 0.018 *** | 2.090 *** | 0.939 *** | 19.081 | 0.907 | D4.9263 | |||
| D, CL | 0.014 *** | 2.504 *** | 0.290 ** | 23.052 | 0.864 | D5.0652 | |||
| D, H, CW | 0.020 *** | 2.128 *** | −0.110 ns | 0.964 *** | 19.060 | 0.907 | D4.8475 | ||
| D, H, CL | 0.019 *** | 2.572 *** | −0.538 ** | 0.692 *** | 22.606 | 0.869 | D3.4718 | ||
| D, H, CW, CL | 0.033 *** | 1.976 *** | −0.655 *** | 0.985 *** | 0.607 *** | 19.426 | 0.904 | D4.6120 | |
| Foliage | D | 0.019 *** | 1.964 *** | 3.350 | 0.754 | D3.7889 | |||
| D, H | 0.025 *** | 1.981 *** | −0.126 ns | 3.429 | 0.742 | D2.9305 | |||
| D, CW | 0.023 *** | 1.626 *** | 0.473 *** | 3.275 | 0.765 | D3.9640 | |||
| D, CL | 0.017 *** | 2.029 *** | −0.050 ns | 3.370 | 0.751 | D3.4793 | |||
| D, H, CW | 0.020 *** | 1.747 *** | −0.052 ns | 0.441 ** | 3.296 | 0.762 | D3.0790 | ||
| D, H, CL | 0.025 *** | 1.930 *** | −0.053 ns | −0.018 ns | 3.401 | 0.746 | D2.9473 | ||
| D, H, CW, CL | 0.043 *** | 1.692 *** | −0.704 *** | 0.590 *** | 0.323 * | 3.664 | 0.706 | D3.9980 | |
| Root | D | 0.086 *** | 2.072 *** | 14.843 | 0.887 | D4.2914 | |||
| D, H | 0.077 *** | 2.118 *** | −0.008 ns | 14.725 | 0.888 | D3.6076 | |||
| D, CW | 0.087 *** | 2.033 *** | 0.058 ns | 14.991 | 0.884 | D4.2293 | |||
| D, CL | 0.076 *** | 2.091 *** | 0.044 ns | 14.518 | 0.892 | D3.3309 | |||
| D, H, CW | 0.075 ** | 2.112 *** | 0.012 ns | −0.002 ns | 14.769 | 0.888 | D3.4907 | ||
| D, H, CL | 0.081 ** | 2.099 *** | −0.076 ns | 0.083 ns | 14.881 | 0.886 | D3.5074 | ||
| D, H, CW, CL | 0.081 ** | 2.104 *** | −0.069 ns | −0.008 ns | 0.081 ns | 15.008 | 0.884 | D3.4890 | |
| Model System | Component | Model Expressions | RMSE (kg) | |
|---|---|---|---|---|
| System 1 | Wood | Wwd = 0.050D2.473 | 30.602 | 0.886 |
| Bark | Wbk = 0.022D2.249 | 5.397 | 0.920 | |
| Branch | Wbr = 0.011D2.772 | 23.320 | 0.861 | |
| Foliage | Wfo = 0.019D1.964 | 3.350 | 0.754 | |
| Aboveground | Wag = 0.050D2.473 + 0.022D2.249 + 0.011D2.772 + 0.019D1.964 | 45.478 | 0.931 | |
| System 2 | Wood | Wwd = 0.028D1.740H1.299CW0.140CL−0.312 | 18.411 | 0.959 |
| Bark | Wbk = 0.017D2.010H0.502CW−0.132 | 4.473 | 0.945 | |
| Branch | Wbr = 0.017D2.156CW0.870 | 19.005 | 0.908 | |
| Foliage | Wfo = 0.023D1.645CW0.438 | 3.280 | 0.764 | |
| Aboveground | Wag = 0.028D1.740H1.299CW0.140CL−0.312 + 0.017D2.010H0.502CW−0.132 + 0.017D2.156CW0.870 + 0.023D1.645CW0.438 | 31.021 | 0.968 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, S.; Yang, F.; Hu, S.; Wang, H.; Wang, H. Generic Additive Allometric Models and Biomass Allocation for Two Natural Oak Species in Northeastern China. Forests 2021, 12, 715. https://doi.org/10.3390/f12060715
Meng S, Yang F, Hu S, Wang H, Wang H. Generic Additive Allometric Models and Biomass Allocation for Two Natural Oak Species in Northeastern China. Forests. 2021; 12(6):715. https://doi.org/10.3390/f12060715
Chicago/Turabian StyleMeng, Shengwang, Fan Yang, Sheng Hu, Haibin Wang, and Huimin Wang. 2021. "Generic Additive Allometric Models and Biomass Allocation for Two Natural Oak Species in Northeastern China" Forests 12, no. 6: 715. https://doi.org/10.3390/f12060715
APA StyleMeng, S., Yang, F., Hu, S., Wang, H., & Wang, H. (2021). Generic Additive Allometric Models and Biomass Allocation for Two Natural Oak Species in Northeastern China. Forests, 12(6), 715. https://doi.org/10.3390/f12060715