
Growth and Photosynthetic Responses of Seedlings of Japanese White Birch, a Fast-Growing Pioneer Species, to Free-Air Elevated O3 and CO2

 ,
,  , , , and
, , , and
Abstract
1. Introduction
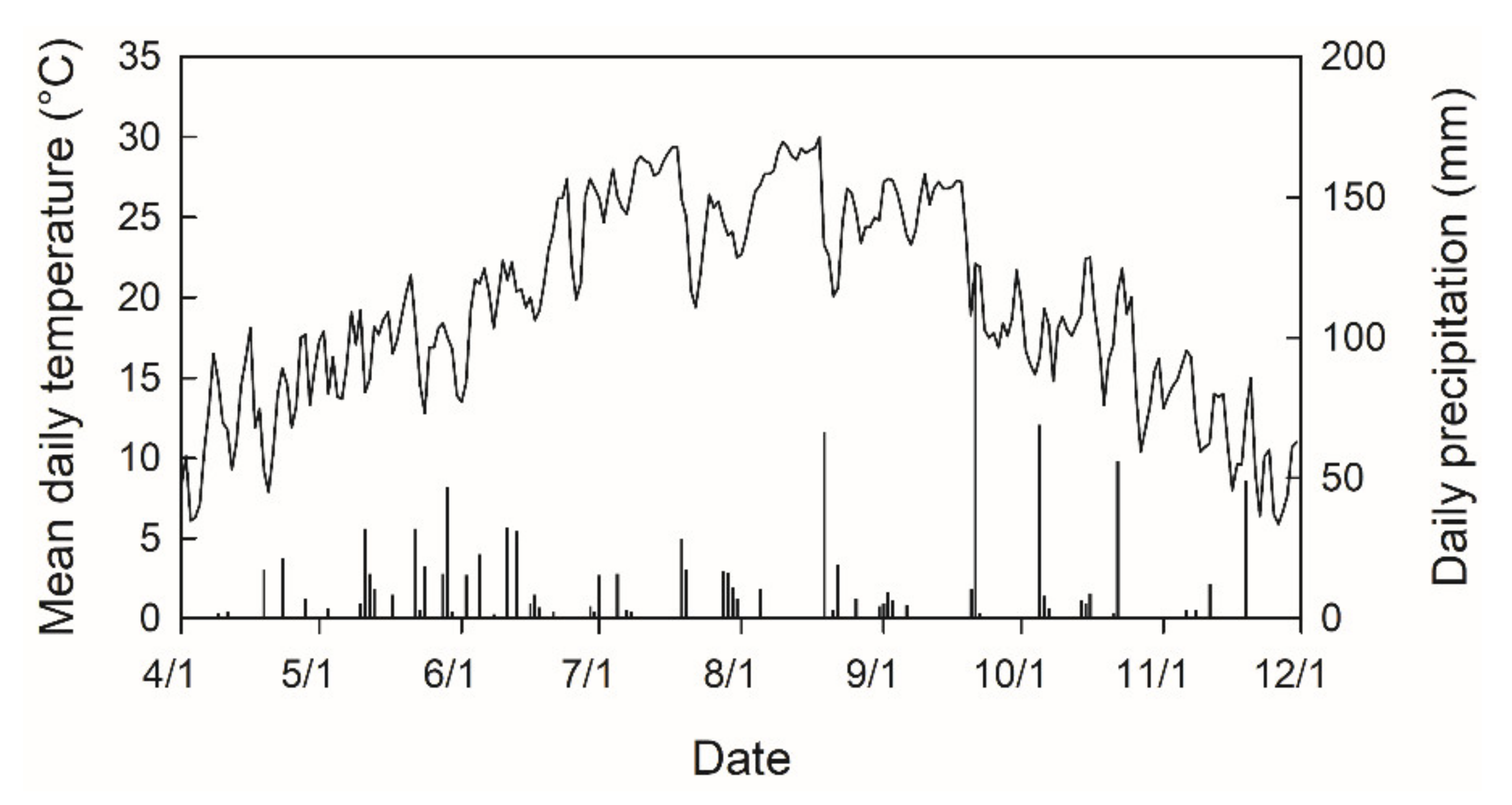
2. Materials and Methods
2.1. Plant Materials
2.2. Measurements of Gas Exchange
2.3. Growth and Biomass Allocation
2.4. Leaf Nitrogen Content
2.5. Statistical Analysis
3. Results
3.1. Growth Responses of Japanese white Birch Seedlings to Elevated CO2 and O3
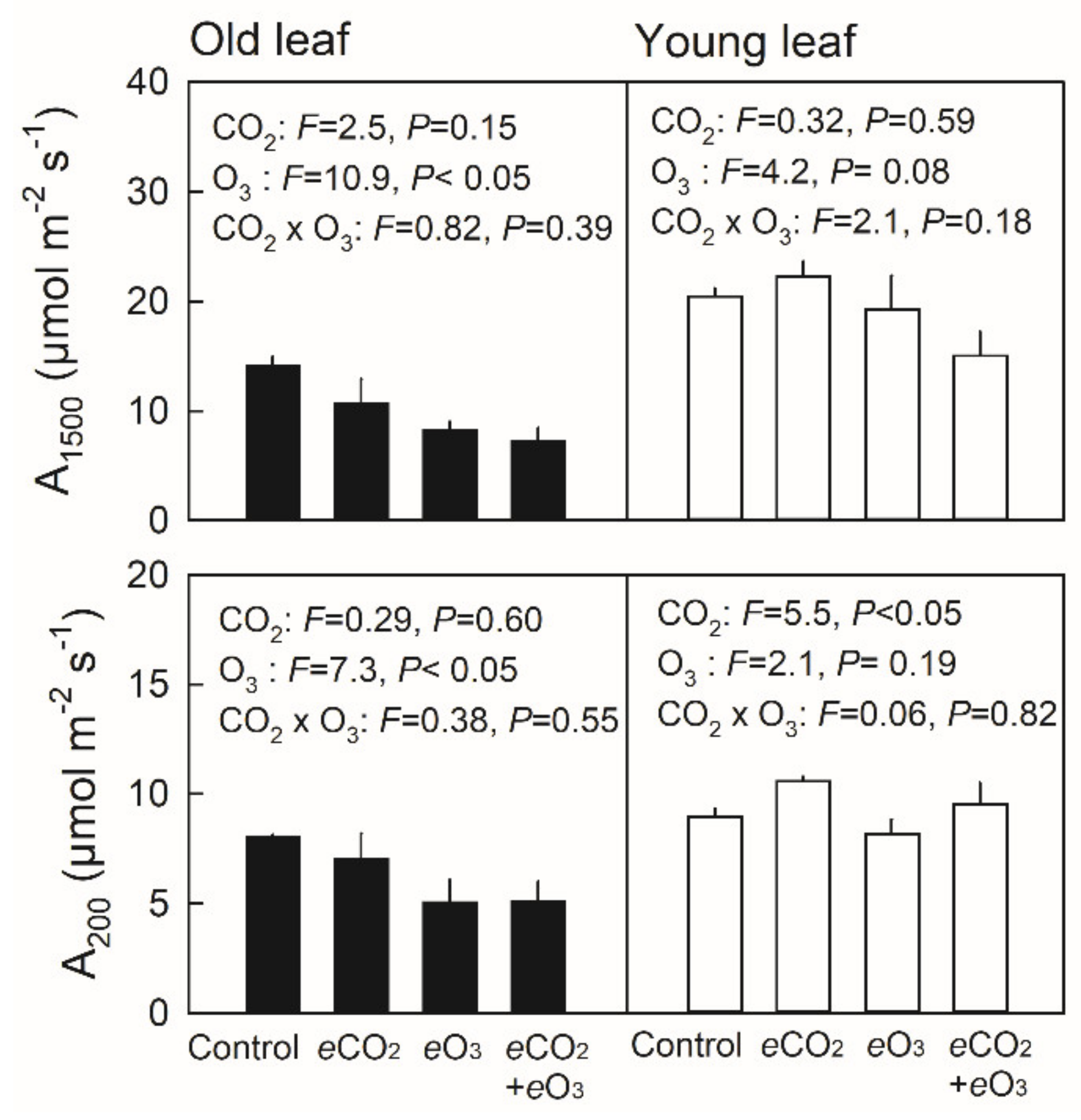
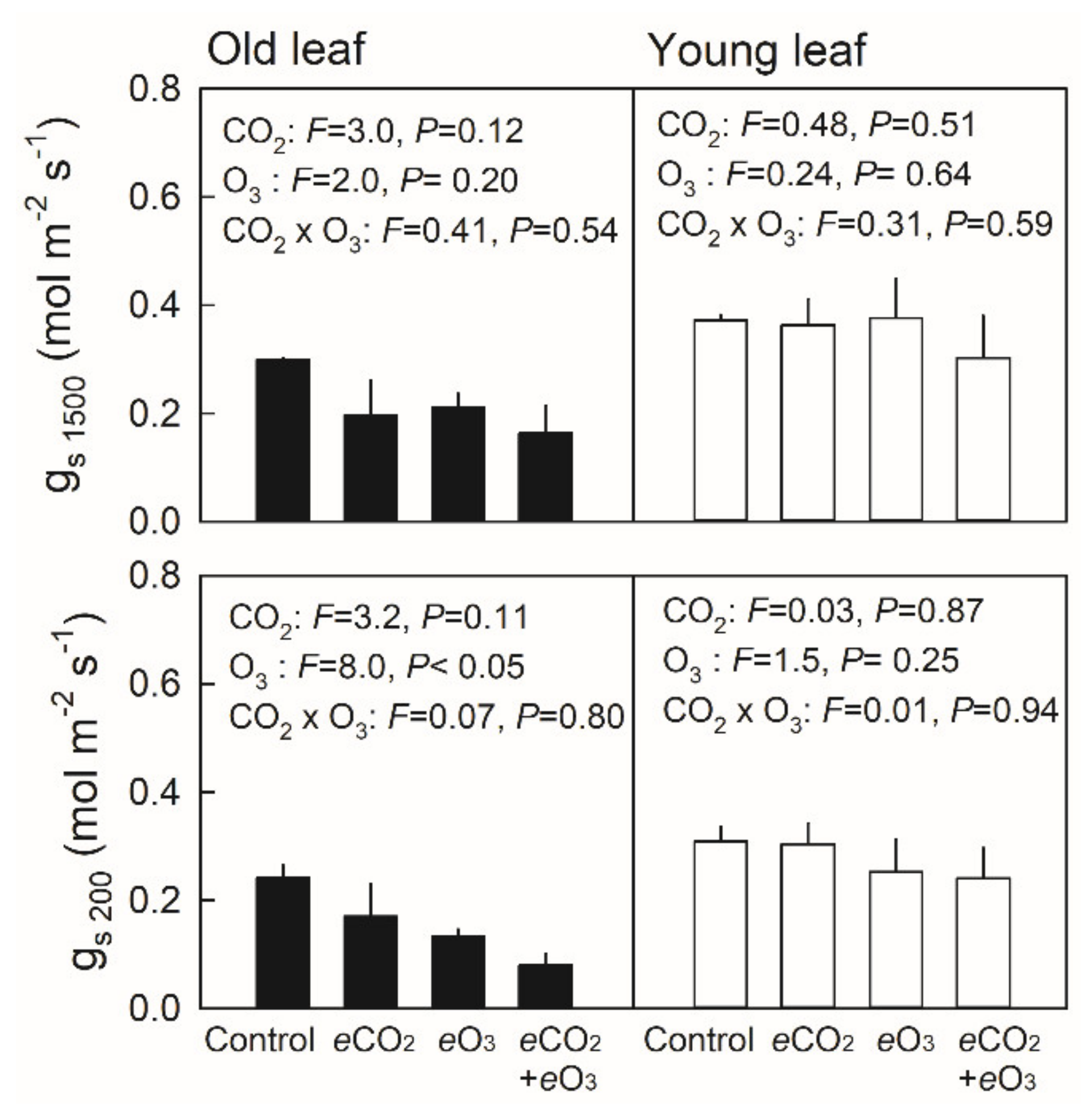
3.2. Net Photosynthetic Rate and Stomatal Conductance in Young and Old Leaves of Japanese Birch Seedlings Grown under the Combination of CO2 and O3
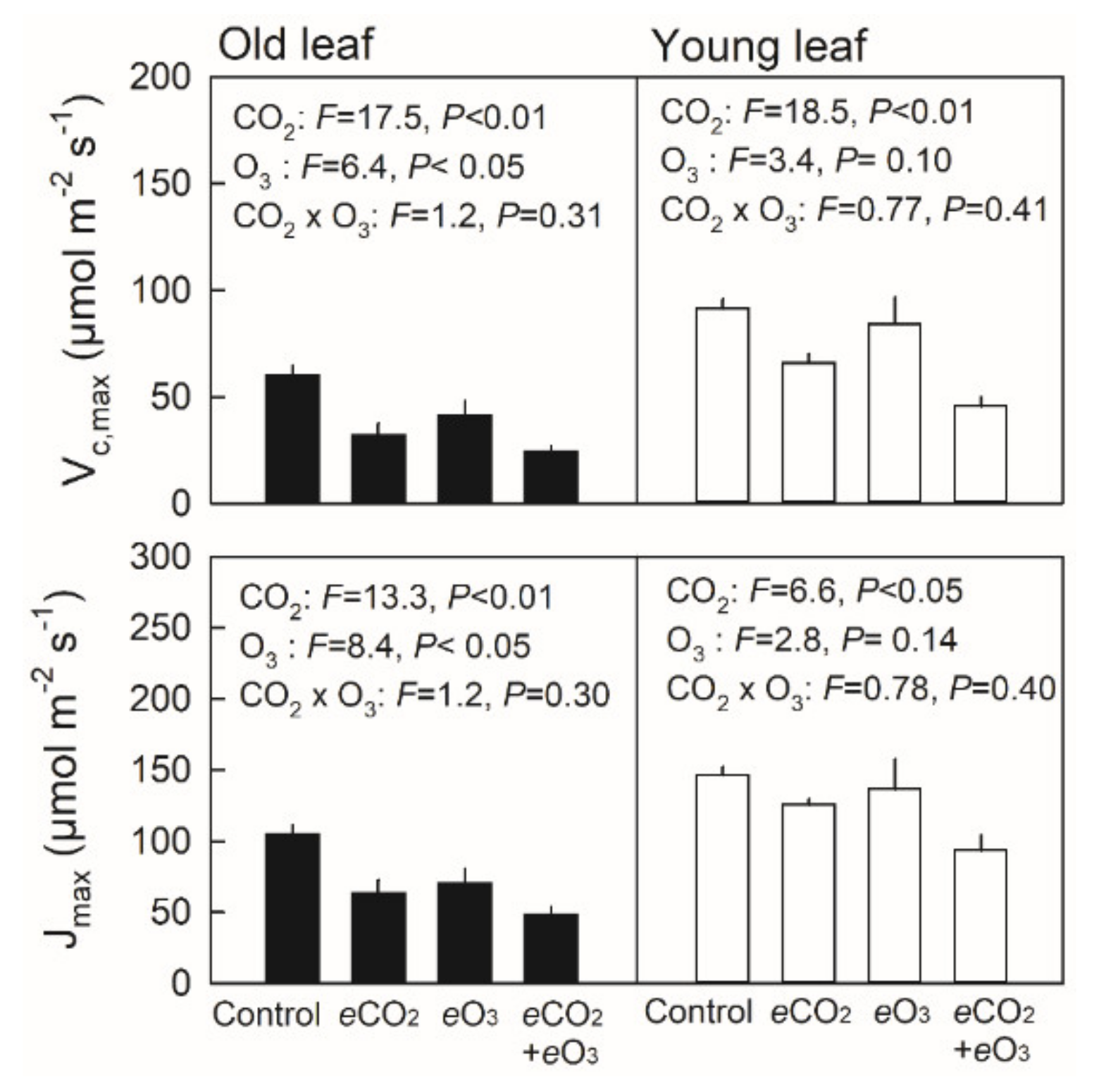
3.3. Maximum Rates of Rubisco Carboxylation (Vc,max) and RuBP Regeneration (Jmax) in Young and Old Leaves of Japanese Birch Seedlings Grown under the Combination of CO2 and O3
3.4. Leaf Nitrogen Content
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007; ISBN 978-0521705967. [Google Scholar]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013; ISBN 978-1-107-05799-1. [Google Scholar]
- Takigawa, M.; Niwano, M.; Akimoto, H.; Takahashi, M.; Kobayashi, K. Projection of surface ozone over East Asia in 2020. J. Agric. Meteorol. 2009, 65, 161–166. [Google Scholar] [CrossRef][Green Version]
- Li, K.; Jacob, D.J.; Liao, H.; Qiu, Y.; Shen, L.; Zhai, S.; Bates, K.H.; Sulprizio, M.P.; Song, S.; Lu, X.; et al. Ozone pollution in the North China Plain spreading into the late-winter haze season. Proc. Natl. Acad. Sci. USA 2021, 118, e2015797118. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Agrawal, M. Air pollutant levels are 12 times higher than guidelines in Varanasi, India. Sources and transfer. Environ. Chem. Lett. 2018, 16, 1009–1016. [Google Scholar] [CrossRef]
- Li, P.; De Marco, A.; Feng, Z.; Anav, A.; Zhou, D.; Paoletti, E. Nationwide ground-level ozone measurements in China suggest serious risks to forests. Environ. Pollut. 2018, 237, 803–813. [Google Scholar] [CrossRef]
- Matyssek, R.; Wieser, G.; Ceulemans, R.; Rennenberg, H.; Pretzsch, H.; Haberer, K.; Löw, M.; Nunn, A.J.J.; Werner, H.; Wipfler, P.; et al. Enhanced ozone strongly reduces carbon sink strength of adult beech (Fagus sylvatica)—Resume from the free-air fumigation study at Kranzberg Forest. Environ. Pollut. 2010, 158, 2527–2532. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Yendrek, C.R.; Sitch, S.; Collins, W.J.; Emberson, L.D. The effects of tropospheric ozone on net primary productivity and implications for climate change. Annu. Rev. Plant Biol. 2012, 63, 637–661. [Google Scholar] [CrossRef]
- Agathokleous, E.; Kitao, M.; Kinose, Y. A review study on ozone phytotoxicity metrics for setting critical levels in Asia. Asian J. Atmos. Environ. 2018, 12, 1–16. [Google Scholar] [CrossRef]
- Feng, Z.; De Marco, A.; Anav, A.; Gualtieri, M.; Sicard, P.; Tian, H.; Fornasier, F.; Tao, F.; Guo, A.; Paoletti, E. Economic losses due to ozone impacts on human health, forest productivity and crop yield across China. Environ. Int. 2019, 131, 104966. [Google Scholar] [CrossRef]
- Mukherjee, A.; Yadav, D.S.; Agrawal, S.B.; Agrawal, M. Ozone a persistent challenge to food security in India: Current status and policy implications. Curr. Opin. Environ. Sci. Health 2021, 19, 100220. [Google Scholar] [CrossRef]
- Hoshika, Y.; Watanabe, M.; Inada, N.; Koike, T. Growth and leaf gas exchange in three birch species exposed to elevated ozone and CO2 in summer. Water. Air. Soil Pollut. 2012, 223, 5017–5025. [Google Scholar] [CrossRef]
- Watanabe, M.; Hoshika, Y.; Inada, N.; Wang, X.; Mao, Q.; Koike, T. Photosynthetic traits of Siebold’s beech and oak saplings grown under free air ozone exposure in northern Japan. Environ. Pollut. 2013, 174, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Feng, Z.; Catalayud, V.; Yuan, X.; Xu, Y.; Paoletti, E. A meta-analysis on growth, physiological and biochemical responses of woody species to ground-level ozone highlights the role of plant functional types. Plant Cell Environ. 2017, 40, 2369–2380. [Google Scholar] [CrossRef]
- Hoshika, Y.; Watanabe, M.; Inada, N.; Mao, Q.; Koike, T. Photosynthetic response of early and late leaves of white birch (Betula platyphylla var. japonica) grown under free-air ozone exposure. Environ. Pollut. 2013, 182, 242–247. [Google Scholar]
- Kitao, M.; Löw, M.; Heerdt, C.; Grams, T.E.E.; Häberle, K.-H.; Matyssek, R. Effects of chronic elevated ozone exposure on gas exchange responses of adult beech trees (Fagus sylvatica) as related to the within-canopy light gradient. Environ. Pollut. 2009, 157, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Hoshika, Y.; Watanabe, M.; Kitao, M.; Häberle, K.-H.; Grams, T.E.E.; Koike, T.; Matyssek, R. Ozone induces stomatal narrowing in European and Siebold’s beeches: A comparison between two experiments of free-air ozone exposure. Environ. Pollut. 2015, 196, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Norby, R.J.; Zak, D.R. Ecological Lessons from Free-Air CO2 Enrichment (FACE) Experiments. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 181–203. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef]
- Koike, T.; Watanabe, M.; Watanabe, Y.; Agathokleous, E.; Eguchi, N.; Takagi, K.; Satoh, F.; Kitaoka, S.; Funada, R. Ecophysiology of deciduous trees native to Northeast Asia grown under FACE (Free Air CO2 Enrichment). J. Agric. Meteorol. 2015, 71, 174–184. [Google Scholar] [CrossRef]
- Rogers, A.; Ellsworth, D.S. Photosynthetic acclimation of Pinus taeda (loblolly pine) to long-term growth in elevated pCO2 (FACE). Plant Cell Environ. 2002, 25, 851–858. [Google Scholar] [CrossRef]
- Poorter, H. Interspecific variation in relative growth rate: On ecological causes and physiological consequences. In Causes and Consequences of Variation in Growth Rate and Productivity of Higher Plants; Lambers, H., Cambridge, M.L., Konings, H., Pons, T.L., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1989; pp. 45–68. ISBN 9051030339. [Google Scholar]
- Landolt, W.; Bühlmann, U.; Bleuler, P.; Bucher, J. Ozone exposure–response relationships for biomass and root/shoot ratio of beech (Fagus sylvatica), ash (Fraxinus excelsior), Norway spruce (Picea abies) and Scots pine (Pinus sylvestris). Environ. Pollut. 2000, 109, 473–478. [Google Scholar] [CrossRef]
- Oksanen, E.; Rousi, M. Differences of Betula origins in ozone sensitivity based on open-field experiment over two growing seasons. Can. J. For. Res. 2001, 31, 804–811. [Google Scholar] [CrossRef]
- Calatayud, V.; Cerveró, J.; Calvo, E.; García-Breijo, F.J.; Reig-Arminana, J.; Sanz, M.J. Responses of evergreen and deciduous Quercus species to enhanced ozone levels. Environ. Pollut. 2011, 159, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Agathokleous, E.; Saitanis, C.J.; Wang, X.; Watanabe, M.; Koike, T. A review study on past 40 years of research on effects of tropospheric O3 on belowground structure, functioning, and processes of trees: A linkage with potential ecological implications. Water Air Soil Pollut. 2016, 227, 33. [Google Scholar] [CrossRef]
- Kikuzawa, K. Leaf survival of woody plants in deciduous broad-leaved forests. 1. Tall trees. Can. J. Bot. 1983, 61, 2133–2139. [Google Scholar] [CrossRef]
- Koike, T. Leaf structure and photosynthetic performance as related to the forest succession of deciduous broad-leaved trees 1. Plant Species Biol. 1988, 3, 77–87. [Google Scholar] [CrossRef]
- Kitao, M.; Komatsu, M.; Yazaki, K.; Kitaoka, S.; Tobita, H. Growth overcompensation against O3 exposure in two Japanese oak species, Quercus mongolica var. crispula and Quercus serrata, grown under elevated CO2. Environ. Pollut. 2015, 206, 133–141. [Google Scholar] [PubMed]
- Tobita, H.; Komatsu, M.; Harayama, H.; Yazaki, K.; Kitaoka, S.; Kitao, M. Effects of combined CO2 and O3 exposures on net CO2 Assimilation and biomass allocation in seedlings of the late-successional Fagus crenata. Climate 2019, 7, 117. [Google Scholar] [CrossRef]
- Koike, T. Physiological ecology of the growth characteristics of Japanese mountain birch in northern Japan: A comparison with Japanese mountain white birch. In Vegetation Science in Forestry: Global Perspective Based on Forest Ecosystems of East and Southeast Asia; Box, E.E.O., Peet, R.K., Masuzawa, T., Yamada, I., Fujiwara, K., Maycock, P.F., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 409–422. ISBN 978-0-7923-2679-3. [Google Scholar]
- Leverenz, J.W.; Falk, S.; Pilström, C.M.; Samuelsson, G. The effects of photoinhibition on the photosynthetic light-response curve of green plant cells (Chlamydomonas reinhardtii). Planta 1990, 182, 161–168. [Google Scholar] [CrossRef]
- Long, S.P.; Bernacchi, C.J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef]
- Bernacchi, C.J.; Singsaas, E.L.; Pimentel, C.; Portis, A.R., Jr.; Long, S.P. Improved temperature response functions for models of Rubisco-limited photosynthesis. Plant Cell Environ. 2001, 24, 253–259. [Google Scholar] [CrossRef]
- Wilson, K.B.; Baldocchi, D.D.; Hanson, P.J. Spatial and seasonal variability of photosynthetic parameters and their relationship to leaf nitrogen in a deciduous forest. Tree Physiol. 2000, 20, 565–578. [Google Scholar] [CrossRef]
- De Kauwe, M.G.; Lin, Y.; Wright, I.J.; Medlyn, B.E.; Crous, K.Y.; Ellsworth, D.S.; Maire, V.; Prentice, I.C.; Atkin, O.K.; Rogers, A.; et al. A test of the ‘one-point method’ for estimating maximum carboxylation capacity from field-measured, light-saturated photosynthesis. New Phytol. 2016, 210, 1130–1144. [Google Scholar] [CrossRef] [PubMed]
- Kitao, M.; Agathokleous, E.; Harayama, H.; Yazaki, K.; Tobita, H. Constant ratio of Cc to Ci under various CO2 concentrations and light intensities, and during progressive drought, in seedlings of Japanese white birch. Photosynth. Res. 2021, 147, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage Publications: Thousand Oaks, CA, USA, 2019; ISBN 9781544336473. [Google Scholar]
- McDonald, E.P.; Kruger, E.L.; Riemenschneider, D.E.; Isebrands, J.G. Competitive status influences tree-growth responses to elevated CO2 and O3 in aggrading aspen stands. Funct. Ecol. 2002, 16, 792–801. [Google Scholar] [CrossRef]
- Watanabe, M.; Umemoto-Yamaguchi, M.; Koike, T.; Izuta, T. Growth and photosynthetic response of Fagus crenata seedlings to ozone and/or elevated carbon dioxide. Landsc. Ecol. Eng. 2010, 6, 181–190. [Google Scholar] [CrossRef]
- Seiwa, K.; Kikuzawa, K.; Kadowaki, T.; Akasaka, S.; Ueno, N. Shoot life span in relation to successional status in deciduous broad-leaved tree species in a temperate forest. New Phytol. 2006, 169, 537–548. [Google Scholar] [CrossRef]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef]
- Kiba, T.; Takei, K.; Kojima, M.; Sakakibara, H. Side-chain modification of cytokinins controls shoot growth in Arabidopsis. Dev. Cell 2013, 27, 452–461. [Google Scholar] [CrossRef]
- Winwood, J.; Pate, A.E.; Price, A.J.; Hanke, D.E. Effects of long-term, free-air ozone fumigation on the cytokinin content of mature beech trees. Plant Biol. 2007, 9, 265–278. [Google Scholar] [CrossRef]
- Kitao, M.; Koike, T.; Tobita, H.; Maruyama, Y. Elevated CO2 and limited nitrogen nutrition can restrict excitation energy dissipation in photosystem II of Japanese white birch (Betula platyphylla var. japonica) leaves. Physiol. Plant. 2005, 125, 64–73. [Google Scholar] [CrossRef]
- Koike, T.; Kitao, M.; Hikosaka, K.; Agathokleous, E.; Watanabe, Y.; Watanabe, M.; Eguchi, N.; Funada, R. Photosynthetic and photosynthesis-related responses of Japanese native trees to CO2: Results from phytotrons, open-top chambers, natural CO2 springs, and free-air CO2 enrichment. In The Leaf: A Platform for Performing Photosynthesis. Advances in Photosynthesis and Respiration (Including Bioenergy and Related Processes); Adams, W.W., III, Terashima, I., Eds.; Springer: Cham, Switzerland, 2018; pp. 425–449. ISBN 978-3-319-93592-8. [Google Scholar]
- Sperdouli, I.; Moustakas, M. Leaf developmental stage modulates metabolite accumulation and photosynthesis contributing to acclimation of Arabidopsis thaliana to water deficit. J. Plant Res. 2014, 127, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Taub, D.R.; Wang, X. Why are nitrogen concentrations in plant tissues lower under elevated CO2? A critical examination of the hypotheses. J. Integr. Plant Biol. 2008, 50, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- Von Caemmerer, S. Biochemical Models of Leaf Photosynthesis; CSIRO Pub: Collingwood, Australia, 2000; ISBN 064306379X. [Google Scholar]
- Kitao, M.; Hida, T.; Eguchi, N.; Tobita, H.; Utsugi, H.; Uemura, A.; Kitaoka, S.; Koike, T. Light compensation points in shade-grown seedlings of deciduous broadleaf tree species with different successional traits raised under elevated CO2. Plant Biol. 2016, 18, 22–27. [Google Scholar] [CrossRef]
- Matyssek, R.; Baumgarten, M.; Hummel, U.; Häberle, K.-H.; Kitao, M.; Wieser, G. Canopy-level stomatal narrowing in adult Fagus sylvatica under O3 stress—Means of preventing enhanced O3 uptake under high O3 exposure? Environ. Pollut. 2015, 196, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Kinose, Y.; Matsumura, H.; Izuta, T. Evaluation of O3 Effects on Cumulative Photosynthetic CO2 Uptake in Seedlings of Four Japanese Deciduous Broad-Leaved Forest Tree Species Based on Stomatal O3 Uptake. Forests 2019, 10, 556. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Control | eCO2 | eO3 | eCO2 + eO3 |
|---|---|---|---|---|
| CO2 (µmol mol−1) | 377 ± 1.0 | 563 ± 9.0 | 381 ± 1.2 | 546 ± 2.3 |
| O3 (nmol mol−1) | 30.2 ± 0.4 | 30.2 ± 0.7 | 52.2 ± 2.1 | 51.6 ± 3.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitao, M.; Agathokleous, E.; Yazaki, K.; Komatsu, M.; Kitaoka, S.; Tobita, H. Growth and Photosynthetic Responses of Seedlings of Japanese White Birch, a Fast-Growing Pioneer Species, to Free-Air Elevated O3 and CO2. Forests 2021, 12, 675. https://doi.org/10.3390/f12060675
Kitao M, Agathokleous E, Yazaki K, Komatsu M, Kitaoka S, Tobita H. Growth and Photosynthetic Responses of Seedlings of Japanese White Birch, a Fast-Growing Pioneer Species, to Free-Air Elevated O3 and CO2. Forests. 2021; 12(6):675. https://doi.org/10.3390/f12060675
Chicago/Turabian StyleKitao, Mitsutoshi, Evgenios Agathokleous, Kenichi Yazaki, Masabumi Komatsu, Satoshi Kitaoka, and Hiroyuki Tobita. 2021. "Growth and Photosynthetic Responses of Seedlings of Japanese White Birch, a Fast-Growing Pioneer Species, to Free-Air Elevated O3 and CO2" Forests 12, no. 6: 675. https://doi.org/10.3390/f12060675
APA StyleKitao, M., Agathokleous, E., Yazaki, K., Komatsu, M., Kitaoka, S., & Tobita, H. (2021). Growth and Photosynthetic Responses of Seedlings of Japanese White Birch, a Fast-Growing Pioneer Species, to Free-Air Elevated O3 and CO2. Forests, 12(6), 675. https://doi.org/10.3390/f12060675