Effects of Residue Retention and Removal Following Thinning on Soil Bacterial Community Composition and Diversity in a Larix olgensis Plantation, Northeast China

Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design and Soil Sample Collection
2.3. Soil Physical and Chemical Properties and Biological Analysis
2.3.1. Physicochemical Analysis
2.3.2. DNA Extraction and PCR Amplification
2.4. Bioinformatics and Statistical Analyses
3. Results
3.1. Soil Chemical and Physical Properties
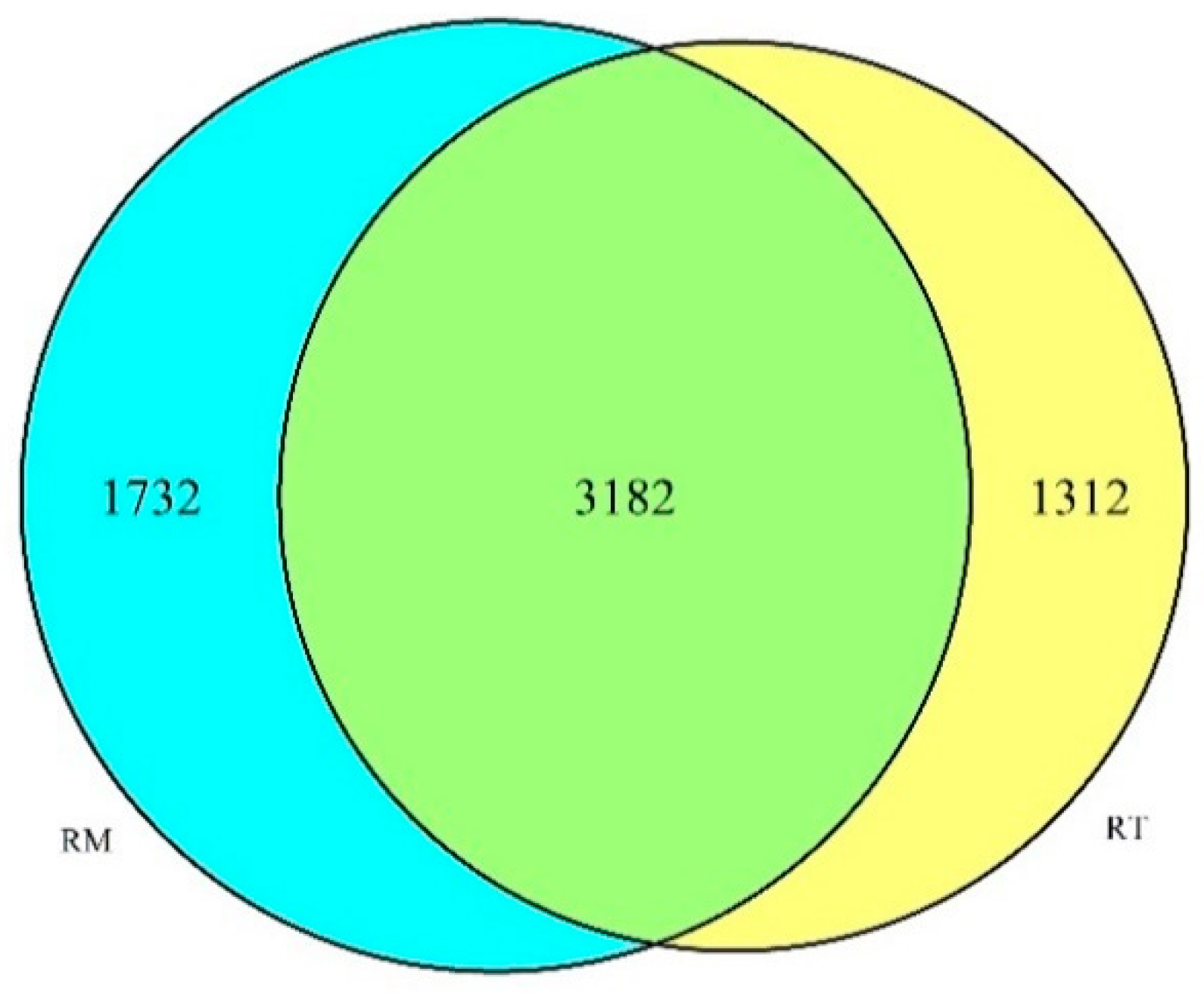
3.2. OTU Statistics and Bacterial Community Diversity
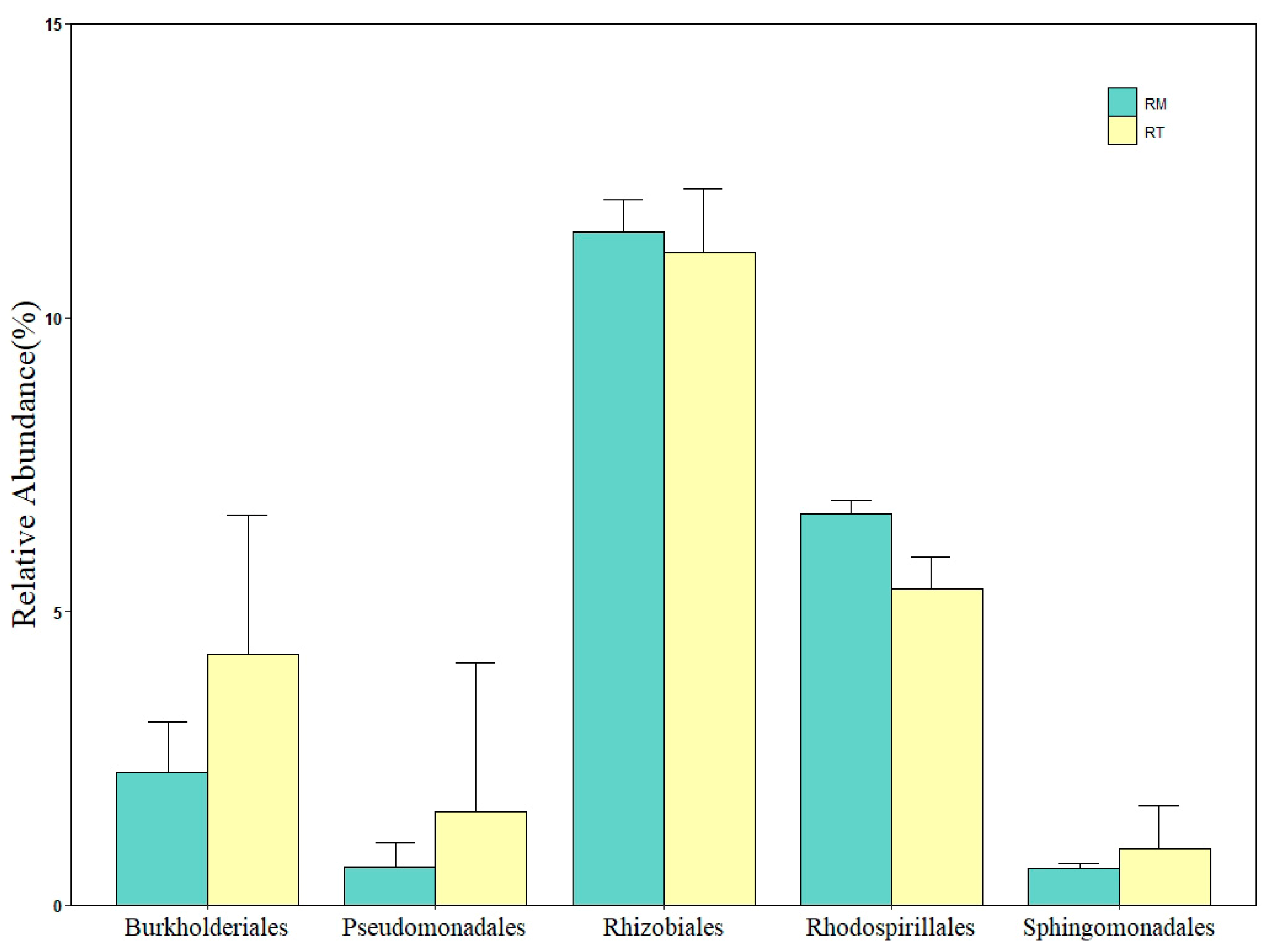
3.3. Composition and Soil Bacterial Community Differences between Different Thinning Residue Treatments
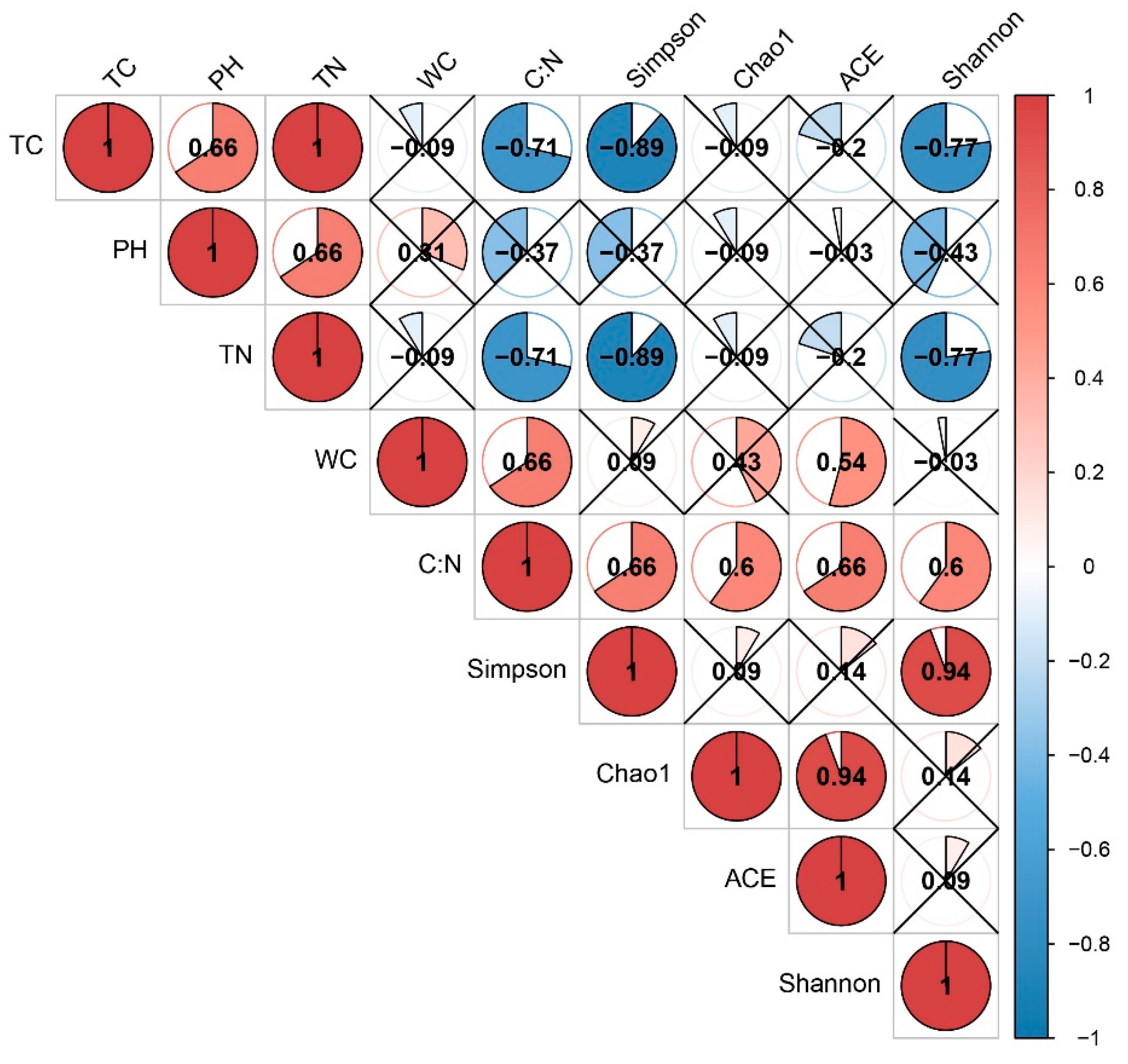
3.4. Comparison of Soil Bacterial Community Structure and Its Relationship with Environmental Factors
4. Discussion
4.1. Response of Soil Bacterial Community Composition to the Different Thinning Residue Treatments
4.2. Effects of Different Thinning Residue Treatments on Soil Bacterial α-Diversity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lei, X.; Lu, Y.; Peng, C.; Zhang, X.; Chang, J.; Hong, L. Growth and structure development of semi-natural larch-spruce-fir (Larix olgensis-Picea jezoensis-Abies nephrolepis) forests in northeast China: 12-year results after thinning. For. Ecol. Manag. 2007, 240, 165–177. [Google Scholar] [CrossRef]
- Smolander, A.; Kitunen, V.; Tamminen, P.; Kukkola, M. Removal of logging residue in Norway spruce thinning stands: Long-term changes in organic layer properties. Soil Biol. Biochem. 2010, 42, 1222–1228. [Google Scholar] [CrossRef]
- Rocha, J.H.T.; Gonçalves, J.L.d.M.; Brandani, C.B.; de Ferraz, A.V.; Franci, A.F.; Marques, E.R.G.; Arthur Junior, J.C.; Hubner, A. Forest residue removal decreases soil quality and affects wood productivity even with high rates of fertilizer application. For. Ecol. Manag. 2018, 430, 188–195. [Google Scholar] [CrossRef]
- Tamminen, P.; Saarsalmi, A.; Smolander, A.; Kukkola, M.; Helmisaari, H.S. Effects of logging residue harvest in thinnings on amounts of soil carbon and nutrients in Scots pine and Norway spruce stands. For. Ecol. Manag. 2012, 263, 31–38. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Jin, Y.; Sun, Z. Impacts of thinning on soil carbon and nutrients and related extracellular enzymes in a larch plantation. For. Ecol. Manag. 2019, 450, 117523. [Google Scholar] [CrossRef]
- García-Oliva, F.; Sveshtarova, B.; Oliva, M. Seasonal effects on soil organic carbon dynamics in a tropical deciduous forest ecosystem in western Mexico. J. Trop. Ecol. 2003, 19, 179–188. [Google Scholar] [CrossRef]
- Semenov, A.V.; Pereira e Silva, M.C.; Szturc-Koestsier, A.E.; Schmitt, H.; Falcão Salles, J.; Van Elsas, J.D. Impact of incorporated fresh 13C potato tissues on the bacterial and fungal community composition of soil. Soil Biol. Biochem. 2012, 49, 88–95. [Google Scholar] [CrossRef]
- Wang, J.J.; Li, X.Y.; Zhu, A.N.; Zhang, X.K.; Zhang, H.W.; Liang, W.J. Effects of tillage and residue management on soil microbial communities in north china. Plant Soil Environ. 2012, 58, 28–33. [Google Scholar] [CrossRef]
- Leff, J.W.; Wieder, W.R.; Taylor, P.G.; Townsend, A.R.; Nemergut, D.R.; Grandy, A.S.; Cleveland, C.C. Experimental litterfall manipulation drives large and rapid changes in soil carbon cycling in a wet tropical forest. Glob. Chang. Biol. 2012, 18, 2969–2979. [Google Scholar] [CrossRef]
- Wan, X.H.; Huang, Z.Q.; He, Z.M.; Yu, Z.P.; Wang, M.H.; Liu, R.Q.; Zheng, L.J. Changes of above- and belowground carbon input affected soil microbial biomass and community composition in two tree species plantations in subtropical China. Acta Ecol. Sin. 2016, 36, 3582–3590. [Google Scholar] [CrossRef]
- Jacobson, S.; Kukkola, M.; Mälkönen, E.; Tveite, B. Impact of whole-tree harvesting and compensatory fertilization on growth of coniferous thinning stands. For. Ecol. Manag. 2000, 129, 41–51. [Google Scholar] [CrossRef]
- Smolander, A.; Levula, T.; Kitunen, V. Response of litter decomposition and soil C and N transformations in a Norway spruce thinning stand to removal of logging residue. For. Ecol. Manag. 2008, 256, 1080–1086. [Google Scholar] [CrossRef]
- Rosenberg, O.; Jacobson, S. Effects of repeated slash removal in thinned stands on soil chemistry and understorey vegetation. Silva Fenn. 2004, 38, 133–142. [Google Scholar] [CrossRef][Green Version]
- Chen, C.R.; Xu, Z.H. Soil carbon and nitrogen pools and microbial properties in a 6-year-old slash pine plantation of subtropical Australia: Impacts of harvest residue management. For. Ecol. Manag. 2005, 206, 237–247. [Google Scholar] [CrossRef]
- Tutua, S.; Zhang, Y.; Xu, Z.; Blumfield, T. Residue retention mitigated short-term adverse effect of clear-cutting on soil carbon and nitrogen dynamics in subtropical Australia. J. Soils Sediments 2019, 19, 3786–3796. [Google Scholar] [CrossRef]
- Nave, L.E.; Vance, E.D.; Swanston, C.W.; Curtis, P.S. Harvest impacts on soil carbon storage in temperate forests. For. Ecol. Manag. 2010, 259, 857–866. [Google Scholar] [CrossRef]
- Mathers, N.J.; Xu, Z. Solid-state 13C NMR spectroscopy: Characterization of soil organic matter under two contrasting residue management regimes in a 2-year-old pine plantation of subtropical Australia. Geoderma 2003, 114, 19–31. [Google Scholar] [CrossRef]
- Blumfield, T.J.; Xu, Z.H.; Saffigna, P.G. Carbon and nitrogen dynamics under windrowed residues during the establishment phase of a second-rotation hoop pine plantation in subtropical Australia. For. Ecol. Manag. 2004, 200, 279–291. [Google Scholar] [CrossRef]
- Powers, R.F.; Scott, D.A.; Sanchez, F.G.; Voldseth, R.A.; Page-Dumroese, D.; Elioff, J.D.; Stone, D.M. The North American long-term soil productivity experiment: Findings from the first decade of research. For. Ecol. Manag. 2005, 220, 31–50. [Google Scholar] [CrossRef]
- Smaill, S.J.; Clinton, P.W.; Greenfield, L.G. Postharvest organic matter removal effects on FH layer and mineral soil characteristics in four New Zealand Pinus radiata plantations. For. Ecol. Manag. 2008, 256, 558–563. [Google Scholar] [CrossRef]
- Huang, Z.; Clinton, P.W.; Davis, M.R. Post-harvest residue management effects on recalcitrant carbon pools and plant biomarkers within the soil heavy fraction in Pinus radiata plantations. Soil Biol. Biochem. 2011, 43, 404–412. [Google Scholar] [CrossRef]
- Kumaraswamy, S.; Mendham, D.S.; Grove, T.S.; O’Connell, A.M.; Sankaran, K.V.; Rance, S.J. Harvest residue effects on soil organic matter, nutrients and microbial biomass in eucalypt plantations in Kerala, India. For. Ecol. Manag. 2014, 328, 140–149. [Google Scholar] [CrossRef]
- Lull, C.; Bautista, I.; Lidón, A.; del Campo, A.D.; González-Sanchis, M.; García-Prats, A. Temporal effects of thinning on soil organic carbon pools, basal respiration and enzyme activities in a Mediterranean Holm oak forest. For. Ecol. Manag. 2020, 464, 118088. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, M.; Tang, L.; Che, R.; Chen, H.; Blumfield, T.; Boyd, S.; Nouansyvong, M.; Xu, Z. Long-Term Harvest Residue Retention Could Decrease Soil Bacterial Diversities Probably Due to Favouring Oligotrophic Lineages. Microb. Ecol. 2018, 76, 771–781. [Google Scholar] [CrossRef]
- Fernandez, A.L.; Sheaffer, C.C.; Wyse, D.L.; Staley, C.; Gould, T.J.; Sadowsky, M.J. Structure of bacterial communities in soil following cover crop and organic fertilizer incorporation. Appl. Microbiol. Biotechnol. 2016, 100, 9331–9341. [Google Scholar] [CrossRef]
- Fernandez, A.L.; Sheaffer, C.C.; Wyse, D.L.; Staley, C.; Gould, T.J.; Sadowsky, M.J. Associations between soil bacterial community structure and nutrient cycling functions in long-term organic farm soils following cover crop and organic fertilizer amendment. Sci. Total Environ. 2016, 566–567, 949–959. [Google Scholar] [CrossRef]
- Mendham, D.S.; O’Connell, A.M.; Grove, T.S.; Rance, S.J. Residue management effects on soil carbon and nutrient contents and growth of second rotation eucalypts. For. Ecol. Manag. 2003, 181, 357–372. [Google Scholar] [CrossRef]
- De la Cruz-Barrón, M.; Cruz-Mendoza, A.; Navarro–Noya, Y.E.; Ruiz-Valdiviezo, V.M.; Ortíz-Gutiérrez, D.; Ramírez-Villanueva, D.A.; Luna-Guido, M.; Thierfelder, C.; Wall, P.C.; Verhulst, N.; et al. The Bacterial Community Structure and Dynamics of Carbon and Nitrogen when Maize (Zea mays L.) and Its Neutral Detergent Fibre Were Added to Soil from Zimbabwe with Contrasting Management Practices. Microb. Ecol. 2017, 73, 135–152. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.S.; Garrett, L.G.; Beets, P.N.; Kimberley, M.O.; Oliver, G.R. Impacts of Harvest Residue Management on Soil Carbon Stocks in a Plantation Forest. Soil Sci. Soc. Am. J. 2008, 72, 1621–1627. [Google Scholar] [CrossRef]
- Hannam, K.D.; Quideau, S.A.; Kishchuk, B.E. Forest floor microbial communities in relation to stand composition and timber harvesting in northern Alberta. Soil Biol. Biochem. 2006, 38, 2565–2575. [Google Scholar] [CrossRef]
- Zechmeister-Boltenstern, S.; Keiblinger, K.M.; Mooshammer, M.; Peñuelas, J.; Richter, A.; Sardans, J.; Wanek, W. The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecol. Monogr. 2015, 85, 133–155. [Google Scholar] [CrossRef]
- Månsson, K.; Bengtson, P.; Falkengren-Grerup, U.; Bengtsson, G. Plant-microbial competition for nitrogen uncoupled from soil C:N ratios. Oikos 2009, 118, 1908–1916. [Google Scholar] [CrossRef]
- Luiro, J.; Kukkola, M.; Saarsalmi, A.; Tamminen, P.; Helmisaari, H.S. Logging residue removal after thinning in boreal forests: Long-term impact on the nutrient status of Norway spruce and Scots pine needles. Tree Physiol. 2010, 30, 78–88. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Helmisaari, H.S.; Hanssen, K.H.; Jacobson, S.; Kukkola, M.; Luiro, J.; Saarsalmi, A.; Tamminen, P.; Tveite, B. Logging residue removal after thinning in Nordic boreal forests: Long-term impact on tree growth. For. Ecol. Manag. 2011, 261, 1919–1927. [Google Scholar] [CrossRef]
- Qiu, J.; Cao, J.; Lan, G.; Liang, Y.; Wang, H.; Li, Q. The influence of land use patterns on soil bacterial community structure in the karst graben basin of Yunnan province, China. Forests 2020, 11, 51. [Google Scholar] [CrossRef]
- Wu, S.J.; Deng, J.J.; Yin, Y.; Qin, S.J.; Zhu, W.X.; Zhou, Y.B.; Wang, B.; Ruan, H.; Jin, L. Bacterial community changes associated with land use type in the forest montane region of northeast China. Forests 2020, 11, 40. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, S.; Meng, X.; Li, M.; Mu, L.; Lei, J.; Sui, X. Conversion from natural wetlands to forestland and farmland alters the composition of soil fungal communities in Sanjiang Plain, Northeast China. Biotechnol. Biotechnol. Equip. 2018, 32, 951–960. [Google Scholar] [CrossRef]
- Mi, L.; Wang, G.; Jin, J.; Sui, Y.; Liu, J.; Liu, X. Comparison of microbial community structures in four black soils along a climatic gradient in northeast China. Can. J. Soil Sci. 2012, 92, 543–549. [Google Scholar] [CrossRef]
- Chen, D.; Huang, X.; Zhang, S.; Sun, X. Biomass modeling of larch (Larix spp.) plantations in China based on the mixed model, dummy variable model, and Bayesian hierarchical model. Forests 2017, 8, 268. [Google Scholar] [CrossRef]
- Yang, K.; Zhu, J.; Xu, S.; Zheng, X. Conversion from temperate secondary forests into plantations (Larix spp.): Impact on belowground carbon and nutrient pools in northeastern China. L. Degrad. Dev. 2018, 29, 4129–4139. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Carbon and organic matter. In Methods of Soil Analysis-Part 2: Chemical and Microbiological Properties; Page, A.L., Mille, R.H., Eds.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 561–579. [Google Scholar]
- Thomas, R.L.; Sheard, R.W.; Moyer, J.R. Comparison of Conventional and Automated Procedures for Nitrogen, Phosphorus, and Potassium Analysis of Plant Material Using a Single Digestion. Agron. J. 1967, 59, 240–243. [Google Scholar] [CrossRef]
- Wilke, B. Determination of chemical and physical soil properties. In Monitoring and Assessing Soil Bioremediation; Springer: Berlin/Heidelberg, Germany, 2005; pp. 47–95. [Google Scholar]
- Aneja, M.K.; Sharma, S.; Fleischmann, F.; Stich, S.; Heller, W.; Bahnweg, G.; Munch, J.C.; Schloter, M. Microbial colonization of beech and spruce litter—Influence of decomposition site and plant litter species on the diversity of microbial community. Microb. Ecol. 2006, 52, 127–135. [Google Scholar] [CrossRef]
- Li, Y.; Bezemer, T.M.; Yang, J.; Lü, X.; Li, X.; Liang, W.; Han, X.; Li, Q. Changes in litter quality induced by N deposition alter soil microbial communities. Soil Biol. Biochem. 2019, 130, 33–42. [Google Scholar] [CrossRef]
- Wallenstein, M.D.; McMahon, S.; Schimel, J. Bacterial and fungal community structure in Arctic tundra tussock and shrub soils. FEMS Microbiol. Ecol. 2007, 59, 428–435. [Google Scholar] [CrossRef]
- Navarro-Noya, Y.E.; Gómez-Acata, S.; Montoya-Ciriaco, N.; Rojas-Valdez, A.; Suárez-Arriaga, M.C.; Valenzuela-Encinas, C.; Jiménez-Bueno, N.; Verhulst, N.; Govaerts, B.; Dendooven, L. Relative impacts of tillage, residue management and crop-rotation on soil bacterial communities in a semi-arid agroecosystem. Soil Biol. Biochem. 2013, 65, 86–95. [Google Scholar] [CrossRef]
- Im, W.T.; Kim, S.H.; Kim, M.K.; Ten, L.N.; Lee, S.T. Pleomorphomonas koreensis sp. nov., a nitrogen-fixing species in the order Rhizobiales. Int. J. Syst. Evol. Microbiol. 2006, 56, 1663–1666. [Google Scholar] [CrossRef]
- Yarwood, S.A.; Myrold, D.D.; Högberg, M.N. Termination of belowground C allocation by trees alters soil fungal and bacterial communities in a boreal forest. FEMS Microbiol. Ecol. 2009, 70, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cong, J.; Lu, H.; Li, G.; Qu, Y.; Su, X.; Zhou, J.; Li, D. Community structure and elevational diversity patterns of soil Acidobacteria. J. Environ. Sci. 2014, 26, 1717–1724. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Krankina, O.N.; Harmon, M.E.; Griazkin, A.V. Nutrient stores and dynamics of woody detritus in a boreall forest: Modeling potential implications at the stand level. Can. J. For. Res. 1999, 29, 20–32. [Google Scholar] [CrossRef]
- Hyvönen, R.; Olsson, B.A.; Lundkvist, H.; Staaf, H. Decomposition and nutrient release from Picea abies (L.) Karst. and Pinus sylvestris L. logging residues. For. Ecol. Manag. 2000, 126, 97–112. [Google Scholar] [CrossRef]
- Zheng, M.; Zhou, Z.; Luo, Y.; Zhao, P.; Mo, J. Global pattern and controls of biological nitrogen fixation under nutrient enrichment: A meta-analysis. Glob. Chang. Biol. 2019, 25, 3018–3030. [Google Scholar] [CrossRef] [PubMed]
- Ballard, T.M. Impacts of forest management on northern forest soils. For. Ecol. Manag. 2000, 133, 37–42. [Google Scholar] [CrossRef]
- Hartmann, M.; Howes, C.G.; Vaninsberghe, D.; Yu, H.; Bachar, D.; Christen, R.; Henrik Nilsson, R.; Hallam, S.J.; Mohn, W.W. Significant and persistent impact of timber harvesting on soil microbial communities in Northern coniferous forests. ISME J. 2012, 6, 2199–2218. [Google Scholar] [CrossRef]
- Su, P.; Lou, J.; Brookes, P.C.; Luo, Y.; He, Y.; Xu, J. Taxon-specific responses of soil microbial communities to different soil priming effects induced by addition of plant residues and their biochars. J. Soils Sedim. 2017, 17, 674–684. [Google Scholar] [CrossRef]
- Che, R.; Liu, D.; Qin, J.; Wang, F.; Wang, W.; Xu, Z.; Li, L.; Hu, J.; Tahmasbian, I.; Cui, X. Increased litter input significantly changed the total and active microbial communities in degraded grassland soils. J. Soils Sediments 2020, 20, 2804–2816. [Google Scholar] [CrossRef]
- Yao, H.; He, Z.; Wilson, M.J.; Campbell, C.D. Microbial biomass and community structure in a sequence of soils with increasing fertility and changing land use. Microb. Ecol. 2000. [Google Scholar] [CrossRef]
- Zeng, Q.; An, S.; Liu, Y.; Wang, H.; Wang, Y. Biogeography and the driving factors affecting forest soil bacteria in an arid area. Sci. Total Environ. 2019, 680, 124–131. [Google Scholar] [CrossRef]
- Meng, H.; Li, K.; Nie, M.; Wan, J.R.; Quan, Z.X.; Fang, C.M.; Chen, J.K.; Gu, J.D.; Li, B. Responses of bacterial and fungal communities to an elevation gradient in a subtropical montane forest of China. Appl. Microbiol. Biotechnol. 2013, 97, 2219–2230. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Lin, J.; Guo, X.; Liu, X.; Wu, J.; Zhao, Y.; Zhang, J. Impacts of forest conversion on soil bacterial community composition and diversity in subtropical forests. Catena 2019, 175, 167–173. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Luo, Y. Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ceja-Navarro, J.A.; Rivera, F.N.; Patiño-Zúñiga, L.; Govaerts, B.; Marsch, R.; Vila-Sanjurjo, A.; Dendooven, L. Molecular characterization of soil bacterial communities in contrasting zero tillage systems. Plant Soil 2010, 329, 127–137. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TC | TN | pH | C:N | WC | |
|---|---|---|---|---|---|
| g/kg | g/kg | % | |||
| RT | 51.42 ± 3.86 | 4.46 ± 0.34 | 5.83 ± 0.10 | 11.52 ± 0.12 | 31.08 ± 3.33 |
| RM | 38.72 ± 1.92 | 3.30 ± 0.26 | 5.76 ± 0.11 | 11.75 ± 0.34 | 30.15 ± 2.43 |
| p-value | 0.007 | 0.009 | 0.255 | 0.317 | 0.715 |
| Simpson | Chao1 | ACE | Shannon | |
|---|---|---|---|---|
| RT | 0.9949 ± 0.03 | 2792.84 ± 683.03 | 2798.22 ± 697.32 | 9.67 ± 0.52 |
| RM | 0.9969 ± 0.01 | 2932.78 ± 256.66 | 2976.28 ± 317.56 | 10.07 ± 0.11 |
| p-value | 0.270 | 0.756 | 0.708 | 0.253 |
| Soil Environmental Factor | RDA1 | RDA2 | r2 | Pr (>r) |
|---|---|---|---|---|
| TC | 0.9907 | 0.1362 | 0.9700 | 0.0041 ** |
| TN | 0.9776 | 0.2104 | 0.9798 | 0.0055 ** |
| C:N | −0.6029 | −0.7978 | 0.8712 | 0.0514 |
| pH | 0.6577 | 0.7533 | 0.3531 | 0.5042 |
| WC | −0.0786 | −0.9969 | 0.3779 | 0.5236 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, X.; Du, X.; Sun, Z.; Chen, X. Effects of Residue Retention and Removal Following Thinning on Soil Bacterial Community Composition and Diversity in a Larix olgensis Plantation, Northeast China. Forests 2021, 12, 559. https://doi.org/10.3390/f12050559
Dong X, Du X, Sun Z, Chen X. Effects of Residue Retention and Removal Following Thinning on Soil Bacterial Community Composition and Diversity in a Larix olgensis Plantation, Northeast China. Forests. 2021; 12(5):559. https://doi.org/10.3390/f12050559
Chicago/Turabian StyleDong, Xue, Xin Du, Zhihu Sun, and Xiangwei Chen. 2021. "Effects of Residue Retention and Removal Following Thinning on Soil Bacterial Community Composition and Diversity in a Larix olgensis Plantation, Northeast China" Forests 12, no. 5: 559. https://doi.org/10.3390/f12050559
APA StyleDong, X., Du, X., Sun, Z., & Chen, X. (2021). Effects of Residue Retention and Removal Following Thinning on Soil Bacterial Community Composition and Diversity in a Larix olgensis Plantation, Northeast China. Forests, 12(5), 559. https://doi.org/10.3390/f12050559