
Shift from Acquisitive to Conservative Root Resource Acquisition Strategy Associated with Increasing Tree Age: A Case Study of Fraxinus mandshurica



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Trees Selection and Root Sampling
2.3. Root Exudation Measurements
2.4. Root Respiration Measurements
2.5. Measurements of Root Morphological Traits
2.6. Measurements of Soil Properties and Root Nitrogen Concentration
2.7. Data Analysis
3. Results
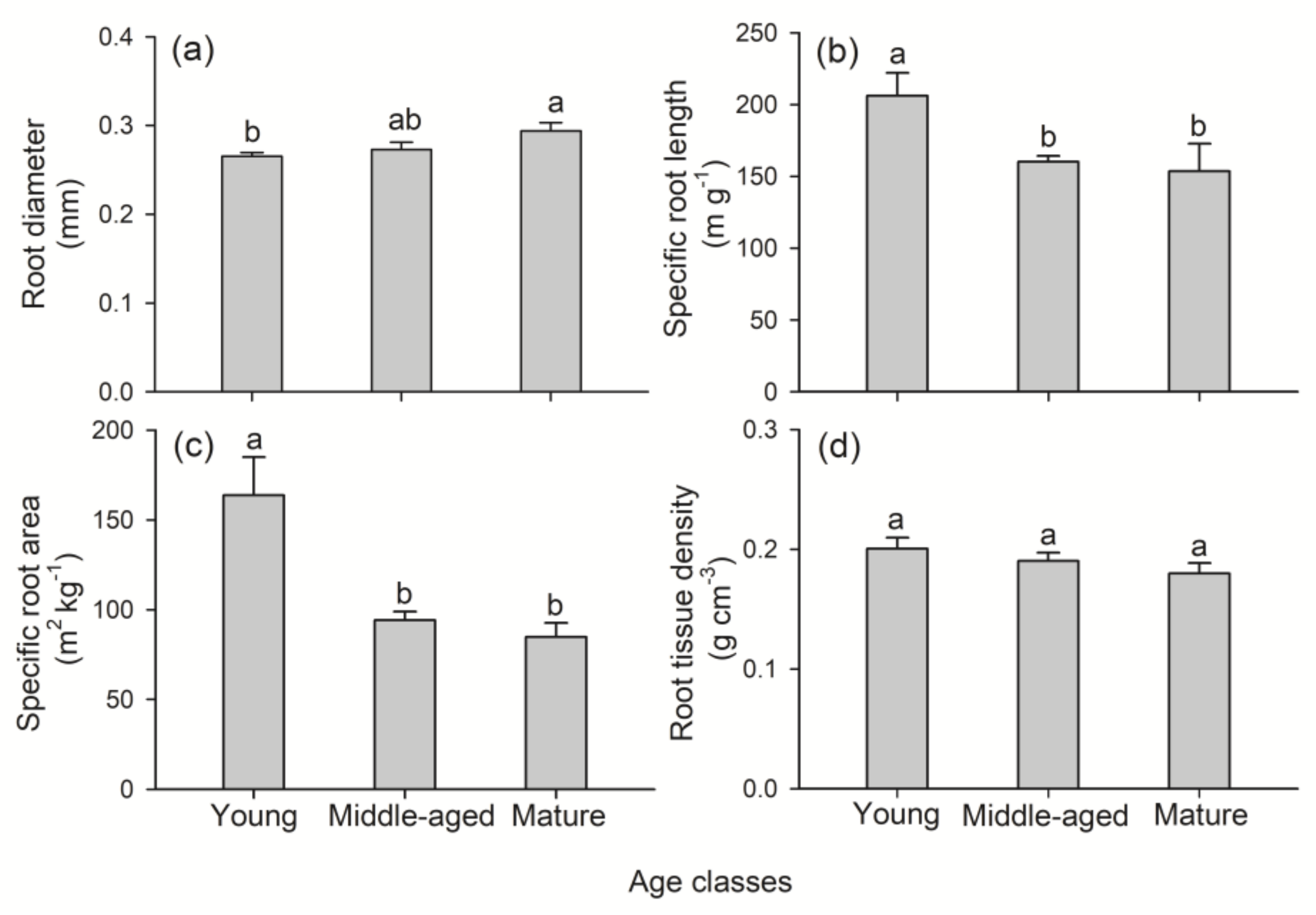
3.1. Root Morphological Traits
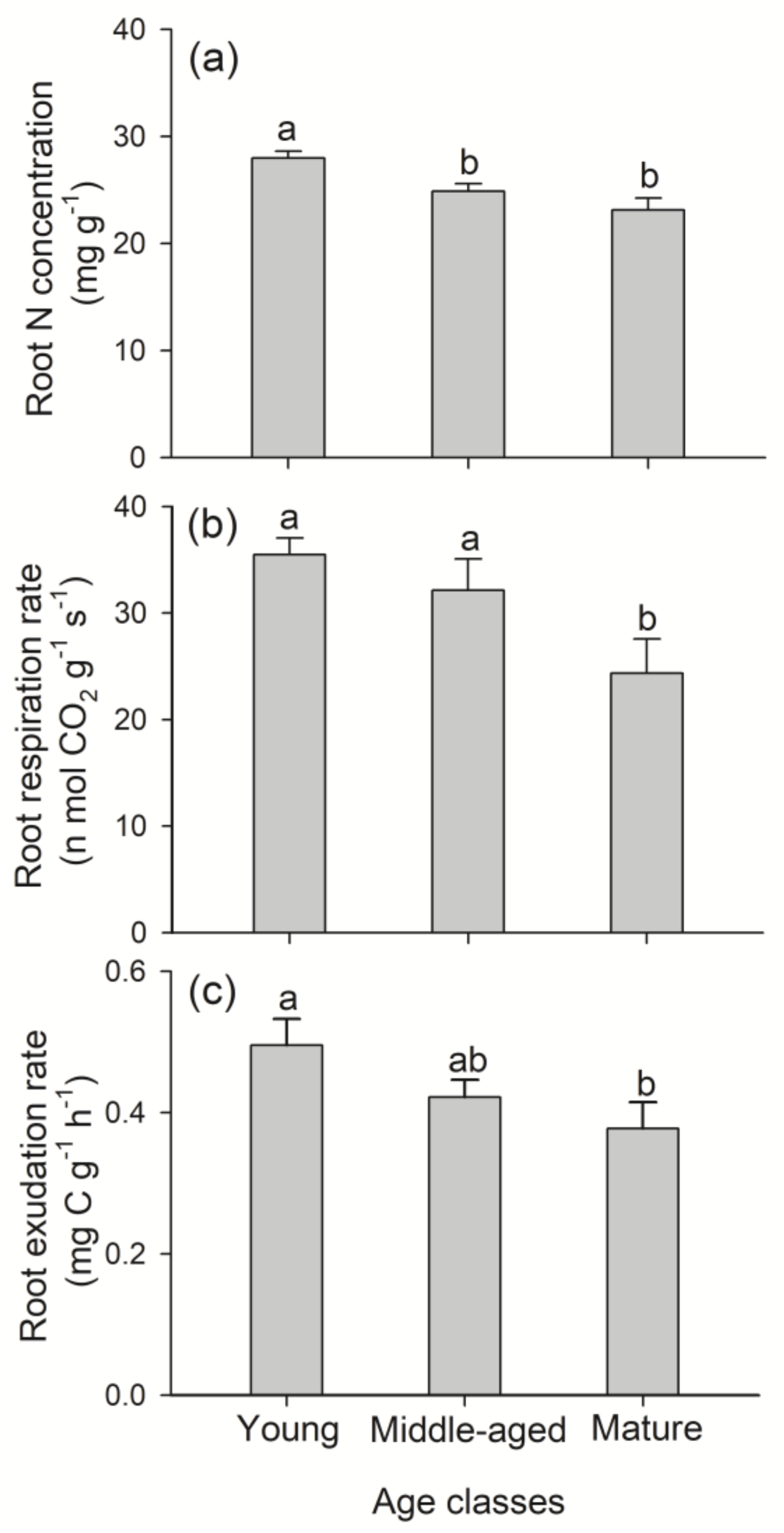
3.2. Root N Concentration, Respiration and Exudation Rates
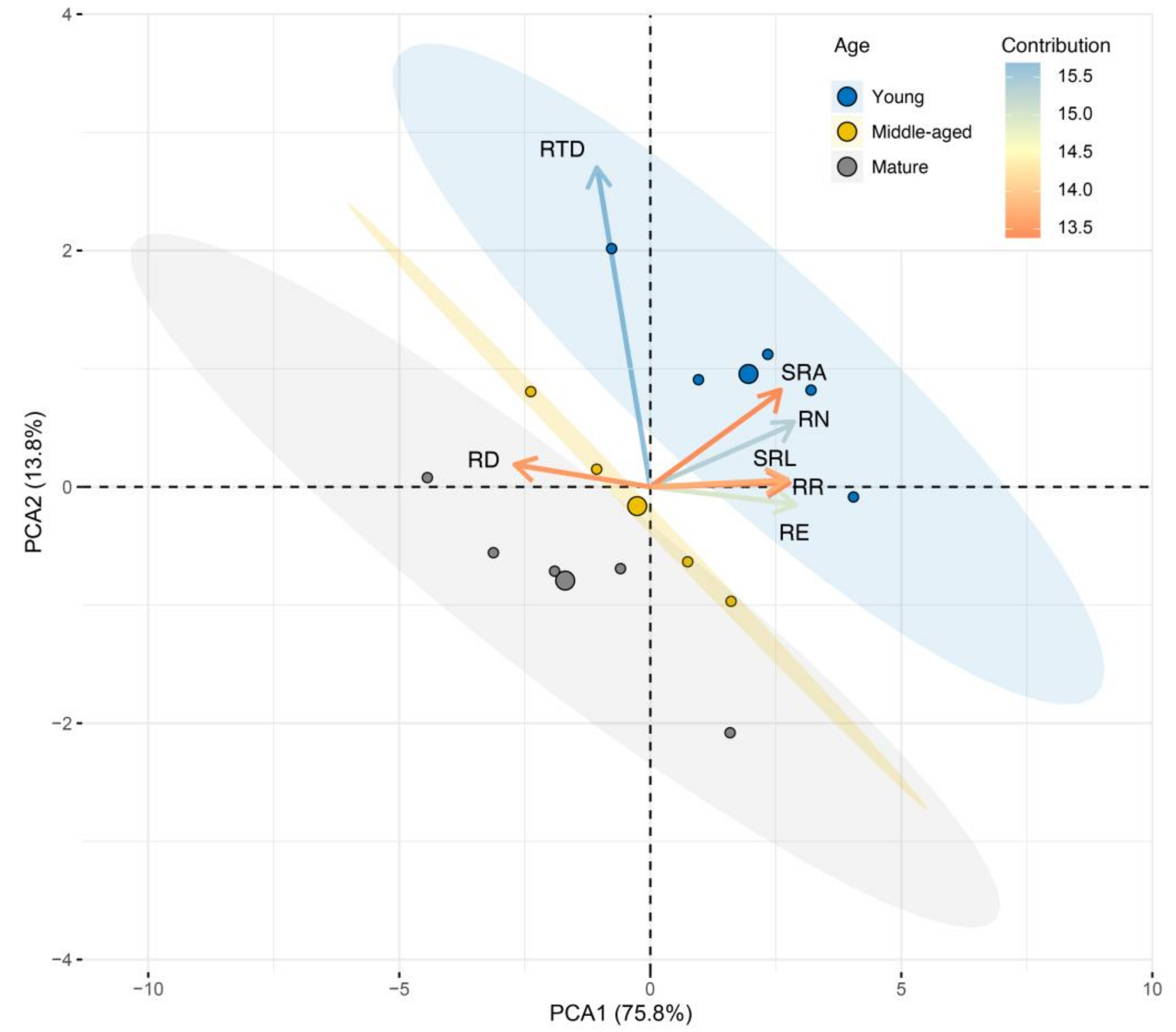
3.3. Linkages of Root Respiration and Exudation Rates to Root Morphological and Chemical Traits
3.4. Relationship between Root Respiration Rate and Exudation Rate
4. Discussion
4.1. Variations of Root Traits Associated with Tree Age
4.2. Relationships of Root Respiration and Exudation with Other Root Traits
4.3. Root-Derived C Allocation to Root Respiration and Exudation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laliberte, E. Below-ground frontiers in trait-based plant ecology. New Phytol. 2017, 213, 1597–1603. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Mommer, L.; De Vries, F.T. Going underground: Root traits as drivers of ecosystem processes. Trends Ecol. Evol. 2014, 29, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Liu, Z.; Wang, Y.; Wang, S.; Ju, C.; Gu, J. Tamm review: Fine root biomass in the organic (O) horizon in forest ecosystems: Global patterns and controlling factors. For. Ecol. Manag. 2021, 491, 119–208. [Google Scholar] [CrossRef]
- Sun, L.; Ataka, M.; Han, M.; Han, Y.; Gan, D.; Xu, T.; Guo, Y.; Zhu, B. Root exudation as a major competitive fine-root functional trait of 18 coexisting species in a subtropical forest. New Phytol. 2021, 229, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Guo, D.; Xu, X.; Lu, M.; Bardgett, R.D.; Eissenstat, D.M.; McCormack, M.L.; Hedin, L. Evolutionary history resolves global organization of root functional traits. Nature 2018, 555, 94–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.; Zhu, B. Linking root respiration to chemistry and morphology across species. Glob. Change Biol. 2021, 27, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Rosenvald, K.; Ostonen, I.; Uri, V.; Varik, M.; Tedersoo, L.; Lõhmus, K. Tree age effect on fine-root and leaf morphology in a silver birch forest chronosequence. Eur. J. For. Res. 2013, 132, 219–230. [Google Scholar] [CrossRef]
- Chen, L.; Deng, Q.; Yuan, Z.; Mu, X.; Kallenbach, R.L. Age-related C:N:P stoichiometry in two plantation forests in the Loess Plateau of China. Ecol. Eng. 2018, 120, 14–22. [Google Scholar] [CrossRef]
- Jagodziński, A.M.; Ziółkowski, J.; Warnkowska, A.; Prais, H. Tree age effects on fine root biomass and morphology over chronosequences of Fagus sylvatica, Quercus robur and Alnus glutinosa stands. PLoS ONE 2016, 11, e0148668. [Google Scholar] [CrossRef] [PubMed]
- Bouma, T.J.; Yanai, R.D.; Elkin, A.D.; Hartmond, U.; Alva, D.E.; Eissenstat, D.M. Estimating age-dependent costs and benefits of roots with contrasting life span: Comparing apples and oranges. New Phytol. 2001, 150, 685–695. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Respiration. In Plant Physiological Ecology, 2nd ed.; Lambers, H., Chapin, F.S., Pons, T.L., Eds.; Springer: New York, NY, USA, 2008; Volume 3, pp. 101–150. [Google Scholar]
- Chai, Y.N.; Schachtman, D.P. Root exudates impact plant performance under abiotic stress. Trends Plant Sci. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.B.; Brzostek, E.R.; Craig, M.E.; Fisher, J.B.; Phillips, R.P. Root-derived inputs are major contributors to soil carbon in temperate forests, but vary by mycorrhizal type. Ecol. Lett. 2021, 24, 626–635. [Google Scholar] [CrossRef]
- Walker, T.S.; Bais, H.P.; Vivanco, G. Root exudation and rhizosphere biology. Plant Physiol. 2003, 132, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.; De Vries, F.T. Plant root exudation under drought: Implications for ecosystem functioning. New Phytol. 2020, 225, 1899–1905. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Ge, Z.; An, G.; Duan, Q.; You, X.; Huang, Y. Soil respiration in poplar plantations in northern China at different forest ages. Plant Soil. 2012, 360, 109–122. [Google Scholar] [CrossRef]
- Li, J.; Jiang, X.; Yin, H.; Yin, C.; Wei, Y.; Liu, Q. Root exudates and soil microbes in three Picea asperata plantations with different stand ages. Chin. J. Appl. Ecol. 2014, 25, 325–332. [Google Scholar]
- Sun, L.; Ataka, M.; Kominami, Y.; Yoshimura, K. Relationship between fine-root exudation and respiration of two Quercus species in a Japanese temperate forest. Tree Physiol. 2017, 37, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Stovall, J.P.; Seiler, J.R.; Fox, T.R. Respiratory C fluxes and root exudation differ in two full-sib clones of Pinus taeda (L.) under contrasting fertilizer regimes in a greenhouse. Plant Soil. 2013, 363, 257–271. [Google Scholar] [CrossRef]
- De Vries, F.T.; Williams, A.; Stringer, F.; Willcocks, R.; McEwing, R.; Langridge, H.; Straathof, A.L. Changes in root-exudate-induced respiration reveal a novel mechanism through which drought affects ecosystem carbon cycling. New Phytol. 2019, 224, 132–145. [Google Scholar] [CrossRef]
- Hill, P.; Kuzyakov, Y.; Jones, D.; Farrar, J. Response of root respiration and root exudation to alterations in root C supply and demand in wheat. Plant Soil. 2007, 291, 131–141. [Google Scholar] [CrossRef]
- Ataka, M.; Sun, L.; Nakaji, T.; Katayama, A.; Hiura, T. Five-year nitrogen addition affects fine root exudation and its correlation with root respiration in a dominant species, Quercus crispula, of a cool temperate forest, Japan. Tree Physiol. 2020, 40, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; McLaughlin, N.B.; Gu, J.; Li, X.; Wang, Z. Relationships between root respiration rate and root morphology, chemistry and anatomy in Larix gmelinii and Fraxinus mandshurica. Tree Physiol. 2013, 33, 579–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Wang, X.; Mao, Z.; Jiang, Z.; Gao, Y.; Chen, X.; Aubrey, D.P. Root exudation rates decrease with increasing latitude in some tree species. Forests 2020, 11, 1045. [Google Scholar] [CrossRef]
- Mori, S.; Hagihara, A. Root respiration in Chamaecyparis obtusa trees. Tree Physiol. 1991, 8, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, X.; Zhou, J.; Wei, H.; Du, Z. Fine root distribution and activity of Fraxinus velutina plantations based on the forest stand age in the Yellow River Delta. J. Arid Land Resour. Environ. 2020, 34, 171–177. [Google Scholar]
- Jin, Y.; Wang, C.; Zhou, Z. Conifers but not angiosperms exhibit vulnerability segmentation between leaves and branches in a temperate forest. Tree Physiol. 2018, 85, 141–147. [Google Scholar] [CrossRef]
- Yin, H.; Wheeler, E.; Phillips, R.P. Root-induced changes in nutrient cycling in forests depend on exudation rates. Soil Biol. Biochem. 2014, 78, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, Z.; Wang, Z.; Gu, J. Functional trait plasticity but not coordination differs in absorptive and transport fine roots in response to soil depth. Forests 2020, 11, 42. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.; Xia, M.; Wei, X.; Chang, W.; Liu, Y.; Wang, Z. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species. New Phytol. 2008, 180, 673–683. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Deforest, L.J.; Burton, J.A.; Allen, F.M.; Ruess, W.R.; Hendrick, L.R. Fine root architecture of nine north American trees. Ecol. Monogr. 2002, 2, 293–309. [Google Scholar] [CrossRef]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef]
- Phillips, R.P.; Erlitz, Y.; Bier, R.; Bernhardt, E.S. New approach for capturing soluble root exudates in forest soils source. Funct Ecol. 2008, 6, 990–999. [Google Scholar] [CrossRef]
- Makita, N.; Hieano, Y.; Dannoura, D.; Kominami, Y.; Mizoguchi, T.; Ishii, H.; Kanazawa, Y. Fine root morphological traits determine variation in root respiration of Quercus serrata. Tree Physiol. 2009, 29, 579–585. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Wang, Y.; Hoch, G.; Wang, Z.; Gu, J. Linkage of root morphology to anatomy with increasing nitrogen availability in six temperate tree species. Plant Soil. 2018, 425, 189–200. [Google Scholar] [CrossRef]
- Gao, G.; Goebel, M.; Wang, Y.; Wang, Z.; Gu, J. Spatial–temporal variations of absorptive fine roots in the organic and soil layers of a Larix gmelinii forest. Trees 2021, 35, 1013–1023. [Google Scholar] [CrossRef]
- Falster, D.S.; Wavid, I.W.; Wright, I.J. User’s guide to smatr: Standardised major axis tests and routines version 2.0, copyright 2006. Biol. Rev. 2006, 81, 259–291. [Google Scholar]
- Børja, I.; De Wit, H.A.; Steffenrem, A.; Majdi, H. Stand age and fine root biomass, distribution and morphology in a Norway spruce chronosequence in southeast Norway. Tree Physiol. 2008, 28, 773–784. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, X.; Wang, H.; Wang, Z.; Gu, J. Root tip morphology, anatomy, chemistry and potential hydraulic conductivity vary with soil depth in three temperate hardwood species. Tree Physiol. 2016, 36, 99–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nan, H.; Liu, Q.; Xiao, Q. Root foraging ability of spruce (Picea asperata) in two growth stages. Chin. J. Appl. Environ. Biol. 2015, 21, 547–554. [Google Scholar]
- Yang, Z.; Song, S.; Li, J.; Li, H.; Luo, Z. A study on root characteristics and nutrients of different aged Larix principis-rupprechtii Mayr. plantations. Acta Bot. Boreali-Occident. Sin. 2014, 34, 1432–1442. [Google Scholar]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Truiillo, M.; Rivas-Ubach, A.; Oravec, M.; Vecerova, K.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; et al. Opposite metabolic responses of shoots and roots to drought. Sci. Rep. 2014, 4, 6829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, D.; Fridley, J.D. Does plant biomass partitioning reflect energetic investments in carbon and nutrient foraging? Funct. Ecol. 2019, 33, 1627. [Google Scholar] [CrossRef]
- Kurosawa, Y.; Mori, M.; Ferrio, J.P.; Yamaji, K.; Koyama, K.; Haruma, T.; Doyama, K. Initial burst of root development with decreasing respiratory carbon cost in Fagus crenata Blume seedlings. Plant Species Biol. 2020, 36, 146–156. [Google Scholar] [CrossRef]
- Freschet, G.T.; Pagès, L.; Iversen, C.; Comas, L.; Rewald, B.; Roumet, C.; Klimešová, J.; Zadworny, M.; Poorter, H.; Postma, J.; et al. A starting guide to root ecology: Strengthening ecological concepts and standardizing root classification, sampling, processing and trait measurements. New Phytol. 2020, 232, 973–1122. [Google Scholar] [CrossRef] [PubMed]
- Meier, I.C.; Finzi, A.C.; Phillips, R.P. Root exudates increase N availability by stimulating microbial turnover of fast-cycling N pools. Soil Biol. Biochem. 2017, 106, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Burton, A.J.; Jarvvey, A.C.; Jarvi, M.P.; Pregitzer, K.S. Chronic N deposition alters root respiration-tissue N relationship in northern hardwood forests. Glob. Change Biol. 2015, 18, 258–266. [Google Scholar] [CrossRef] [Green Version]
- Kramer-Walter, K.R.; Bellingham, P.J.; Millar, T.R.; Smissen, R.D.; Richardson, S.J.; Laughlin, D.C. Root traits are multidimensional: Specific root length is independent from root tissue density and the plant economic spectrum. J. Ecol. 2016, 104, 1299–1310. [Google Scholar] [CrossRef]
- Weemstra, M.; Mommer, L.; Visser, E.; Ruijven, J.; Kuyper, T.W.; Mohren, G.; Sterck, F. Towards a multidimensional root trait framework: A tree root review. New Phytol. 2016, 211, 1159–1169. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, J.; Weigelt, A.; Plas, F.; Laughlin, D.C.; Kuyper, T.W.; Guerrero-Ramirez, N.; Iversen, C.M.; Kattge, J.; McCormack, M.L.; Meier, I.C.; et al. The fungal collaboration gradient dominates the root economics space in plants. Sci. Adv. 2020, 6, eaba3756. [Google Scholar] [CrossRef]
- Lambers, H.; Atkin, O.K.; Millenaar, F.F. Respiratory patterns in roots in relation to their functioning. In Plant Roots: The Hidden Half, 3rd ed.; Waisel, Y., Eshel, A., Kafkaki, U., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 2002; Volume 3, pp. 521–552. [Google Scholar]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Life Cycles: Environmental influences and adaptations. In Plant Physiological Ecology, 2nd ed.; Lambers, H., Chapin, F.S., Pons, T.L., Eds.; Springer: New York, NY, USA, 2008; Volume 11, pp. 375–402. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Ontogenetic Stage (Tree Age (Years)) | Mean DBH/Ground Diameter (Cm) | Mean Height (M) | Soil Ph | Total Soil Carbon (G Kg−1) | Total Soil Nitrogen (G Kg−1) | Soil Total Available Nitrogen (Mg Kg−1) | Soil Available Phosphorus (Mg Kg−1) |
|---|---|---|---|---|---|---|---|
| Young (4–10) | 2.17 ± 0.41 | 0.64 ± 0.08 | 5.55 ± 0.06b | 66.44 ± 6.22b | 4.79 ± 0.43b | 24.54 ± 2.20a | 7.93 ± 1.35a |
| Middle-aged (30–50) | 16.26 ± 1.80 | 17.16 ± 1.61 | 5.51 ± 0.10b | 151.44 ± 14.81a | 12.05 ± 0.99a | 24.48 ± 1.45a | 9.65 ± 1.81a |
| Mature (70–90) | 35.46 ± 0.82 | 24.97 ± 1.69 | 6.08 ± 0.13a | 149.93 ± 21.89a | 12.23 ± 1.69a | 25.94 ± 1.92a | 7.94 ± 1.23a |
| Root Traits | RR (n mol CO2 g−1 h−1) | RE (mg C g−1 h−1) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Young | Middle-Aged | Mature | All Ages | Young | Middle-Aged | Mature | All Ages | ||
| RD (mm) | r | −0.89 | −0.94 | −0.85 | −0.93 | −0.74 | −0.90 | −0.91 | −0.84 |
| p | 0.046 | 0.016 | 0.067 | <0.001 | 0.156 | 0.039 | 0.035 | <0.001 | |
| SRL (m g−1) | r | 0.95 | 0.97 | 0.71 | 0.73 | 0.82 | 0.96 | 0.76 | 0.84 |
| p | 0.012 | 0.005 | 0.177 | 0.002 | 0.088 | 0.011 | 0.137 | <0.001 | |
| SRA (m2 kg−1) | r | 0.96 | 0.97 | 0.99 | 0.72 | 0.97 | 0.94 | 0.97 | 0.87 |
| p | 0.010 | 0.005 | 0.002 | 0.001 | 0.005 | 0.018 | 0.007 | 0.011 | |
| RTD (g cm−3) | r | −0.98 | −0.97 | −0.82 | −0.28 | −0.93 | −0.97 | −0.90 | −0.40 |
| p | 0.004 | 0.007 | 0.088 | 0.309 | 0.024 | 0.008 | 0.039 | 0.135 | |
| RN (mg g−1) | r | 0.98 | 0.76 | 0.94 | 0.89 | 0.89 | 0.84 | 0.97 | 0.91 |
| p | 0.004 | 0.133 | 0.019 | <0.001 | 0.044 | 0.075 | 0.008 | <0.001 | |
| Ontogenetic Stage (Tree Age (Years)) | Slope | Intercept | R2 | p |
|---|---|---|---|---|
| Young (4–10) | 0.55a | −0.34 | 0.84 | 0.028 |
| Middle-aged (30–50) | 0.19b | 0.15 | 0.94 | 0.007 |
| Mature (70–90) | 0.27ab | 0.09 | 0.95 | 0.005 |
| All ages | 0.26 | 0.11 | 0.82 | 0.022 |
| Fine Root Traits | Total Soil Carbon (g kg−1) | Total Soil Nitrogen (g kg−1) | Soil pH | |||
|---|---|---|---|---|---|---|
| Correlation Coefficient (r) | p | Correlation Coefficient (r) | p | Correlation Coefficient (r) | p | |
| RR (n mol CO2 g−1 h−1) | −0.011 | 0.969 | −0.077 | 0.785 | −0.062 | 0.826 |
| RE (mg C g−1 h−1) | −0.088 | 0.755 | −0.138 | 0.624 | 0.060 | 0.829 |
| RD (mm) | −0.107 | 0.704 | −0.044 | 0.875 | 0.036 | 0.899 |
| SRL (m g−1) | −0.127 | 0.653 | −0.152 | 0.588 | 0.067 | 0.814 |
| SRA (m2 kg−1) | −0.473 | 0.075 | −0.500 | 0.058 | −0.105 | 0.709 |
| RTD (g cm−3) | −0.757 | 0.001 | −0.739 | 0.002 | −0.743 | 0.002 |
| RN (mg g−1) | −0.227 | 0.416 | −0.282 | 0.309 | −0.081 | 0.774 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Liu, Z.; Gao, G.; Yang, X.; Gu, J. Shift from Acquisitive to Conservative Root Resource Acquisition Strategy Associated with Increasing Tree Age: A Case Study of Fraxinus mandshurica. Forests 2021, 12, 1797. https://doi.org/10.3390/f12121797
Li Z, Liu Z, Gao G, Yang X, Gu J. Shift from Acquisitive to Conservative Root Resource Acquisition Strategy Associated with Increasing Tree Age: A Case Study of Fraxinus mandshurica. Forests. 2021; 12(12):1797. https://doi.org/10.3390/f12121797
Chicago/Turabian StyleLi, Zuwang, Zhi Liu, Guoqiang Gao, Xinlei Yang, and Jiacun Gu. 2021. "Shift from Acquisitive to Conservative Root Resource Acquisition Strategy Associated with Increasing Tree Age: A Case Study of Fraxinus mandshurica" Forests 12, no. 12: 1797. https://doi.org/10.3390/f12121797
APA StyleLi, Z., Liu, Z., Gao, G., Yang, X., & Gu, J. (2021). Shift from Acquisitive to Conservative Root Resource Acquisition Strategy Associated with Increasing Tree Age: A Case Study of Fraxinus mandshurica. Forests, 12(12), 1797. https://doi.org/10.3390/f12121797