1. Introduction
As a fast-growing bamboo species with ecological and economic benefits,
Phyllostachys edulis (Carrière) J. Houz plays an important role in replacing wood with bamboo and ecological construction. According to the Intergovernmental Panel on Climate Change (IPCC), with the aggravation of global extreme climate that has been linked to the increased frequency and intensity of drought stress events, the high-risk level confidence will increase [
1]. Drought caused by seasonal factors in bamboo areas of China was considered to be one of the difficult problems affecting the cultivation of bamboo forests and the preservation of germplasm resources. In addition, water is also an important ecological factor that restricts the geographical distribution and growth of bamboo. Therefore, to explore the physiological response mechanism of bamboo under drought stress was an important breakthrough for the study of drought resistance adaptability and water-saving and the efficient cultivation of bamboo, and also a strong guarantee for improving the conservation strategy of bamboo germplasm resources.
Yuan et al. found that water potential was the key factor driving the rapid growth of
P. edulis during the growth dynamic process from bamboo shoots to young bamboo [
2]. The related physiological research of
P. edulis in response to drought stress mainly focused on single physiological indexes such as Photosynthetic Fluorescence, Chlorophyll Content, Electrical Conductivity or Antioxidant Enzyme Activity [
3,
4,
5,
6]. Li’s research results showed that the activities of antioxidant enzymes such as superoxide dismutase (SOD) and peroxidase (POD) in bamboo leaves were significantly increased under drought stress to remove reactive oxygen species and alleviate lipid peroxidation of the cell membrane. Furthermore, the contents of soluble sugars such as sucrose and fructose in
P. edulis leaves were increased to maintain cell turgor pressure [
5]. In rice in the same family of Gramineae as bamboo, Ca
2+-dependent protein kinase plays an important role in regulating its drought tolerance response [
7]. The endogenous Ca
2+ cascade of H
2O
2 and NO signaling molecules improves the activity of C
4 phosphoenolpyruvate carboxylase (PEPC) by affecting the transcription level and phosphorylation level, which respond quickly to the external environment to resist stress [
8,
9]. Ying et al. used polyethylene glycol (PEG) with different concentrations to replace drought stress patterns. The results showed that the Ca
2+ fluorescence intensity in the pileorhiza and elongation region is stronger, and the Ca
2+ accumulation in the root tip cytoplasm is more obvious with the intensity of drought stress [
10]. However, the physiological response mechanism of
P. edulis to drought stress based on Ca
2+ signaling has not been systematically reported.
To fill this research gap, in this study we used polyethylene glycol-6000 (PEG-6000) to replace drought stress treatment to monitor the Ca2+ flux and distribution response characteristics in the root tip of P. edulis seedlings. The physiological indicators of samples of leaves with 13 gradients, which were selected from the continuous multi-level soil water content by a manual control experiment, were detected. The main physiological information of the top three principal components, which were selected by PCA and correlation analysis (calculated based on the contribution rate of cumulative variance above 85%), from all the data was analyzed. The combination of Ca2+ flux and relevant physiological indicators was analyzed to reveal the ecological adaptation strategy of P. edulis to drought stress, and provide water-related physiological data support for germplasm resource conservation and efficient cultivation.
2. Materials and Methods
2.1. Plant Materials and Treatment
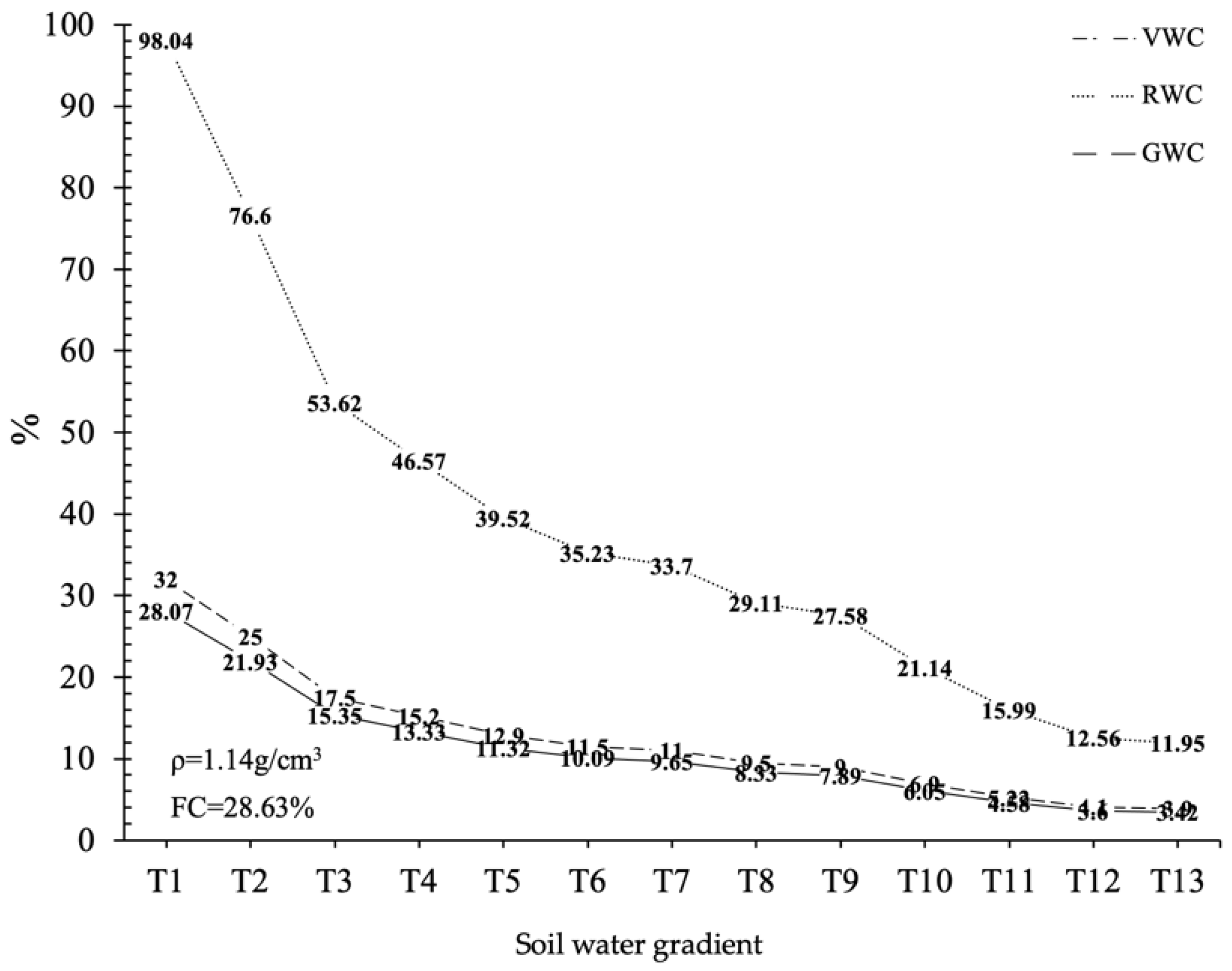
The experimental materials were three-year-old P. edulis seedlings (collected from the same parent bamboo in Guilin, Guangxi, China). The experiment was set in the International Centre for Bamboo and Rattan, Anhui Taiping Experimental Center (30.4° N, 118.2° E). The experimental area was set in the subtropical climate zone. The average temperature is above 0 °C in January and about 26 °C in July, and about 15 °C throughout the year. The annual precipitation is about 800–1600mm. The planting container of P. edulis seedlings was a cylindrical plant pot (diameter × height, 26 cm × 30 cm). The soil bulk density (ρ) and the maximum field water holding capacity (FC) in the planting pot were determined by the ring knife method and the drying method. The basin was filled with soil (ρ = 1.14 g/cm3. FC = 28.63%), which was compacted.
Healthy and growth-uniform seedlings were selected, and then soaked with the plant pot until the soil water was saturated. Test seedlings were placed on the flat ground under independent space made of waterproof cloth to avoid the interference of external water factors such as soil water storage and rainwater. During the experiment,
P. edulis seedlings were not watered when the soil was saturated with water. The soil water content of
P. edulis seedlings in the planting pot was reduced by transpiration until the drought stress of the
P. edulis seedlings, which gradually wilted and died. Soil volumetric water content (VWC) was recorded by Em-50 (Decagon, Pullman, WA, USA) soil temperature and humidity monitoring system, and 13 soil VWC gradients were selected, as shown in
Figure 1. The three attribute values of 13 soil water contents, which were converted from VWC, GWC and RWC, were labeled, respectively, as T1, T2, T3, T4, T5, T6, T7, T8, T9, T10, T11, T12 and T13.
The experiment used continuous monitoring and regular sampling method to collect healthy leaves (about 10 g of fresh sample) from the middle and upper parts of branches of the P. edulis seedlings under 13 soil water gradients, which were quick-frozen and preserved. Five replicates (each replicate was one bamboo sample) were used for physiological index determination in each treatment.
In addition,
P. edulis seedling roots were treated by drought stress simulated by 20% PEG-6000 (Coolaber, Beijing, China) [
10]. The real-time changes of net Ca
2+ flux were measured by non-invasive micro-test technology (Physiolyzer, Younger USA LLC, Amherst, MA, USA; Xuyue (Beijing) Sci.&Tech. Co., Ltd., Beijing, China).
2.2. Determination of Antioxidant Enzyme Activity in P. edulis Seedling Leaves
In the experiments, healthy leaf (vein removal) samples were obtained from the branches of middle and upper parts of P. edulis seedlings and placed in a pre-cooling agate mortar, and were ground with 2 mL of pre-cooled phosphate buffer (PBS, 50 mmol·L−1) and a small amount of quartz sand. The mixture was then centrifuged for 15 min at 10,000 rpm and supernatant solution was decanted.
The supernatant solution was used for further studies, and superoxide dismutase (SOD) activity was detected by the NBT method. POD activity was detected by the guaiacol colorimetric method. Catalase (CAT) activity was detected by the UV absorption method.
2.3. Determination of Substance Content in P. edulis Seedling Leaves by Enzyme-Linked Immunosorbent Assay
Ribulose-1,5-diphosphate (RuBP), calcium receptor proteins (Calmodulin, CaM; Calcium-dependent protein kinases, CDPKs), Malondialdehyde (MDA), osmoregulatory substances (Proline, Pro; Betaine, Bet) and hormones (Abscisic acid, ABA; Indole-3-acetic acid, IAA and Jasmonic acid, JA) of P. edulis seedling leaves were determined by enzyme-linked immunosorbent assay (MEIMIAN Industrial Co., Ltd., Yancheng, Jiangsu, China) to affirm their contents.
The leaf samples were ground in PBS and the extract was obtained for testing, with 5 biological replicates per treatment. The absorbance (OD) was measured at 450 nm by a microplate reader (Finland, Labsystems Multiskan MS, 352 Model), and the content of each substance was calculated by drawing a standard curve.
2.4. Real-Time Determination of Ca2+ Flux in P. edulis Seedlings
The real-time regular of net flux of Ca2+ in P. edulis seedlings was measured by non-invasive micro-test technology (Physiolyzer, Younger USA LLC, Amherst, MA, USA; Xuyue (Beijing) Sci.&Tech. Co., Ltd., Beijing, China). The root samples of Phyllostachys edulis seedlings were carefully rinsed with distilled water and fixed at the bottom of the Petri dish. Test solution (0.1 CaCl2, 0.5 KCl, pH 6.0) was added to the Petri dish at room temperature, and discarded after standing for 10min. Then, 5 mL of fresh test solution was added. The tip of the Ca2+ flux microsensor was positioned to the test area of the root about 600 μm from the root apex, and valid data were recorded for 3–5 min. Subsequently, the test fluid in the Petri dish was replaced with the test fluid with 20% PEG-6000, which can simulate drought stress, and Ca2+ flux data were recorded by imFluxes v2.0 software (Xuyue (Beijing) Sci.&Tech. Co., Ltd., Beijing, China). The experiment was repeated with 6 P. edulis seedlings per treatment. The leaf samples were incubated in the measuring solution to equilibrate for 30 min and then transferred to a measuring chamber containing 10 mL of a fresh measuring solution. The method for the measurement of Ca2+ flux in leaf samples was the same as that for roots.
2.5. Observation of P. edulis Seedlings by Laser Confocal Microscopy Luminescence Imaging
The calcium ion fluorescent probe Fluo-8 TM (AAT Bioquest, Sunnyvale, CA, USA) was fully dissolved in anhydrous dimethyl sulfoxide DMSO (AAT Bioquest, Sunnyvale, CA, USA) to a concentration of 1 mmol/L. The P. edulis roots and the lower epidermis of the leaves were incubated in Hank’s balanced salt solution (without calcium ions) containing 20 µmol/L Fluo-8TM, AM for 40 min at room temperature in the dark, and then the lower epidermis of P. edulis leaves was rinsed several times with the buffer and incubated for another 20 min at room temperature (to ensure sufficient dissolution of the esterification probe), which was finally placed on a slide and 0.5 mL of Hank’s buffer was added dropwise to prepare the slide. The root tip sampling should be performed by cutting the root tips of intact, healthy and white-colored P. edulis hydroponics seedlings and made into slides, which was labeled by the above fluorescent method. All parameters of the instrument were kept consistent during the test (excitation wavelength 488 nm, BP 505-530, Pinhole 280, DG 581, AO 0.1, AG 1.34). At least three fields of view in different replicates were selected for each experiment for observational analysis. Vascular bundle characteristics of P. edulis stem sections were observed using laser confocal microscopy (LSM510, LeicaDM4, Berlin, Germany) at 488 nm excitation wavelength based on lignin autofluorescence properties.
2.6. Statistical Analysis
Data were analyzed by Excle 2020, SPSS 18.0 (SPSS Inc., Chicago, IL, USA) and RStudio software. The physiological indexes of P. edulis seedlings under different soil moisture gradients were standardized to form 12 principal components. FactoMineR and other program packages were used for principal component analysis, and the eigenvectors, eigenvalues and cumulative contribution rates of the sample correlation matrix were calculated. A regression equation model was established between soil relative water content (RWC) and principal component factors with large contribution rates by SPSS 18.0 (SPSS Inc., Chicago, IL, USA).
3. Results
3.1. PCA Analysis of Physiological Characteristics of P. edulis Seedlings under Different Soil Water Treatments
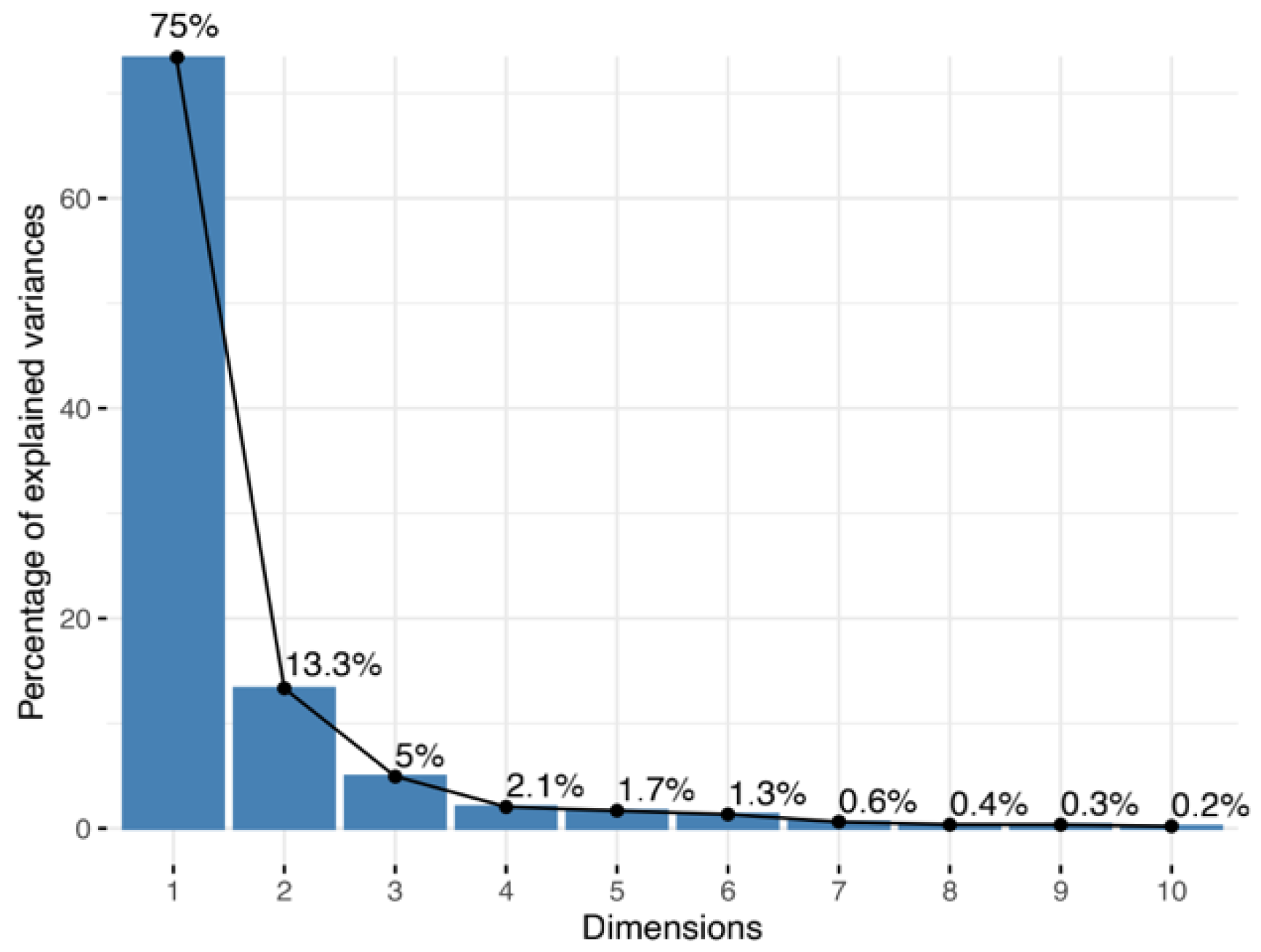
Twelve relevant physiological indicators of
P. edulis seedlings under 13 soil water gradient treatments were transformed into different principal components. The contribution rates of the variance of the first three principal components of the
P. edulis seedlings related to soil water content were 75.0%, 13.3% and 5.0%, respectively, with a cumulative contribution rate of 93.3%, which represented 93.3% of the comprehensive information (moisture physiology) of the related indexes (
Figure 2). Therefore, only the first three principal components that can comprehensively reflect the main information of the raw data of each test index were selected as effective principal components for the analysis.
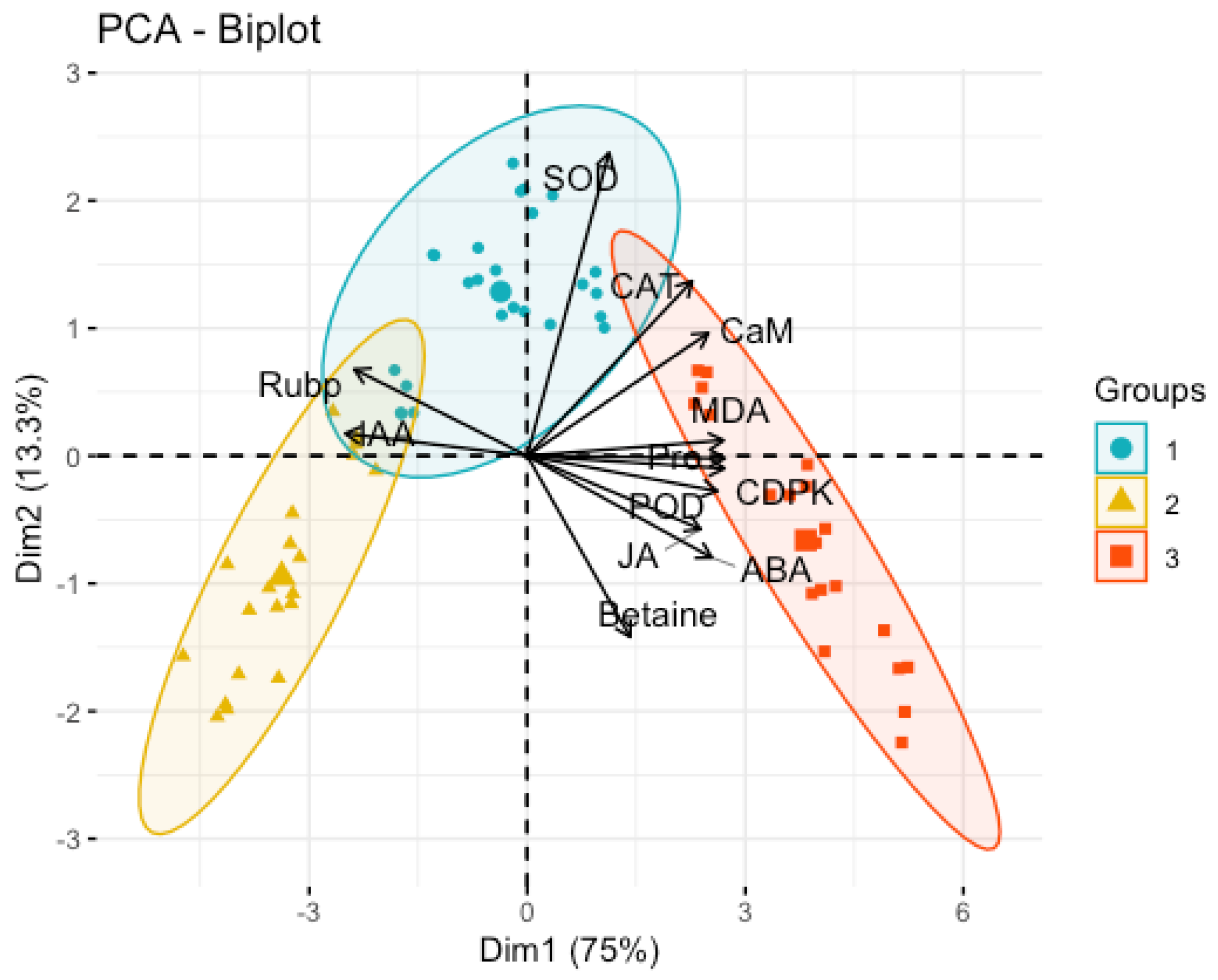
The factor loading contributions of the first and second principal components were graphically visualized, as shown in
Figure 3. The analysis revealed that the factors contributing more to their loadings in the first principal component were Pro, CaM, CDPKs, MDA, IAA, JA, and ABA, and in the meantime, SOD factor loading was large in the second principal component.
In order to accurately describe the factor loading contribution of each principal component, this study further quantified the physiological factor loadings (contribution) for the first three principal components related to the soil water content of
P. edulis, as shown in
Table 1. Among them, the matrix eigenvalue of the first principal component was 3.0002, and the variance contribution rate was 75.0%. As the most important sample matrix among all the variables, they are X11 (cytoplasmic osmoregulatory substance Pro), X10 (calcium signaling protein kinases CDPKs), X8 (membrane oxidation degree MDA), X3 (antioxidant enzyme POD), X4 (endogenous hormone ABA), X6 (endogenous hormone IAA), X9 (CaM) and X5 (endogenous hormone JA) in order of the magnitude of the contribution of the factor loadings (X > 2.9), which are the key factors of
P. edulis in response to drought stress in the first principal component. They can be attributed to the mechanisms of calcium signaling-based regulation of
P. edulis plasma membrane tolerance in response to drought stress and endogenous hormone synergistic growth rate regulation by analyzing factor attributes. The aim is to reflect the main physiological coping strategies of
P. edulis in response to drought stress (based on calcium signals).
The eigenvalue of the second principal component was 1.2647 with a variance contribution of 13.3%. The highest ranking of the loading contribution rates according to the absolute value of eigenvector coefficients resulted in: X1 (antioxidant enzyme activity SOD). Data analysis showed that antioxidant enzyme activity was the most critical factor in the second principal component, which can be increased to scavenge reactive oxygen species to resist drought stress. That is, the second principal component mainly reflects the ability of Phyllostachys edulis to inhibit the oxidation of the organism and resist drought stress.
The eigenvalue of the third principal component was 0.7714 with a variance contribution of only 5.0%, and the factor loading (contribution rate) was mainly from X12 (Betaine), which is an important cytoplasmic osmoregulatory substance to protect osmotic pressure and enhance plants’ resistance to drought. That is to say, the third principal component is involved in regulating osmotic pressure in P. edulis to ease environmental stress from drought.
Then, the study analyzed the correlation of physiological indicators in the leaves associated with water stress of
P. edulis. The results showed that there were significant correlations among calcium sensing protein CaM and CDPK content, osmotic adjustment substances, plant endogenous hormone content and the content of cell membrane lipid oxidation product MDA (
Table 2). However, it was hypothesized that the regulation of osmotic stress on drought resistance and the co-regulation of hormones were both caused by calcium signal transduction.
3.2. The Reply of Calcium Signaling in the Physiological Role of P. edulis in Response to Drought Stress
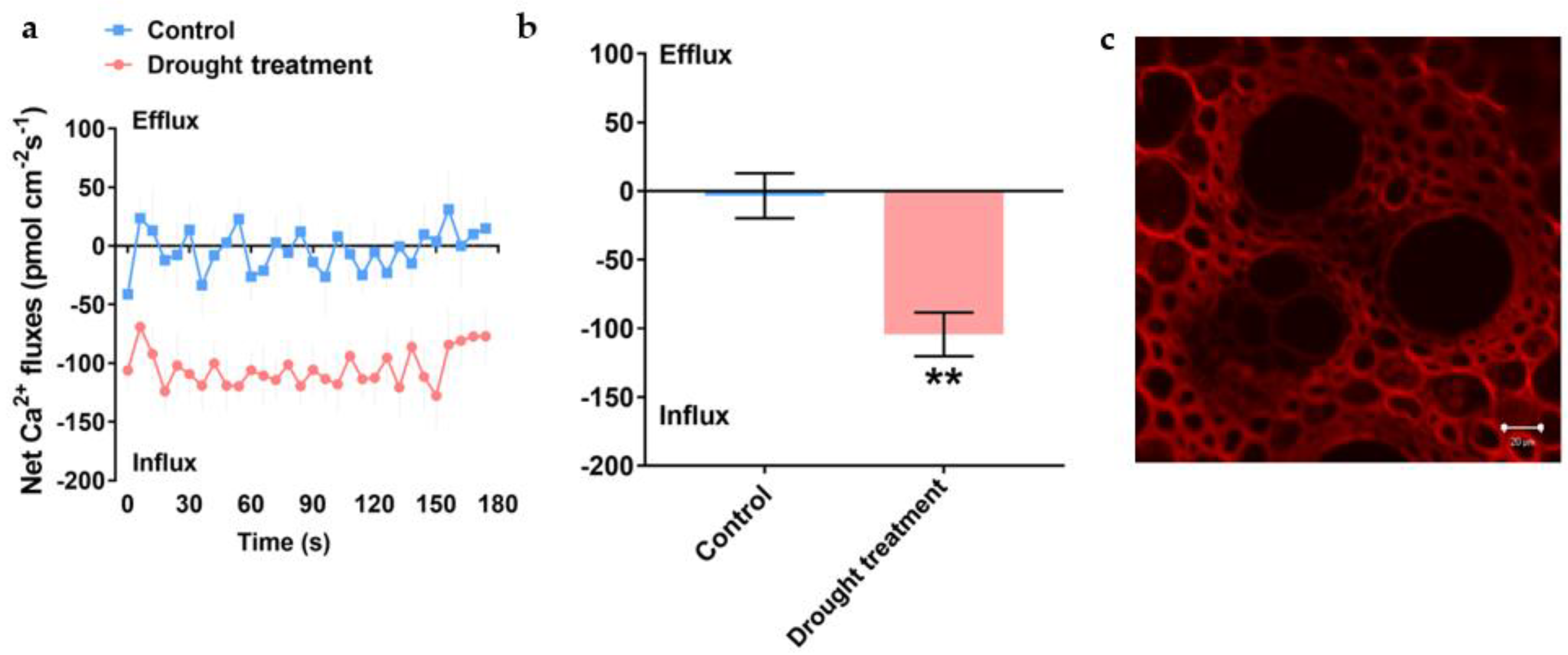
The plant–root system is the direct function organ of the plant to sense the surrounding soil water, which plays a major role in nutrient uptake. Drought stress induces root chemical signals, such as abscisic acid (ABA), growth hormone (IAA) and Ca2+, which are transmitted to the above-ground part of the plant through long-distance transport to reduce stomatal conductance, inhibit transpiration and enhance water use efficiency. To verify the correlation between calcium-sensing protein content and its cellular free Ca2+ concentration in P. edulis under drought stress, we examined the real-time Ca2+ flux in response to drought stress in the root tip and leaf parts of P. edulis.
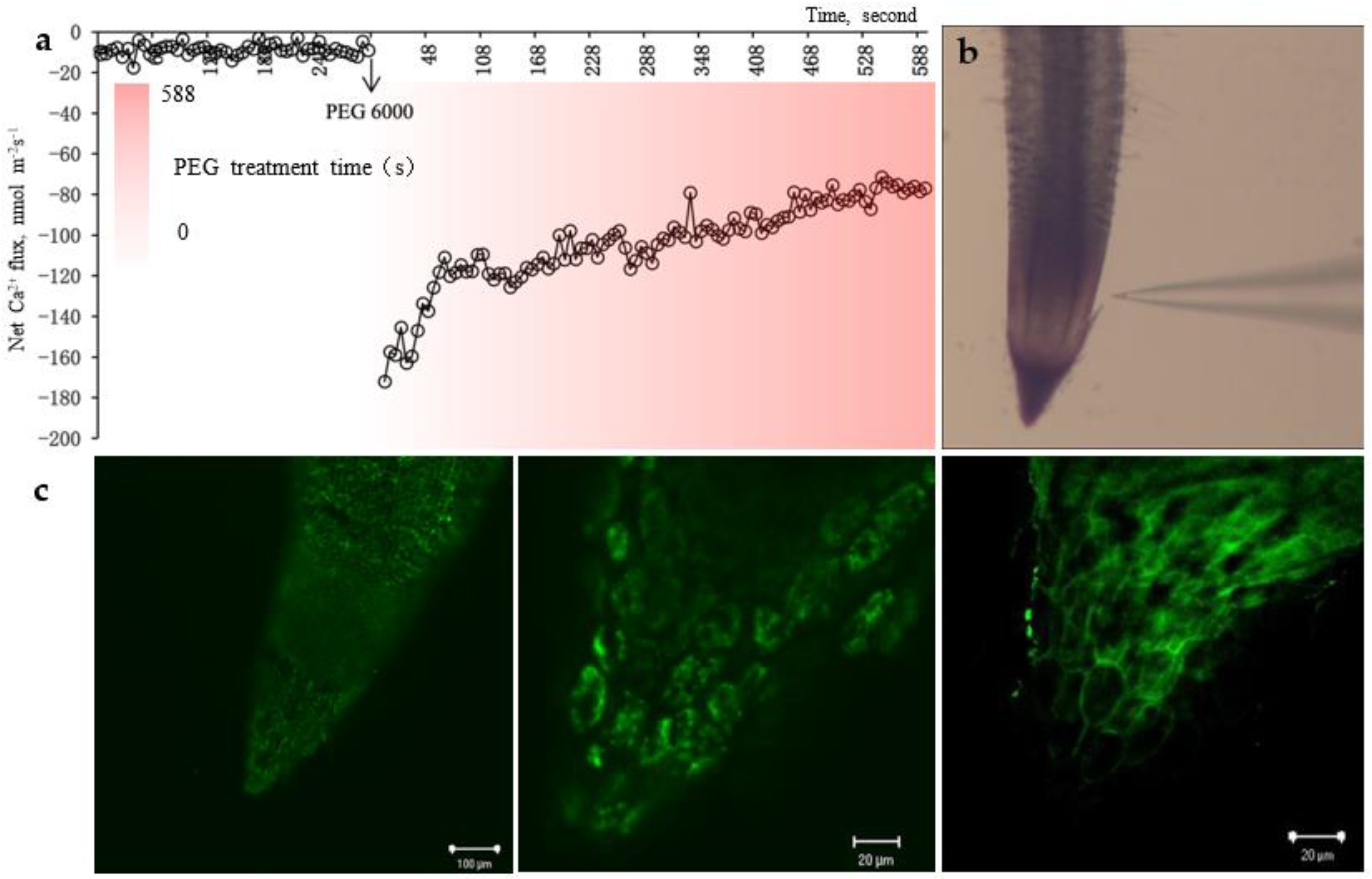
When
P. edulis root tips were treated with 20% PEG-6000 simulated drought stress, the Ca
2+ flux on the root tip surface (monitored at 600 μm from the tip) shifted into a stable weak endocytosis to a strong endocytosis, followed by a gradual weakening of Ca
2+ uptake capacity after reaching a peak, which may be related to the integrated capacity and status of Ca
2+ and its receptors CaM and CDPKs at different time periods during the Ca
2+ signal transduction process, which represents the performance characteristics of root Ca
2+ flux of
P. edulis in response to drought stress (
Figure 4a,b). Ca
2+ in the root tip cell wall of
P. edulis seedlings was treated with drought stress by 20% PEG-6000 diffused into the cytoplasm (
Figure 4c). Further analysis in combination with
Figure 4a showed that the increase in free Ca
2+ concentration in cytoplasmic
P. edulis root cells due to drought stress may be caused by the release of cytosolic calcium pools and the extracellular entry of Ca
2+.
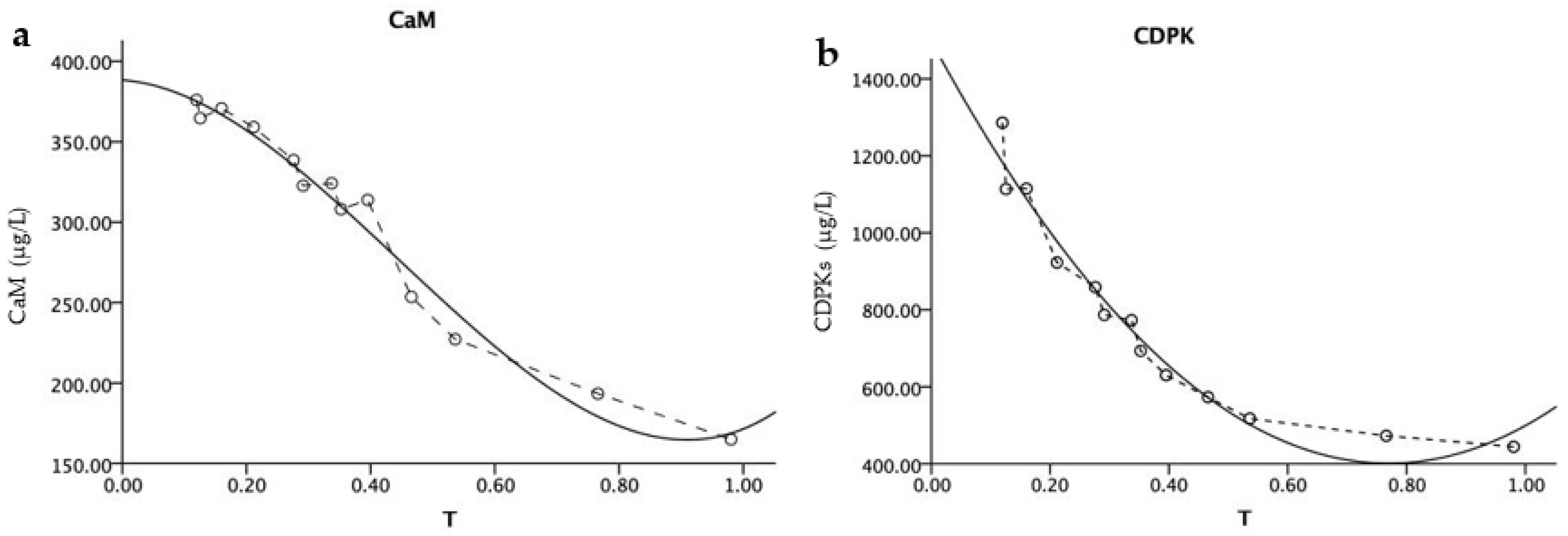
According to the regression model of calcium sensing protein (CaM, CDPKs) contents and relative soil water content (RWC) (
Table 3), the analysis showed that with the decrease in relative soil water content (RWC), the content of CaM and CDPKs in
P. edulis leaves showed a “U” pattern of falling then rising. The water stress affected the contents of CaM and CDPKs, as the receptor of Ca
2+ signaling in drought signal transduction. The results showed that water deficit or excess will cause the calcium sensor protein CaM and CDPK content to increase (
Figure 5).
The stomatal closure of leaves is triggered when plant roots perceive drought stress. It is commonly believed that plant stomatal closure caused by the increase in water potential of guard cells is one of the response events to drought stress. The current theories of stomatal movement include the starch–sugar conversion theory, inorganic ion uptake theory and malate production theory. However, the prevailing theory is that the stomata close because Ca2+ enters the cytosol, depolarizing the membrane and opening the anion channels, releasing chloride ions and malic acid for discharge.
Using fluorescent staining/lignin autofluorescence properties, it was found that drought stress caused stomata composed of dumbbell-shaped guard cells of
P. edulis leaves to be in a closed state and Ca
2+ concentration in stomatal cells increased (
Figure 6a,b). In this study, Ca
2+ flux of uptake was found to be significantly enhanced in its leaf sections by using the NMT technique, which verified the association between the closure of guard cells and intracellular Ca
2+ concentration. This result suggests that the elevated Ca
2+ concentration in guard cells may be related to the cell wall calcium store and the influx of extracellular free Ca
2+, which is considered to be the regulatory mechanism of guard cells in response to drought stress.
The above studies indicated that Ca2+ as the second messenger in response to drought stress plays a role as the signal base of physiological response. Ca2+ is considered to be closely related to endogenous hormone regulation and osmotic regulation. The study’s physiological indicators of coping with drought stress were divided into three categories based on the PCA of Ca2+ signal-related physiological responses of P. edulis. The three categories are endogenous hormone regulation, osmoregulation and antioxidant enzyme activity regulation. In order to further clarify the response pattern of the above main water-related physiological indicators with soil water change, a regression model was further constructed between endogenous hormone regulation (ABA, IAA and JA), osmoregulation indicators and soil water content.
Meanwhile, the root chemical signals of
P. edulis are mainly transmitted to the above-ground parts through the vascular tissues of its stalks. Using lignin autofluorescence to observe the structure of vascular bundles of
P. edulis stalks, as shown in
Figure 6c, it was found that its vascular bundles consisted of air space, vessel element and sieve-tube member, which laid the structural foundation for the long-distance transport of water and chemical signals such as Ca
2+ and root-derived ABA from roots to leaves. The chemical signals can be transported across long distances through the xylem, contributing to reducing transpiration and inhibiting the growth rate of leaves.
3.3. Analysis of Regression Model between Endogenous Hormone Content of P. edulis Leaves and RWC
Previous studies have found that ABA is essential for physiological processes during drought stress responses [
11]. As a signaling substance, ABA controls stomatal movement to control plant transpiration. It was found that when leaves were dehydrated, ABA concentration in leaf tissues increased, followed by stomatal closure. At the same time, studies have shown that stomatal closure occurs when meager ABA concentration is applied to leaves [
12]. ABA can be transported from roots to aboveground parts through xylem transpiration flow, rapidly reaching leaves and accumulating in the lower epidermis where stomata are densely packed and distributed in guard cells and the outer epidermal cytoplasm [
13]. Intracellular Ca
2+ concentration plays an important role in the ABA signaling pathway for plants in response to drought stress. Ca
2+ and anion channels are possible pathways for ABA-induced stomatal closure, and ABA activates selective ion channels in the plasma membrane and Ca
2+/H
+ exchange system in the vesicles, causing extracellular and intra-vesicular Ca
2+ to flow to the cytoplasm, thus increasing intracellular Ca
2+ concentration.
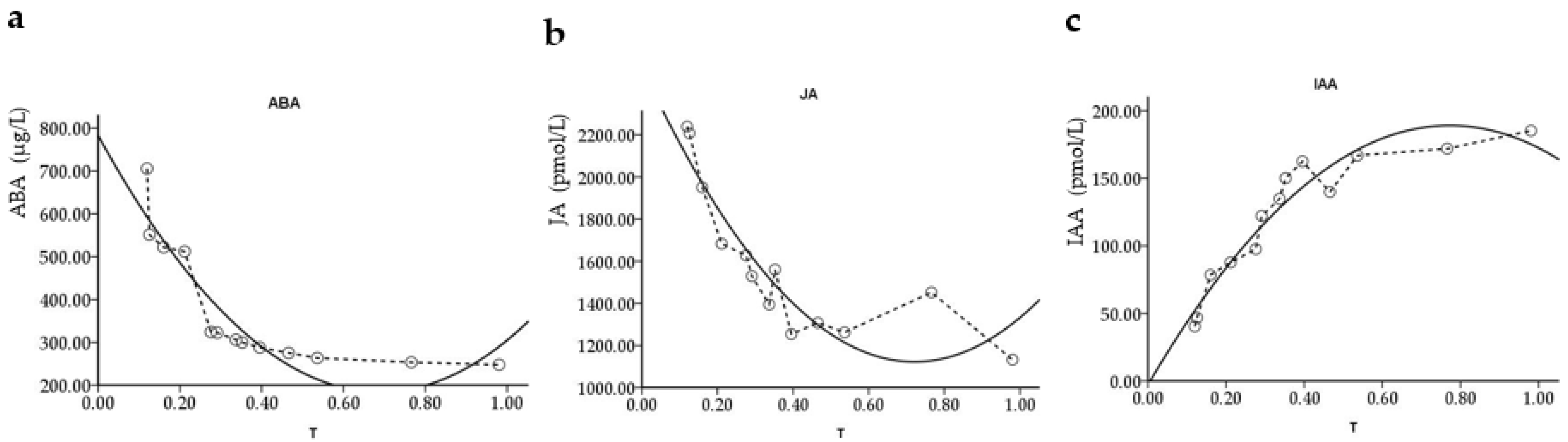
The curvilinear regression of ABA content in
P. edulis leaves with soil RWC is shown in
Figure 7a. The analysis revealed that ABA content was closely related to soil water content around
P. edulis roots. With the decrease in soil RWC from saturated water content, ABA content in leaves firstly decreased and then increased, and the minimum value appeared at RWC = 68.98%. The results showed that with soil water deficiency, and root-derived ABA was transported to leaves from bottom to top with xylem vessel elements, which was the key signal factor leading to its stomatal closure.
IAA as a signaling response substance under drought stress showed a mutually antagonistic relationship with ABA in the process of stomatal closure. It was shown that the highest value of growth hormone content was 194.78 pmol/L in
P. edulis leaves, which occurred at RWC = 77.25%. IAA could regulate stomatal conductance through signaling transport to control
P. edulis growth and transpiration (
Figure 7b).
JA and ABA, meanwhile, interacted under drought stress [
14]. For example, JA can reduce drought-induced damage to the plasma membrane of barley cells [
15] and promote stomatal closure by regulating the defense cell potassium ion channel concentration effect. The analysis of
Figure 7c shows that with the change in soil water content, the trend of JA in the leaves of
P. edulis is opposite to the trend of IAA and the same as the trend of ABA. Combined with the regression model in
Table 4, it is calculated that the minimum value of JA content is 1122.69 pmol/L in the leaves of live
P. edulis seedlings, which stand at the soil RWC of 72.10%.
The results indicate that P. edulis adapts to different soil water contents by physiological mechanisms such as the co-regulation of three hormones, IAA, JA and ABA. Based on calcium signal transduction, the inhibition of plant growth and the regulation of stomatal closure by regulating hormone content help the plant to cope with drought stress.
3.4. Analysis of Regression Model between Osmoregulatory Substances, Malondialdehyde Content of P. edulis Leaves and RWC
Osmotic adjustment (OA) is an important physiological process in P. edulis resistance to drought stress. Ca2+ plays a role in osmotic adjustment (OA), an important physiological process for plants to withstand drought adversity, by stabilizing the balance of anions and cations. Additionally, ABA can induce the synthesis of osmoregulatory substances such as Pro to improve the osmoregulatory ability of cells.
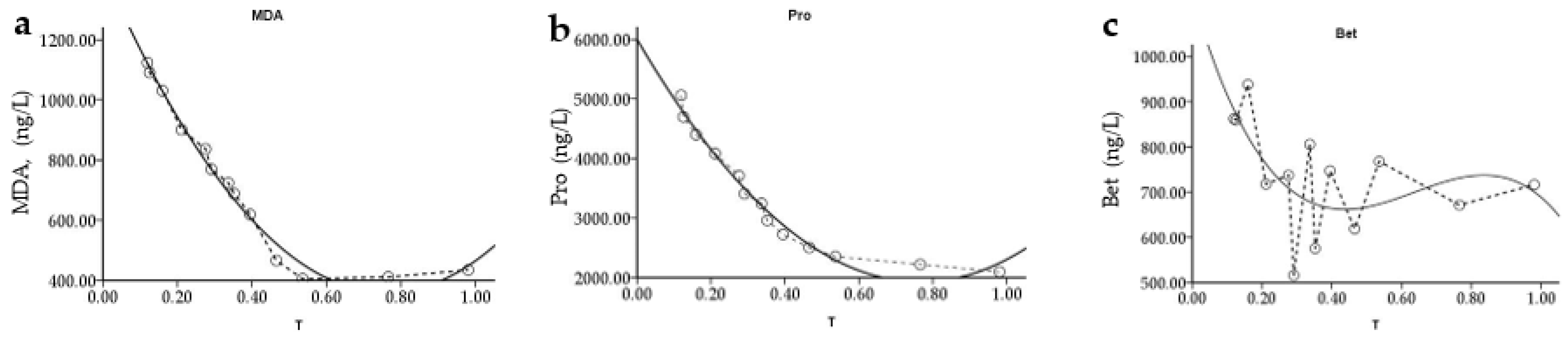
As shown in
Figure 8a, with the decrease in soil water content, MDA content in
P. edulis leaves showed a trend of first decreasing and then increasing. These results indicate that water stress caused by flood and drought soil water content could aggravate the lipid peroxidation of the leaf membrane and increase the content of MDA in the leaf. Combined with the regression model calculation of MDA and RWC in
Table 5, we found that the minimum value of MDA content (RWC
MDAmin) was 76.16%.
With the decrease in RWC, the regression curve between RWC and Pro of
P. edulis leaves showed a U-shaped trend of falling and then rising, while the regression curve between RWC and Bet showed an S-shaped trend of falling–lifting–falling (
Figure 8b,c). Under drought stress,
P. edulis leaves maintain cell expansion pressure by accumulating osmoregulatory substances to resist leaf wilting caused by drought stress.
4. Discussion
Calcium ions are an important second message in the plant response to stress signal transduction [
16]. When plants are stimulated by the environment, the increased Ca
2+ concentration in the cells can trigger the signal to cascade downstream through Ca
2+/CAM, Ca
2+/CDPKs and other pathways, which in turn triggers the expression of related genes and produces a series of physiological responses to resist adversity damage [
17,
18,
19]. Preliminary studies showed that environmental stress signals can activate calcium channels on the plasma membrane of the cell and generate specific calcium signals in the cytoplasm. Then, the activated Ca
2+ signals in the cell are transmitted to calcium signal receptor proteins such as CaM and CDPK. Then, adversity signals, which are cascaded downstream through Ca
2+/CAM and the Ca
2+/CDPKs pathway, trigger the expression of related genes, and a series of physiological responses is produced by plants to resist adversity damage [
20,
21]. In this study, calcium signal-sensing protein kinases (CDPKs), the calcium signal transduction receptor of
P. edulis in response to drought stress, are the most critical factor in the first principal component of
P. edulis in response to drought stress, which comprehensively reflects the calcium signal transduction process of
P. edulis. Further, using PEG-6000 to simulate the drought stress test, the study shows that the root tip Ca
2+ of
P. edulis that has been treated with PEG-6000 shows enhanced influx, which implied that the Ca
2+ signal-related pathways are involved in the response of
P. edulis to drought stress, which was further proved by the increase in Ca
2+ signal receptors CaM and CDPKs. Therefore, the Ca
2+ signal-related pathway can be used as an important evaluation indicator for the response of
P. edulis to drought stress. However, it is noteworthy that this study was conducted in a laboratory setting, and future field trails with sufficient samples are required to support our conclusions. In the future, cell biology research and multi-omics analysis related to the calcium signaling pathway of
P. edulis under drought stress should be strengthened in order to understand the physiology mechanism of
P. edulis in response to drought stress in depth.
Under drought stress, Ca
2+ of plants often regulates physiological responses such as aquaporin and stomatal movement through signal transduction, which can reduce body water loss [
20,
22]. In this study, PCA analysis of the physiological indicators of
P. edulis affected by drought shows that the first and second principal components of
P. edulis physiological responses to drought stress are X9 (membrane oxidation degree), X12 and X13 (cytoplasmic osmotic adjustment substance) and X1 and X2 (antioxidant enzyme activity), which implied that
P. edulis enhances the osmotic adjustment function by maintaining the stability of cell membrane lipids under drought stress. Pro and Bet, as important substances involved in cell osmotic regulation, can maintain the normal turgor pressure of cells to help plants resist drought stress [
23]. However, the synthesis of related proteins in plants is inhibited in the drought environment, leading to the blocking of the main metabolic pathways of Pro, and the accumulated Pro, as a scavenger of oxygen free radicals, alleviates the oxidative damage caused by drought stress [
24,
25]. In this study, the excessive accumulation of osmotic adjustment substances Pro and Bet is due to the osmotic adjustment of
P. edulis to maintain its own turgor pressure to cope with the pressure generated by environmental stress. In addition, under drought stress, leaf cell membrane permeability is destroyed, leading to an increase in the content of membrane lipid peroxidation product MDA [
26], which implies that MDA accumulation is also involved in the physiological response of
P. edulis to drought stress.
Plant hormones are also important signal substances for plant growth and response to environmental stress. It is generally believed that jasmonic acids (JAs), as endogenous signal molecules, participate in the response of plants to abiotic stress. Studies showed that JAs can enhance the drought resistance of plants [
27,
28]. For example, Yuncong Yao et al. found that under moderate drought stress, JAs can promote the accumulation of dry matter in young apple trees, significantly increase the activity of antioxidant enzymes and promote the accumulation of osmotic substances such as proline [
29]. In addition, Xin et al. found that the JA content of the above-ground part of corn increased under water stress by using gramineous plant corn as the experimental material [
30]. In this study, with the decrease in soil water content, the JA content of
P. edulis leaves increased. These results imply that JA plays an important role in the response of plants to water and drought stress. Under drought stress, ABA is accumulated in the leaves, which induces leaf shedding and leads to the weakening of photosynthesis and transpiration of
P. edulis to adapt to the water-deficient environment [
31]. Similarly, in this study, the ABA content of
P. edulis leaves shows an upward trend under the influence of drought stress. In addition, IAA signals of plants regulate plant physiology and development at all levels. However, in this study, the IAA content of
P. edulis shows a downward trend with the decrease in soil moisture content. The possible reason is that under drought stress,
P. edulis weighs the balance of defense and growth, and through the coordination of hormones such as JA, ABA and IAA, slows down metabolism and plant growth to deal with environmental pressure.
Rubp is an important substance in the Calvin cycle of photosynthesis. RubisCO is the first reaction enzyme that catalyzes the carbon cycle and photorespiration of plants, and plays a key role in photosynthesis, which is a key substance for plant carbon assimilation in photosynthesis [
32,
33,
34]. Studies show that the primary metabolism of
P. edulis is significantly inhibited under drought stress [
35]. Therefore, more attention should be paid to research on the photosynthesis of
P. edulis under drought stress.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}