
Root Growth Was Enhanced in China Fir (Cunninghamia lanceolata) after Mechanical Disturbance by Ice Storm

Abstract
:1. Introduction
2. Materials and Methods
2.1. The Experiment Site
2.2. The Studied Species
2.3. Tree Biomass Measurement
2.4. Allometric Model Development and Evaluation
3. Results
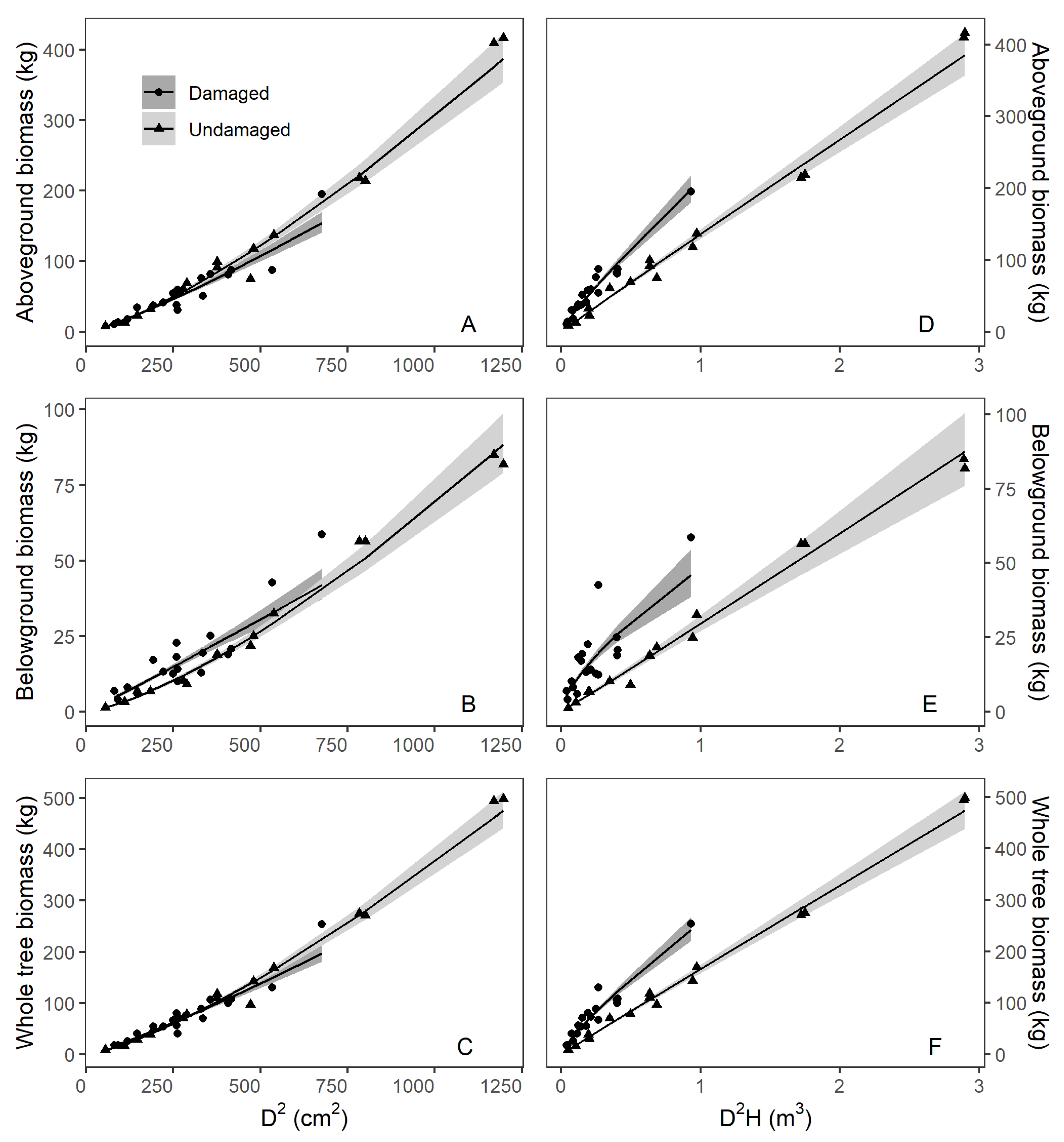
3.1. Allometric Models of Biomass Estimation for Different Components of China Fir with Different Variables
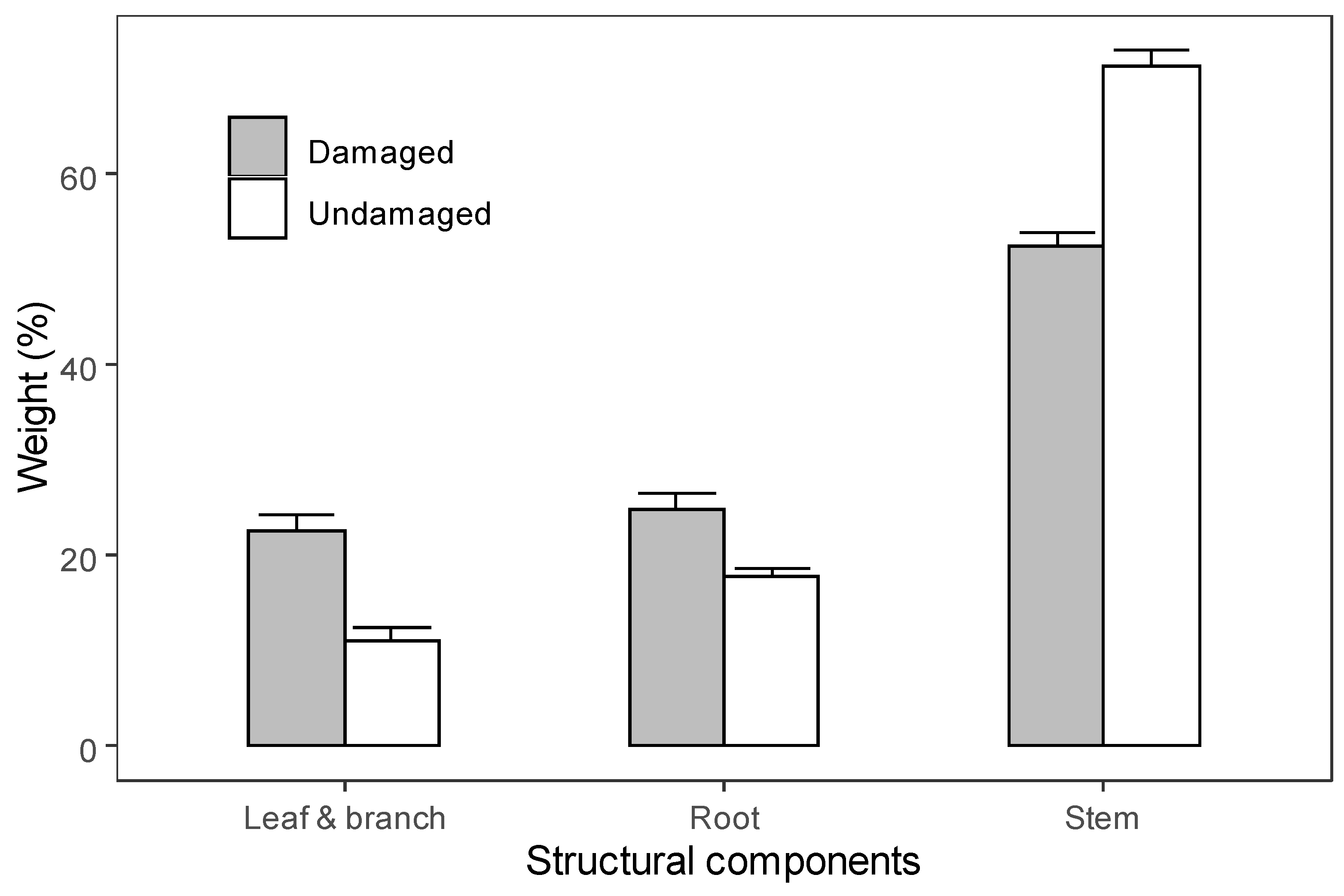
3.2. Biomass Allocations
4. Discussion
4.1. Comparison between Biomass Allometric Models with Different Independent Viriable
4.2. The Impact of Ice Storm on Damaged China Fir
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Houghton, R.A. Aboveground Forest Biomass and the Global Carbon Balance. Glob. Chang. Biol. 2005, 11, 945–958. [Google Scholar] [CrossRef]
- Yu, G.; Chen, Z.; Piao, S.; Peng, C.; Ciais, P.; Wang, Q.; Li, X.; Zhu, X. High carbon dioxide uptake by subtropical forest ecosystems in the East Asian monsoon region. Proc. Natl. Acad. Sci. USA 2014, 111, 4910–4915. [Google Scholar] [CrossRef] [Green Version]
- Brown, S. Measuring carbon in forests: Current status and future challenges. Environ. Pollut. 2002, 116, 363–372. [Google Scholar] [CrossRef]
- Tang, S.; Zhang, H.; Xu, H. Study on Establish and Estimate Method of Compatible Biomass Model. Sci. Silvae Sin. 2000, 36, 19–27. [Google Scholar] [CrossRef]
- Xie, Y.; Sha, Z.; Yu, M. Remote sensing imagery in vegetation mapping: A review. J. Plant Ecol. 2008, 1, 9–23. [Google Scholar] [CrossRef]
- Hossain, M.; Saha, C.; Abdullah, S.M.R.; Saha, S.; Siddique, M.R.H. Allometric biomass, nutrient and carbon stock models for Kandelia candel of the Sundarbans, Bangladesh. Trees 2016, 30, 709–717. [Google Scholar] [CrossRef]
- Hou, Y.-N.; Wu, H.-L. Using Nonlinear Regression Method to Develop Allometric Equations for Aboveground Biomass Estimate of Three Evergreen Broadleaved Tree Species in Subtropical China. J. Cent. South Univ. For. Technol. 2016. 36, 98–101. [CrossRef]
- Ubuy, M.H.; Eid, T.; Bollandsås, O.M.; Birhane, E. Aboveground biomass models for trees and shrubs of exclosures in the drylands of Tigray, northern Ethiopia. J. Arid. Environ. 2018, 156, 9–18. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Roxburgh, S.H.; Paul, K.I.; Clifford, D.; England, J.R.; Raison, R.J. Guidelines for Constructing Allometric Models for the Prediction of Woody Biomass: How Many Individuals to Harvest? Ecosphere 2015, 6, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Basuki, T.M.; van Laake, P.E.; Skidmore, A.K.; Hussin, Y.A. Allometric equations for estimating the above-ground biomass in tropical lowland Dipterocarp forests. For. Ecol. Manag. 2009, 257, 1684–1694. [Google Scholar] [CrossRef]
- Nafus, A.M.; McClaran, M.P.; Archer, S.R.; Throop, H.L. Multispecies Allometric Models Predict Grass Biomass in Semidesert Rangeland. Rangel. Ecol. Manag. 2009, 62, 68–72. [Google Scholar] [CrossRef]
- Melson, S.L.; Harmon, M.E.; Fried, J.S.; Domingo, J.B. Estimates of live-tree carbon stores in the Pacific Northwest are sensitive to model selection. Carbon Balance Manag. 2011, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaturvedi, R.K.; Raghubanshi, A. Allometric Models for Accurate Estimation of Aboveground Biomass of Teak in Tropical Dry Forests of India. For. Sci. 2015, 61, 938–949. [Google Scholar] [CrossRef]
- Xie, T.-T.; Li, G.; Zhou, G.-Y.; Wu, Z.-M.; Zhao, H.-B.; Qiu, Z.-J.; Liang, R.-Y. Aboveground biomass of natural Castanopsis carlesii-Schima superba community in Xiaokeng of Nanling Mountains, South China. Chin. J. Appl. Ecol. 2013, 24, 2399–2407. [Google Scholar]
- Zuo, S.-D.; Ren, Y.; Weng, X.; Ding, H.-F.; Luo, Y.-J. Biomass allometric equations of nine common tree species in an evergreen broadleaved forest of subtropical China. Chin. J. Appl. Ecol. 2015, 26, 356–362. [Google Scholar]
- Lin, K.; Lyu, M.; Jiang, M.; Chen, Y.; Li, Y.; Chen, G.; Xie, J.; Yang, Y. Improved allometric equations for estimating biomass of the three Castanopsis carlesii H. forest types in subtropical China. New For. 2017, 48, 115–135. [Google Scholar] [CrossRef]
- Peng, S.; He, N.; Yu, G.; Wang, Q. Aboveground biomass estimation at different scales for subtropical forests in China. Bot. Stud. 2017, 58, 45. [Google Scholar] [CrossRef] [Green Version]
- Addo-Danso, S.D.; Prescott, C.E.; Smith, A.R. Methods for estimating root biomass and production in forest and woodland ecosystem carbon studies: A review. For. Ecol. Manag. 2016, 359, 332–351. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, J.; Franklin, S.B.; Liang, J.; Ding, P.; Luo, Y.; Lu, Z.; Bao, D.; Jiang, M. Improving allometry models to estimate the above- and belowground biomass of subtropical forest, China. Ecosphere 2015, 6, art289. [Google Scholar] [CrossRef]
- Masson-Delmotte, V.; Zhai, P.; Pirani, A.; Connors, S.L.; Péan, C.; Berger, S.; Caud, N.; Chen, Y.; Goldfarb, L.; Gomis, M.I.; et al. (Eds.) Climate Change 2021: The Physical Science Basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Felton, A.J.; Smith, M.D. Integrating plant ecological responses to climate extremes from individual to ecosystem levels. Philos. Trans. R. Soc. B: Biol. Sci. 2017, 372, 20160142. [Google Scholar] [CrossRef] [Green Version]
- Suarez, M.L.; Kitzberger, T. Differential effects of climate variability on forest dynamics along a precipitation gradient in northern Patagonia. J. Ecol. 2010, 98, 1023–1034. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Tanner, E.V.J.; Healey, J.R. Damage and Responsiveness of Jamaican Montane Tree Species after Disturbance by a Hurricane. Ecology 1995, 76, 2562–2580. [Google Scholar] [CrossRef]
- Renton, K.; Salinas-Melgoza, A.; Rueda-Hernández, R.; Vázquez-Reyes, L.D. Differential resilience to extreme climate events of tree phenology and cavity resources in tropical dry forest: Cascading effects on a threatened species. For. Ecol. Manag. 2018, 426, 164–175. [Google Scholar] [CrossRef]
- Belsky, A.J.; Carson, W.P.; Jensen, C.L.; Fox, G.A. Overcompensation by plants: Herbivore optimization or red herring? Evol. Ecol. 1993, 7, 109–121. [Google Scholar] [CrossRef]
- Wang, X. Effects of Ice Storm on the Structure of Evergreen Broad-Leaved Forest in Mt. Nanling; Chinese Academy of Forestry: Beijing, China, 2012. [Google Scholar]
- Xu, Y.W.; Wu, K.K.; Zhu, L.R.; Lin, Z.G.; Peng, S.L. A Review of Freezing Rain and Snow Impacts on Forests in Southern China. Ecol. Environ. Sci. 2010, 19, 1485–1494. [Google Scholar]
- Zhou, B.; Wang, X.; Cao, Y.; Ge, X.; Gu, L.; Meng, J. Damage assessment to subtropical forests following the 2008 Chinese ice storm. iForest—Biogeosciences For. 2017, 10, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Moussa, M.; Mahamane, L. Allometric models for estimating aboveground biomass and carbon in Faidherbia albida and Prosopis africana under agroforestry parklands in drylands of Niger. J. For. Res. 2018, 29, 1703–1717. [Google Scholar] [CrossRef]
- Carl, C.; Biber, P.; Landgraf, D.; Buras, A.; Pretzsch, H. Allometric Models to Predict Aboveground Woody Biomass of Black Locust (Robinia pseudoacacia L.) in Short Rotation Coppice in Previous Mining and Agricultural Areas in Germany. Forests 2017, 8, 328. [Google Scholar] [CrossRef] [Green Version]
- Gou, M.; Xiang, W.; Song, T.; Lei, P.; Zhang, S.; Ouyang, S.; Zeng, Y.; Deng, X.; Fang, X.; Wang, K. Allometric Equations for Applying Plot Inventory and Remote Sensing Data to Assess Coarse Root Biomass Energy in Subtropical Forests. BioEnergy Res. 2017, 10, 536–546. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.; Duque, A.; Eid, T.; Fearnside, P.; Goodman, R.C.; et al. Improved allometric models to estimate the aboveground biomass of tropical trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef]
- Ifo, A.S.; Gomat, H.Y.; Wenina, Y.E.M.; Lokegna, D.L.; Nzonzi, O.R.M.; Ngala, G.C.A.; Henry, M.; Boundzanga, G.C.; Jourdain, C.; Picard, N. Carbon Stocks and Tree Allometries in the Savannahs of the Plateau Batéké, Central Africa. For. Ecol. Manag. 2018, 427, 86–95. [Google Scholar] [CrossRef]
- Zhao, H.; Li, Z.; Zhou, G.; Qiu, Z.; Wu, Z. Aboveground Biomass Allometric Models for Evergreen Broad-Leaved Forest Damaged by a Serious Ice Storm in Southern China. Forests 2020, 11, 320. [Google Scholar] [CrossRef] [Green Version]
- Schwachtje, J.; Minchin, P.E.H.; Jahnke, S.; van Dongen, J.T.; Schittko, U.; Baldwin, I.T. SNF1-related kinases allow plants to tolerate herbivory by allocating carbon to roots. Proc. Natl. Acad. Sci. USA 2006, 103, 12935–12940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, L.C.; Eubanks, M.D. Overcompensation for insect herbivory: A review and meta-analysis of the evidence. Ecology 2019, 100, e02585. [Google Scholar] [CrossRef]
- Battles, J.J.; Fahey, T.J. Gap Dynamics Following Forest Decline: A Case Study of Red Spruce Forests. Ecol. Appl. 2000, 10, 760–774. [Google Scholar] [CrossRef]
- Rhoads, A.G.; Hamburg, S.P.; Fahey, T.J.; Siccama, T.G.; Hane, E.N.; Battles, J.; Cogbill, C.; Randall, J.; Wilson, G. Effects of an intense ice storm on the structure of a northern hardwood forest. Can. J. For. Res. 2002, 32, 1763–1775. [Google Scholar] [CrossRef] [Green Version]
- Shipley, B.; Meziane, D. The balanced-growth hypothesis and the allometry of leaf and root biomass allocation. Funct. Ecol. 2002, 16, 326–331. [Google Scholar] [CrossRef]
- Ramula, S.; Paige, K.N.; Lennartsson, T.; Tuomi, J. Overcompensation: A 30-year perspective. Ecology 2019, 100, e02667. [Google Scholar] [CrossRef] [Green Version]
- Peschiutta, M.L.; Scholz, F.G.; Goldstein, G.; Bucci, S.J. Lagged effects of sawfly leaf herbivory on reproductive organs in cherry trees: Overcompensation in flower production reduces quality of fruits and seeds. Basic Appl. Ecol. 2020, 45, 22–30. [Google Scholar] [CrossRef]
- Nagel, T.A.; Firm, D.; Rozenbergar, D.; Kobal, M. Patterns and drivers of ice storm damage in temperate forests of Central Europe. Eur. J. For. Res. 2016, 135, 519–530. [Google Scholar] [CrossRef]
- Klopčič, M.; Poljanec, A.; Dolinar, M.; Kastelec, D.; Bončina, A. Ice-storm damage to trees in mixed Central European forests: Damage patterns, predictors and susceptibility of tree species. Forestry 2019, 93, 430–443. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| (A) | |||||||||
| Structural Components | Impacts of Ice Storm | ||||||||
| a | b | a | B | ||||||
| Aboveground | Damaged | 1.21 ± 0.09 | −2.84 ± 0.52 | 0.87 ± 0.06 | −2.70 ± 0.43 | ||||
| Undamaged | 1.32 ± 0.05 | −3.39 ± 0.31 | 0.98 ± 0.03 | −4.12 ± 0.28 | |||||
| Belowground | Damaged | 1.042 ± 0.13 | −3.06 ± 0.72 | 0.68 ± 0.12 | −2.39 ± 0.72 | ||||
| Undamaged | 1.38 ± 0.05 | −5.29 ± 0.27 | 1.02 ± 0.04 | −6.01 ± 0.27 | |||||
| Whole tree | Damaged | 1.16 ± 0.08 | −2.27 ± 0.44 | 0.82 ± 0.06 | −2.01 ± 0.44 | ||||
| Undamaged | 1.33 ± 0.04 | −3.25 ± 0.26 | 0.99 ± 0.06 | −3.98 ± 0.26 | |||||
| (B) | |||||||||
| Structural Components | Impacts of Ice Storm | Bias | CV | Adj. R2 | |||||
| D | D2H | D | D2H | D | D2H | ||||
| Aboveground | Damaged | −0.00322 | −0.00328 | 0.05728 | 0.04932 | 0.90730 | 0.93127 | ||
| Undamaged | −0.00071 | −0.00094 | 0.03879 | 0.03245 | 0.97889 | 0.98522 | |||
| Belowground | Damaged | −0.01245 | −0.02233 | 0.11289 | 0.14611 | 0.79014 | 0.64843 | ||
| Undamaged | −0.00307 | −0.03274 | 0.05285 | 0.05769 | 0.98497 | 0.98209 | |||
| Whole tree | Damaged | −0.00171 | −0.00269 | 0.04490 | 0.04991 | 0.92686 | 0.90966 | ||
| Undamaged | −0.00027 | −0.00057 | 0.03088 | 0.02505 | 0.98549 | 0.99045 | |||
| Component | Factors | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Sum. Square | Df | F Value | p (>F) | Sum. Square | Df | F Value | p (>F) | ||
| Aboveground | Tree size | 8.106 | 1 | 204.859 | <0.001 | 8.306 | 1 | 288.393 | <0.001 |
| Stem breakage | 0.035 | 1 | 0.877 | 0.357 | 0.221 | 1 | 7.686 | 0.010 | |
| Tree size × Stem breakage | 0.045 | 1 | 1.129 | 0.297 | 0.079 | 1 | 2.752 | 0.108 | |
| Belowground | Tree size | 6.042 | 1 | 98.698 | <0.001 | 5.038 | 1 | 51.539 | <0.001 |
| Stem breakage | 0.559 | 1 | 9.130 | 0.005 | 1.437 | 1 | 14.704 | <0.001 | |
| Tree size × Stem breakage | 0.411 | 1 | 6.710 | 0.015 | 0.806 | 1 | 8.242 | 0.008 | |
| Whole tree | Tree size | 7.448 | 1 | 267.152 | <0.001 | 7.318 | 1 | 249.753 | <0.001 |
| Stem breakage | 0.107 | 1 | 3.856 | 0.059 | 0.424 | 1 | 14.462 | <0.001 | |
| Tree size × Stem breakage | 0.105 | 1 | 3.760 | 0.062 | 0.192 | 1 | 6.565 | 0.160 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Zhao, H.; Zhou, G.; Qiu, Z.; Wang, X.; Wu, Z. Root Growth Was Enhanced in China Fir (Cunninghamia lanceolata) after Mechanical Disturbance by Ice Storm. Forests 2021, 12, 1800. https://doi.org/10.3390/f12121800
Li Z, Zhao H, Zhou G, Qiu Z, Wang X, Wu Z. Root Growth Was Enhanced in China Fir (Cunninghamia lanceolata) after Mechanical Disturbance by Ice Storm. Forests. 2021; 12(12):1800. https://doi.org/10.3390/f12121800
Chicago/Turabian StyleLi, Zhaojia, Houben Zhao, Guangyi Zhou, Zhijun Qiu, Xu Wang, and Zhongmin Wu. 2021. "Root Growth Was Enhanced in China Fir (Cunninghamia lanceolata) after Mechanical Disturbance by Ice Storm" Forests 12, no. 12: 1800. https://doi.org/10.3390/f12121800
APA StyleLi, Z., Zhao, H., Zhou, G., Qiu, Z., Wang, X., & Wu, Z. (2021). Root Growth Was Enhanced in China Fir (Cunninghamia lanceolata) after Mechanical Disturbance by Ice Storm. Forests, 12(12), 1800. https://doi.org/10.3390/f12121800