
Conifer Forest Dynamics in the Iberian Pyrenees during the Middle Ages


Abstract
:1. Introduction
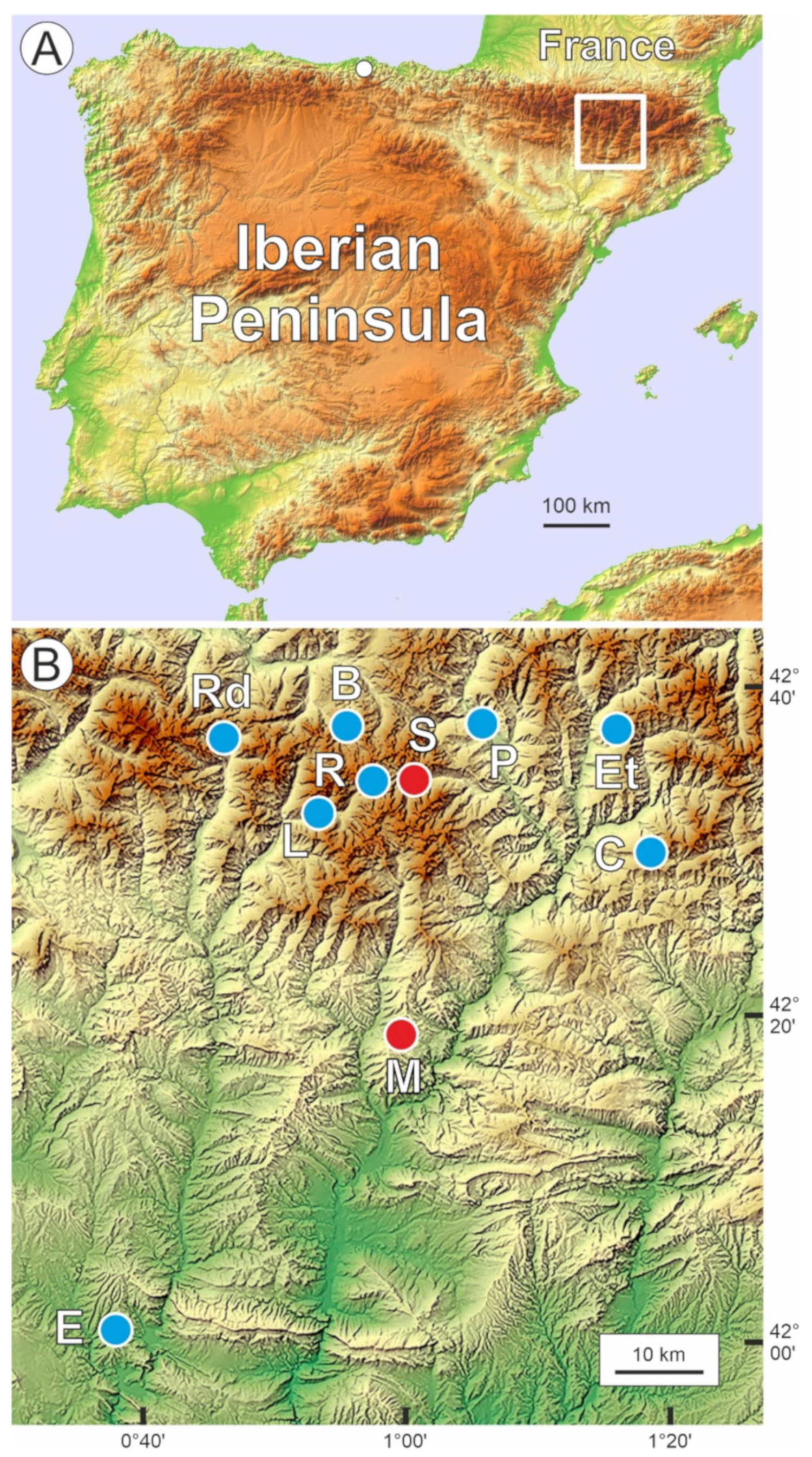
2. Study Area
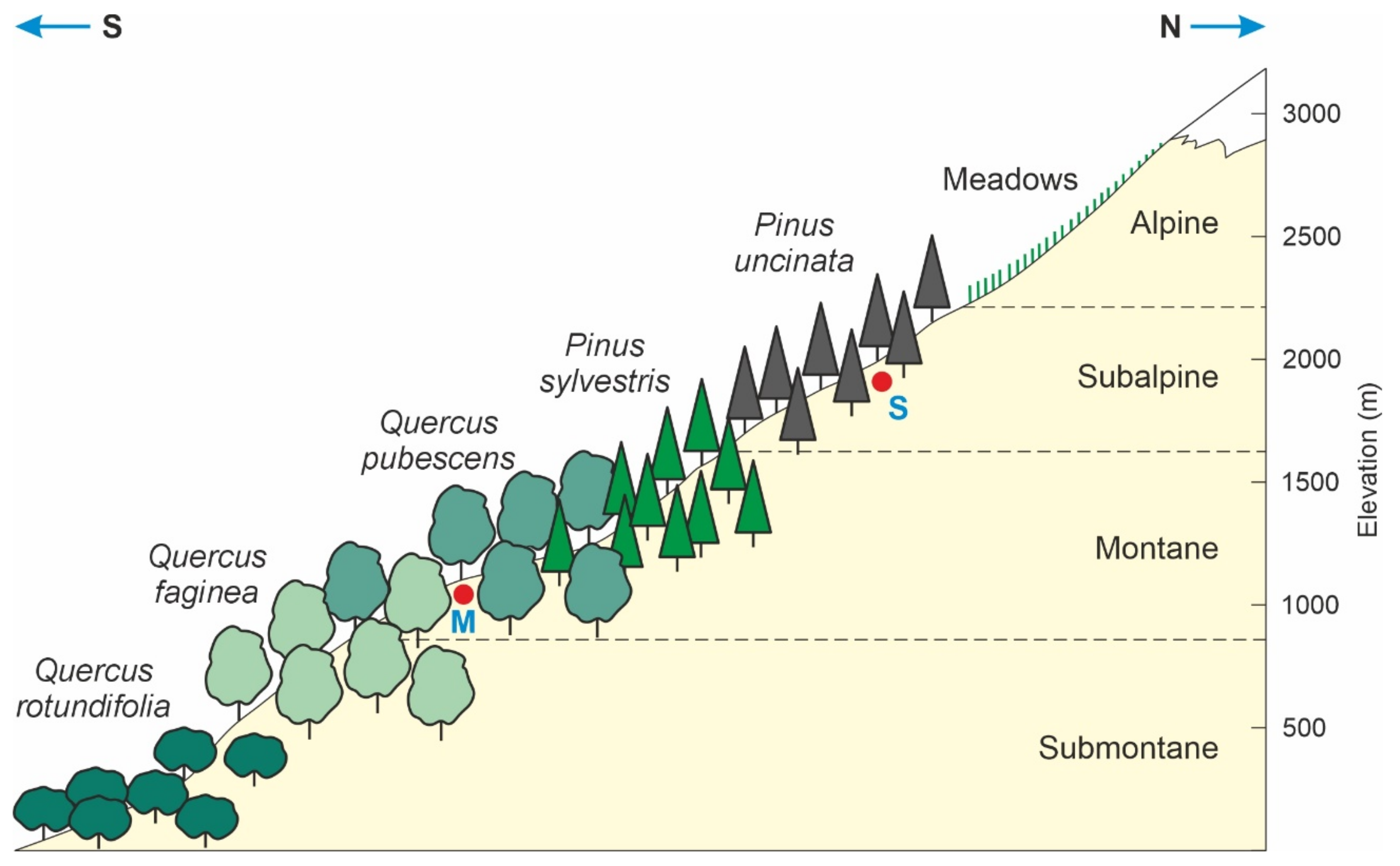
2.1. Altitudinal Forest Zonation
2.2. Study Sites
3. Methods
4. Forest Dynamics
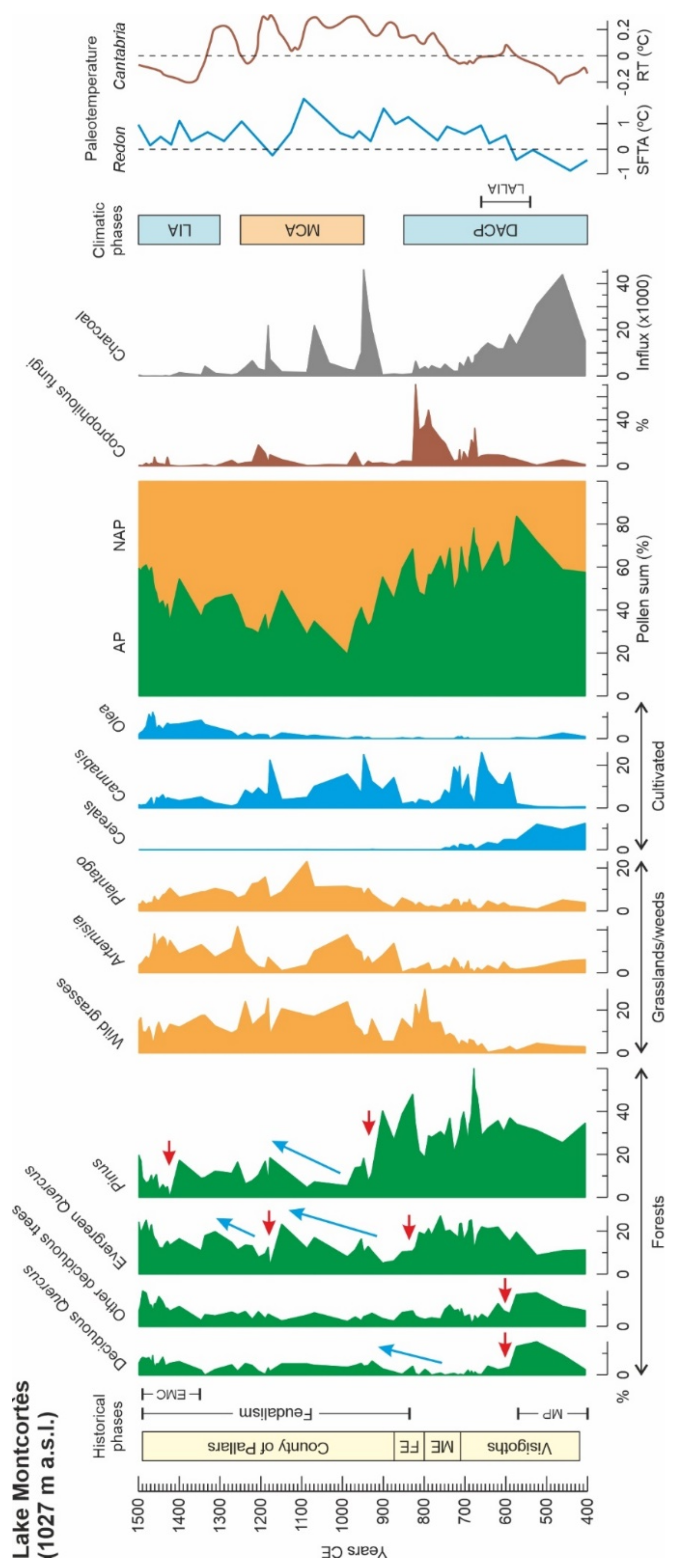
4.1. Montcortès
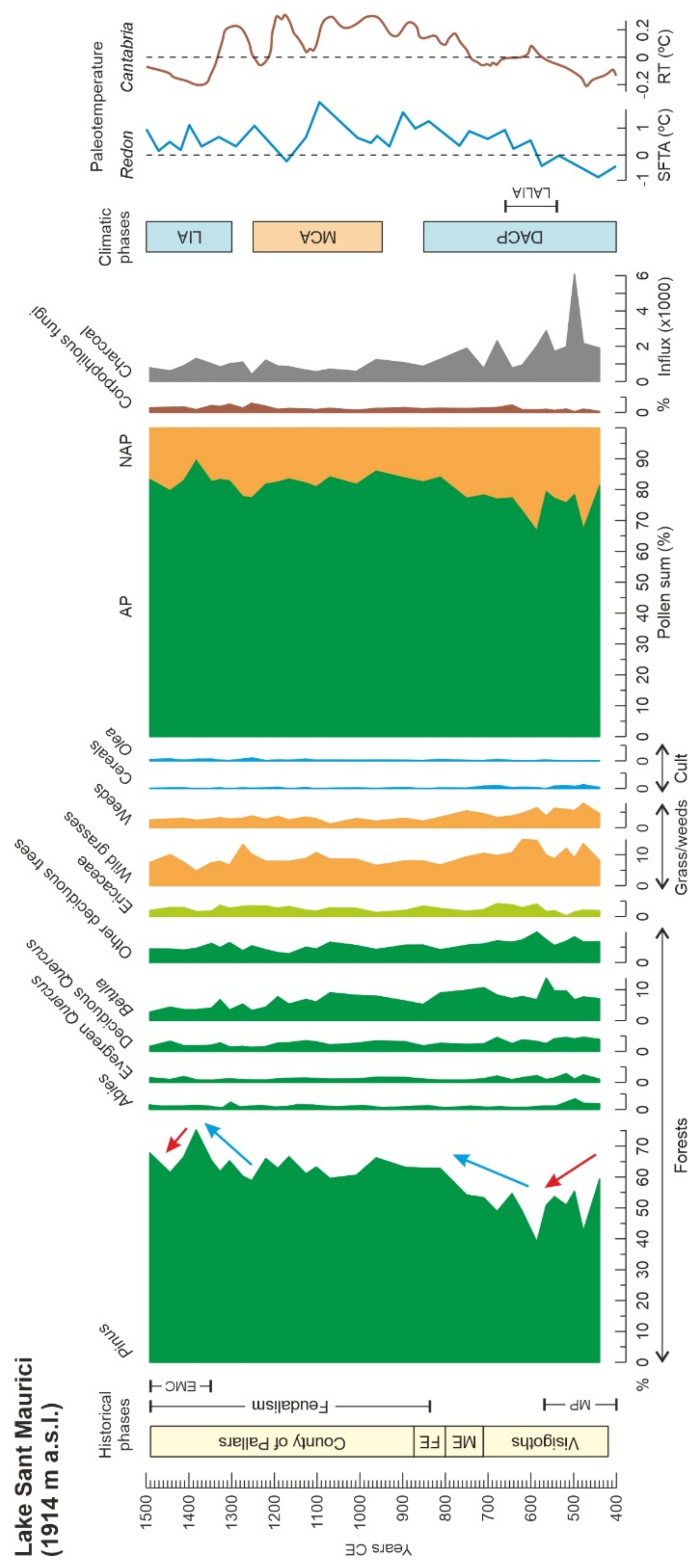
4.2. Sant Maurici
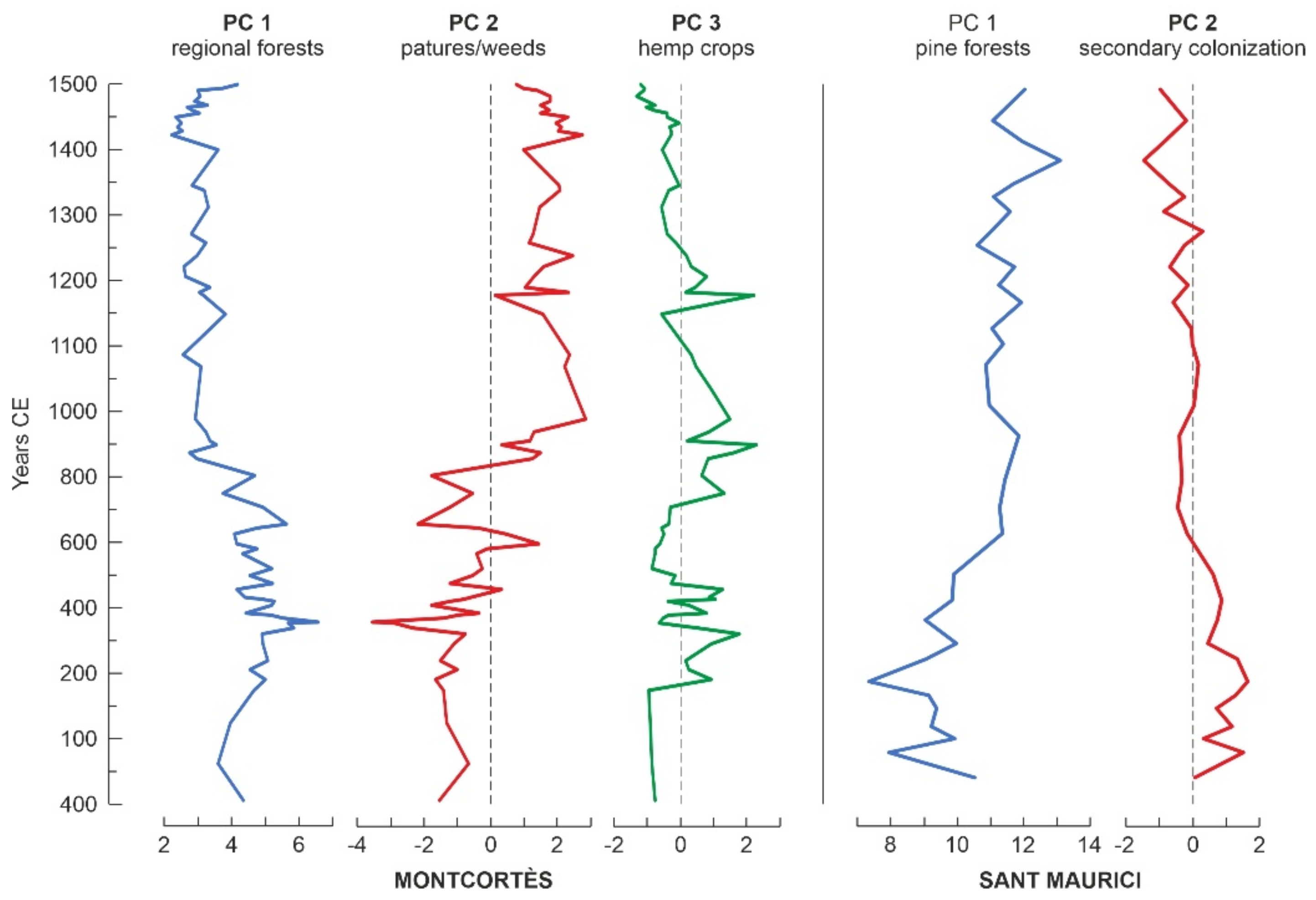
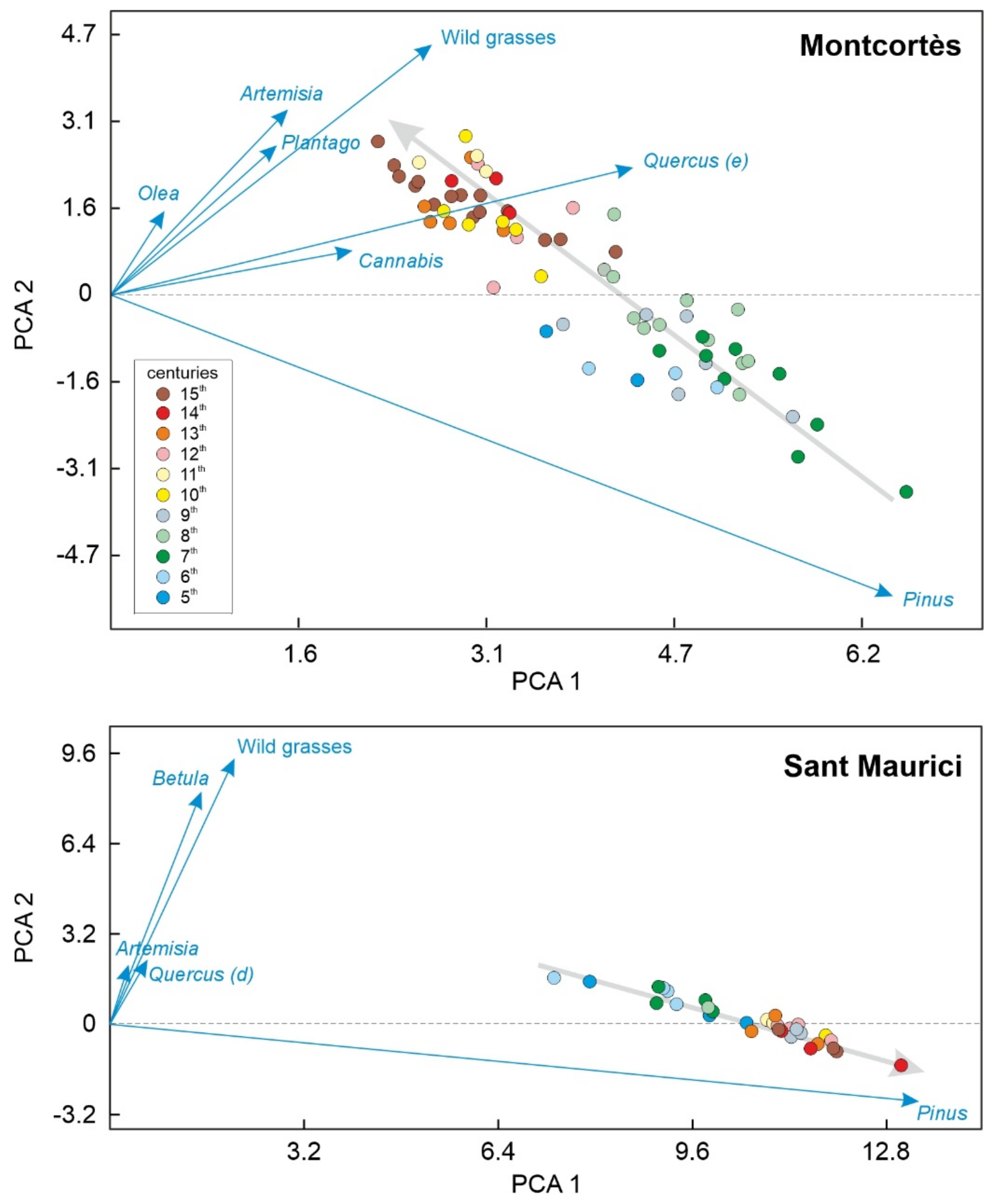
4.3. Numerical Analysis
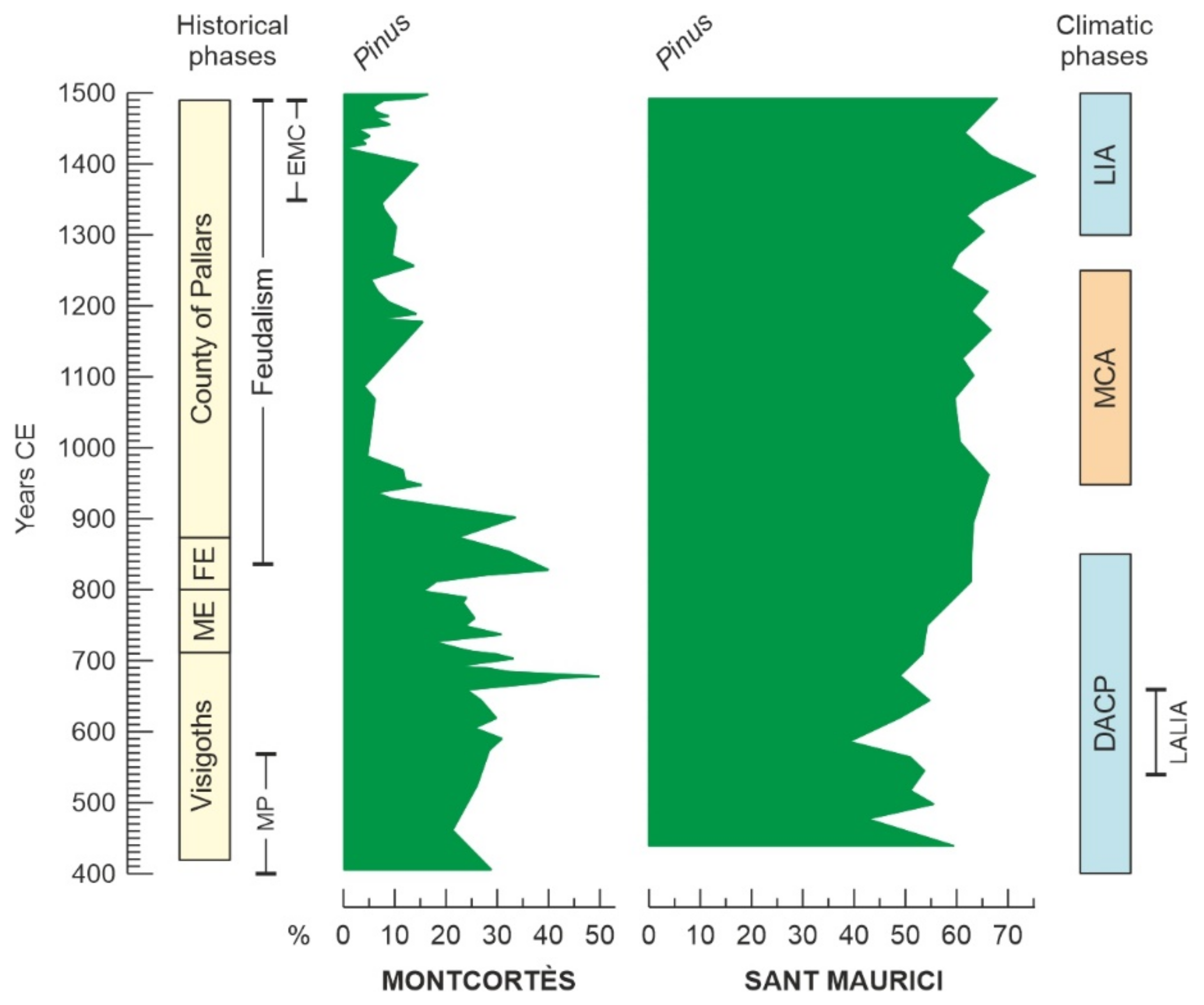
4.4. Conifer Forests
5. Regional Comparisons
6. Conservation
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Medieval History and Culture of the Pallars Region
References
- Halsall, G. Barbarian Migrations and the Roman West; Cambridge University Press: Cambridge, UK, 2008; pp. 376–568. [Google Scholar]
- Marugan, C.M.; Oliver, J. El Pallars Medieval. In Història del Pallars. Dels Orígens als Nostres Dies; Marugan, C.M., Rapalino, V., Eds.; Pagès Editors: Lleida, Spain, 2005; pp. 45–86. [Google Scholar]
- González-Sampériz, P.; Aranbarri, J.; Pérez-Sanz, A.; Gil-Romera, G.; Moreno, A.; Leunda, M.; Sevilla-Callejo, M.; Corella, J.P.; Morellón, M.; Oliva, M.; et al. Environmental and climate change in the southern Central Pyrenees since the Last Glacial Maximum: A review from the lake records. Catena 2017, 149, 668–688. [Google Scholar] [CrossRef]
- González-Sampériz, P.; Montes, L.; Aranbarri, J.; Leunda, M.; Domingo, R.; Laborda, R.; Sanjuan, Y.; Gil-Romera, G.; Lasanta, T.; García-Ruiz, J.M. Escenarios, tiempo e indicadores paleoambientales para la identificación del Antropoceno en el paisaje vegetal del Pirineo central (NE Iberia). Cuad. Investig. Geogr. 2019, 45, 167–193. [Google Scholar] [CrossRef] [Green Version]
- García-Ruiz, J.M.; Tomás-Faci, G.; Diarte-Blasco, P.; Montes, L.; Domingo, R.; Sebastián, M.; Lasanta, T.; González-Sampériz, P.; López-Moreno, J.I.; Arnáez, J.; et al. Transhumance and long-term deforestation in the subalpine belt of the central Spanish Pyrenees: An interdisciplinary approach. Catena 2020, 195, 104744. [Google Scholar] [CrossRef]
- Rull, V.; Vegas-Vilarrúbia, T. A spatiotemporal gradient in the anthropization of Pyrenean landscapes. Preliminary report. Quat. Sci. Rev. 2021, 258, 106909. [Google Scholar] [CrossRef]
- Gassiot, E.; Mazzucco, N.; Clemente, I.; Rodríguez, D.; Obea, L.; Quesada, M.; Díaz, S. The beginning of high mountain occupations in the Pyrenees. Human settlements and mobility from 18,000 cal BC to 2000 cal BC. In High Mountain Conservation in a Changing World; Catalan, J., Ninot, J.M., Aniz, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 75–105. [Google Scholar]
- Rull, V.; Vegas-Vilarrúbia, T.; Corella, J.P.; Trapote, M.C.; Montoya, E.; Valero-Garcés, B. A unique Pyrenean varved record provides a detailed reconstruction of Mediterranean vegetation and land use dynamics over the last three millennia. Quat. Sci. Rev. 2021, 268, 107128. [Google Scholar] [CrossRef]
- Gassiot, E. Montañas Humanizadas. Arqueología del Passtoralismo en el Parque Nacional d’Aigüestortes i Estany de Sant Maurici. Organismo Autónomo de Parques Nacionales: Madrid, Spain, 2016. [Google Scholar]
- Rull, V.; Cañellas-Boltà, N.; Vegas-Vilarrrúbia, T. Late-Holocene forest resilience in the central Pyrenean highlands as deduced from pollen analysis of Lake Sant Maurici sediments. Holocene 2021, 31, 1797–1803. [Google Scholar] [CrossRef]
- Rull, V.; Vegas-Vilarrúbia, T.; Corella, J.P.; Valero-Garcés, B. Bronze Age to Medieval vegetation dynamics and landscape anthropization in the southern-central Pyrenees. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 571, 110392. [Google Scholar] [CrossRef]
- Marugan, C.M.; Rapalino, V. Història del Pallars. Dels Orígens als Nostres Dies; Pagès Editors: Lleida, Spain, 2005. [Google Scholar]
- Rull, V. Cultural development of the Pallars region (NE Iberian Peninsula) from the Bronze Age to the present. PaleorXiv 2021. [Google Scholar] [CrossRef]
- Vigo, J. Flora i Vegetació de l’Alta Muntanya Catalana; Institut d’Estudis Catalans: Barcelona, Spain, 2008. [Google Scholar]
- Carrillo, E.; Aniz, M. Guía del Parque Nacional de Aigüestortes i Estany de Sant Maurici; Organismo Autónomo de Parques Nacionales: Madrid, Spain, 2013. [Google Scholar]
- Ninot, J.M.; Carrillo, E.; Ferré, A. The Pyrenees. In Vegetation of the Iberian Peninsula; Loidi, J., Ed.; Springer Nature: Cham, Switzerland, 2017; pp. 323–366. [Google Scholar]
- Mercadé, A.; Vigo, J.; Rull, V.; Vegas-Vilarrúbia, T.; Garcés, S.; Cañellas-Boltà, N. Vegetation and landscape around Lake Montcortès (Catalan pre-Pyrenees) as a tool for palaeoecological studies of lake sediments. Colect. Bot. 2013, 2013. 32, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Rull, V.; González-Sampériz, P.; Corella, J.P.; Morellón, M.; Giralt, S. Vegetation changes in the southern Pyrenean flank during the last millennium in relation to climate and human activities: The Montcortès lacustrine record. J. Paleolimnol. 2011, 46, 387–404. [Google Scholar] [CrossRef] [Green Version]
- Montoya, E.; Rull, V.; Vegas-Vilarrúbia, T.; Corella, J.P.; Giralt, S.; Valero-Garcés, B. Grazing activities in the southern central Pyrenees during the last millennium as deduced from the non-pollen palynomorphs (NPP) record of Lake Montcortès. Rev. Palaeobot. Palynol. 2018, 254, 8–19. [Google Scholar] [CrossRef] [Green Version]
- Cañellas-Boltà, N.; Rull, V.; Vigo, J.; Mercadé, A. Modern pollen-vegetation relationships along an altitudinal transect in the central Pyrenees (south-western Europe). Holocene 2009, 19, 1185–1200. [Google Scholar] [CrossRef] [Green Version]
- López-Vila, J.; Montoya, E.; Cañellas-Boltà, N.; Rull, V. Modern non-pollen palymomorphs sedimentation along an elevational gradient in the southern-central Pyrenees (southwestern Europe) as a tool for Holocene paleoecological reconstruction. Holocene 2014, 24, 1757–1770. [Google Scholar] [CrossRef] [Green Version]
- Rull, V.; Trapote, M.C.; Safont, E.; Cañellas-Boltà, N.; Pérez-Zanón, N.; Sigró, J.; Buchaca, T.; Vegas-Vilarrúbia, T. Seasonal patterns of pollen sedimentation in Lake Montcortès (Central Pyrenees) and potential applications to high-resolution paleoecology: A 2-year pilot study. J. Paleolimnol. 2017, 57, 96–108. [Google Scholar] [CrossRef] [Green Version]
- Catalan, J.; Pèlachs, A.; Gassiot, E.; Antolín, F.; Ballesteros, A.; Batalla, M.; Burjachs, F.; Buchaca, T.; Camarero, L.; Clemente, I.; et al. Interacción Entre Clima y Ocupación Humana en la Configuración del Paisaje Vegetal en el Parque Nacional de Aigüestortes i Estany de Sant Maurici a lo Largo de los Últimos 15.000 Años. Proyectos de Investigación en Parques Nacionales: 2009–2012; Organismo Autónomo de Parques Nacionales: Madrid, Spain, 2013; pp. 71–92. [Google Scholar]
- Büntgen, U.; Krusic, P.J.; Verstege, A.; Sangüesa-Barreda, G.; Wagner, S.; Camarero, J.J.; Charpentier, F.; Zorita, E.; Oppemheimer, C.; Konter, O.; et al. New tree-ring evidence from the Pyrenees reveals western Mediterranean climate variability since Medieval times. J. Clim. 2017, 30, 5295–5318. [Google Scholar] [CrossRef]
- Martín-Chivelet, J.; Muñoz-García, M.B.; Edwards, R.L.; Edwards, R.L.; Turrero, M.J.; Ortega, A.I. Land surface temperature changes in Northern Iberia since 4000 yr BP, based on δ13C of speleothems. Glob. Planet. Chang. 2011, 77, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Gribbin, J.; Lamb, H.H. Climatic Change. In Climatic Change in Historical Times; Gribbin, J., Ed.; Cambridge University Press: Cambridge, UK, 1978; pp. 68–82. [Google Scholar]
- Mann, M.E.; Zhang, Z.; Rutherford, S.; Bradley, R.S.; Hughes, M.K.; Shindel, D.; Ammann, C.; Faluvegi, G.; Ni, F. Global signatures and dynamic origins of the Little Ice Age and the Medieval Climate Anomaly. Science 2009, 326, 1256–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büntgen, U.; Myglan, V.S.; Charpentier, F.; McCormick, M.; Di Cosmo, N.; Sigl, M.; Jungclaus, J.; Wagner, S.; Wagner, S.; Krusic, P.J.; et al. Cooling and societal change during the Late Antique Little Ice Age from 536 to around 660 AD. Nat. Geosci. 2016, 9, 231–236. [Google Scholar] [CrossRef]
- Helama, S.; Jones, P.D.; Briffa, K.R. Dark Ages Cold Period: A literature review and directions for future research. Holocene 2017, 27, 1600–1606. [Google Scholar] [CrossRef] [Green Version]
- Dubois, H.; Verkasalo, E.; Claessens, H. Potential of Birch (Betula pendula Roth and B. pubescens Ehrh.) for forestry and forest-based industry sector within the changing climate and socio-economic context of Western Europe. Forests 2020, 11, 336. [Google Scholar] [CrossRef] [Green Version]
- Riera, S.; Wansard, G.; Julià, R. 2000-year environmental history of a karstic lake in the Mediterranean Pre-Pyrenees: The Estanya lake (Spain). Catena 2004, 55, 293–324. [Google Scholar] [CrossRef]
- Riera, S.; López-Sáez, J.A.; Julià, R. Lake responses to historical land use changes in northern Spain: The contribution of non-pollen palynomorphs in a multiproxy study. Rev. Palaeobot. Palynol. 2006, 141, 127–137. [Google Scholar] [CrossRef]
- Pèlachs, A.; Pérez-Obiol, R.; Ninyerola, M.; Nadal, J. Landscape dynamics of Abies and Fagus in the southern Pyrenees during the last 2200 years as a result of anthropogenic impacts. Rev. Palaeobot. Palynol. 2009, 156, 337–349. [Google Scholar] [CrossRef]
- Cunill, R.; Soriano, J.N.; Bal, M.C.; Pèlachs, A.; Rodriguez, J.M.; Pérez-Obiol, R. Holocene high-altitude vegetation dynamics in the Pyrenees: A pedoanthracology contribution to an interdisciplinary approach. Quat. Int. 2013, 289, 60–70. [Google Scholar] [CrossRef]
- Garcés-Pastor, S.; Cañellas-Boltà, N.; Pèlachs, A.; Soriano, J.M.; Pérez-Obiol, R.; Pérez-Haase, A.; Calero, M.A.; Oriol, A.; Escolà, N.; Vegas-Vilarrúbia, T. Environmental history and vegetation dynamics in response to climate variations and human pressure during the Holocene in Bassa Nera, Central Pyrenees. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 479, 48–60. [Google Scholar] [CrossRef]
- Pèlachs, A.; Soriano, J.M.; Nadal, J.; Esteban, A. Holocene environmental history and human impact in the Pyrenees. Contrib. Sci. 2007, 3, 421–429. [Google Scholar]
- Rull, V. Microrefugia. J. Biogeogr. 2008, 36, 481–484. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rull, V.; Vegas-Vilarrúbia, T. Conifer Forest Dynamics in the Iberian Pyrenees during the Middle Ages. Forests 2021, 12, 1685. https://doi.org/10.3390/f12121685
Rull V, Vegas-Vilarrúbia T. Conifer Forest Dynamics in the Iberian Pyrenees during the Middle Ages. Forests. 2021; 12(12):1685. https://doi.org/10.3390/f12121685
Chicago/Turabian StyleRull, Valentí, and Teresa Vegas-Vilarrúbia. 2021. "Conifer Forest Dynamics in the Iberian Pyrenees during the Middle Ages" Forests 12, no. 12: 1685. https://doi.org/10.3390/f12121685
APA StyleRull, V., & Vegas-Vilarrúbia, T. (2021). Conifer Forest Dynamics in the Iberian Pyrenees during the Middle Ages. Forests, 12(12), 1685. https://doi.org/10.3390/f12121685