
Impacts of Forest Fire on Understory Species Diversity in Canary Pine Ecosystems on the Island of La Palma

 , , , , and
, , , , and 
Abstract
:1. Introduction
- (A)
- We hypothesized that the highest levels of species alpha and beta diversity would be at intermediate fire intensity;
- (B)
- Lowest abundance of L. campylocladus ssp. hillebrandii is expected at intermediate fire intensities because of the persisting dominance of P. canariensis at low fire intensities and the damage to the seedbank, unfavorable soil conditions, and erosion at high intensities;
- (C)
- As this legume (Lotus) contributes to N-fixing and nutrient availability, we expect an interaction between fire intensity and legume biomass in the effect on biodiversity patterns.
2. Materials and Methods
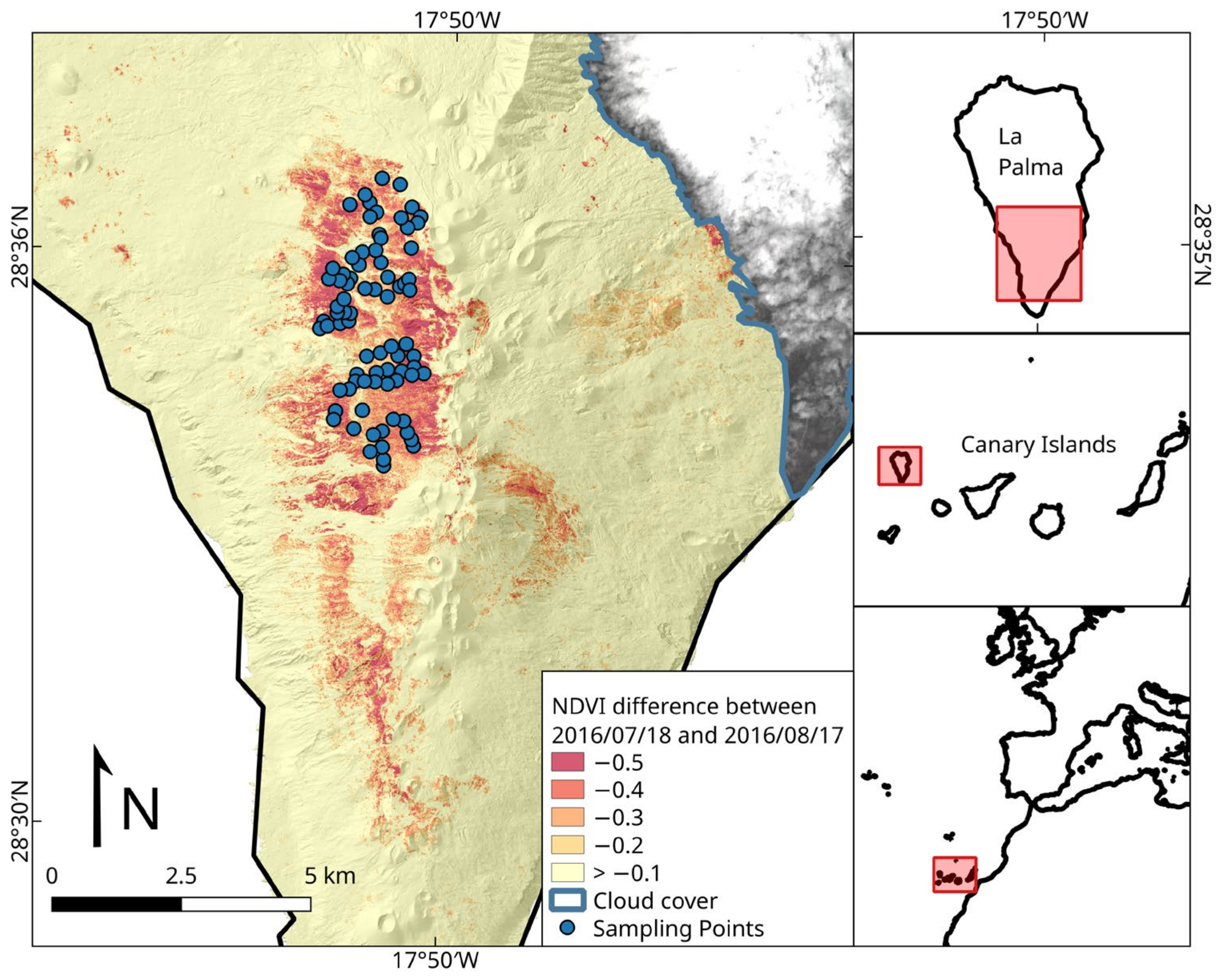
2.1. Study Area
2.2. Remote Sensing
2.3. Field Sampling and Other Explanatory Variables
2.4. Statistical Analyses
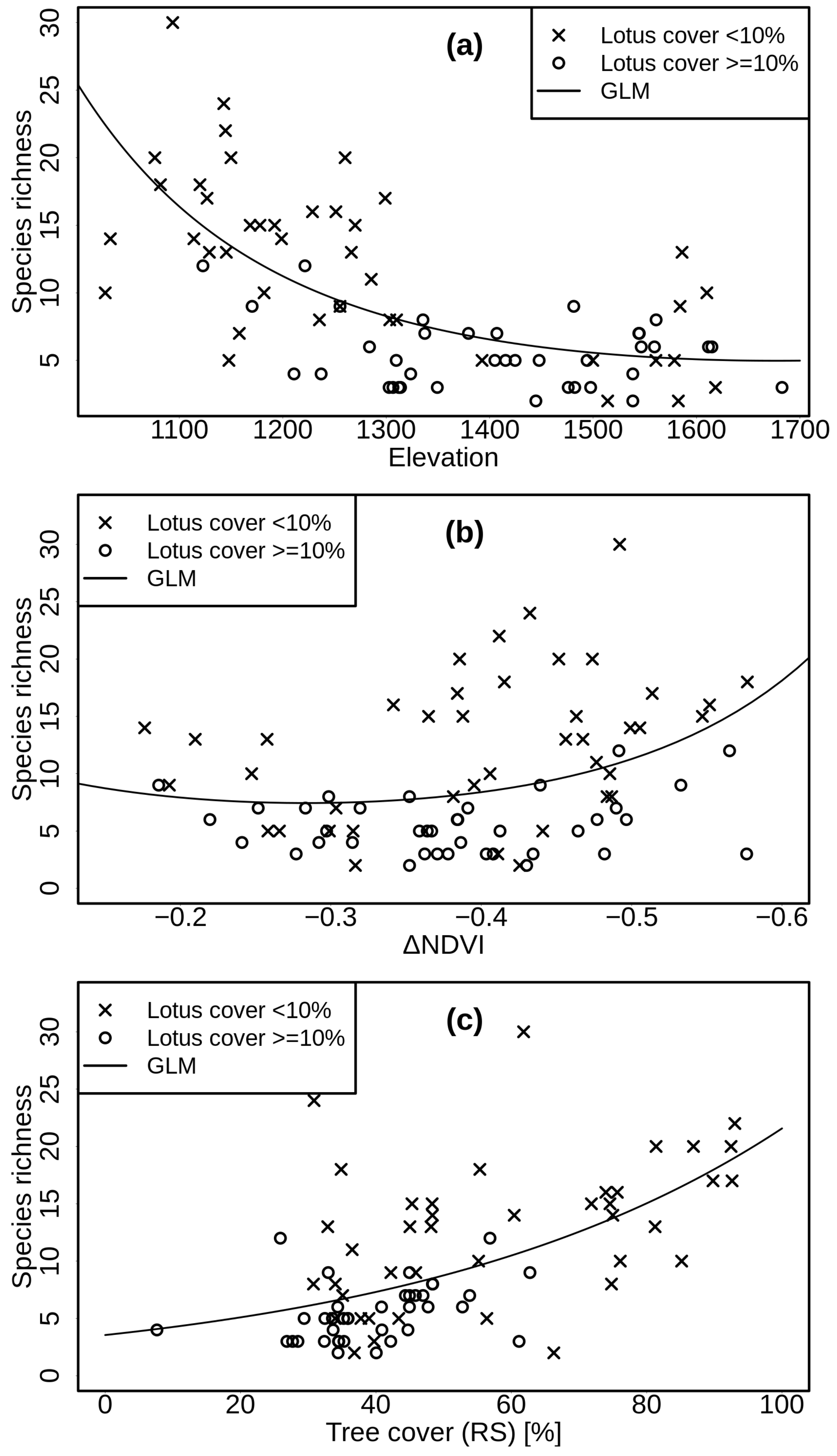
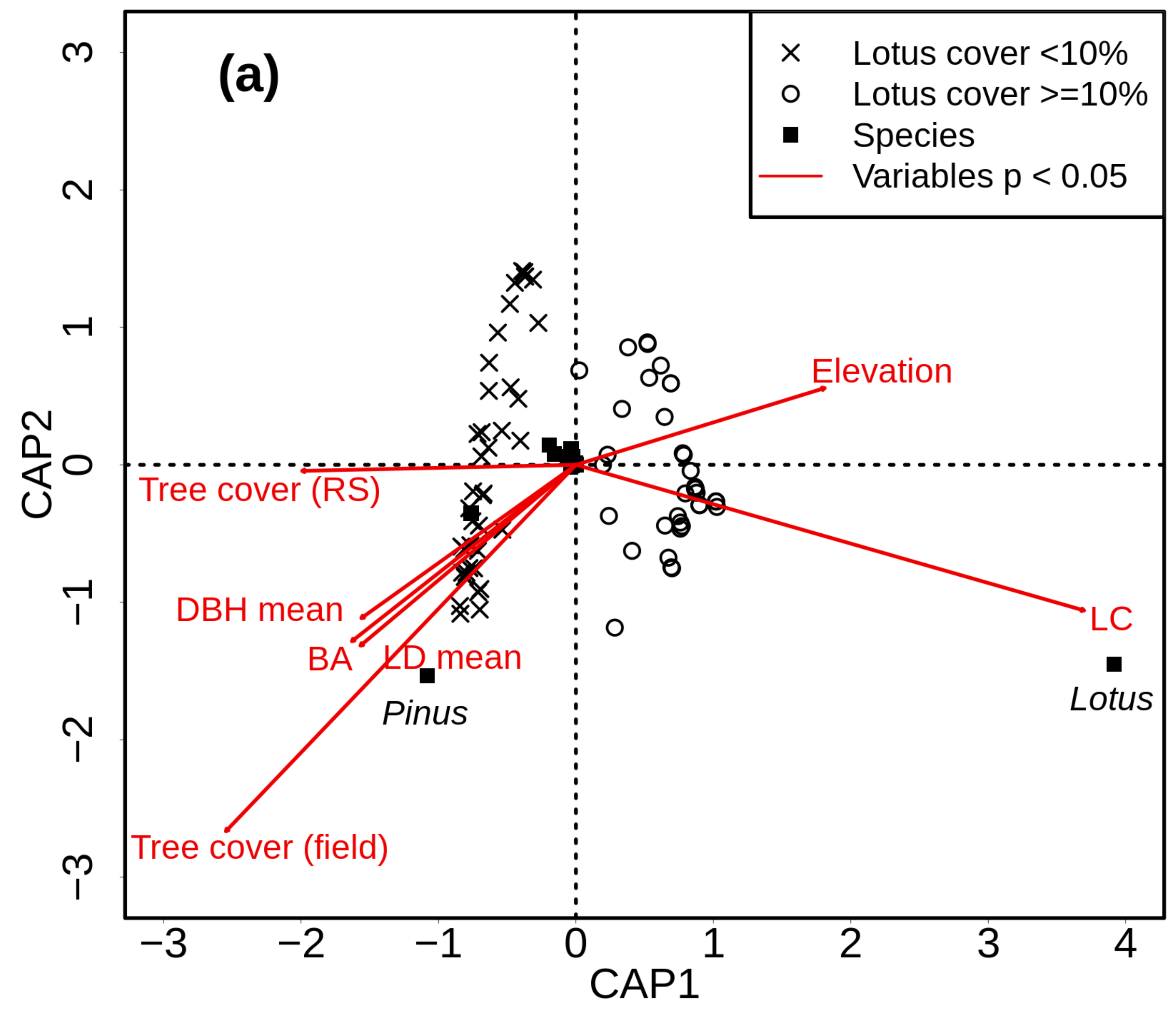
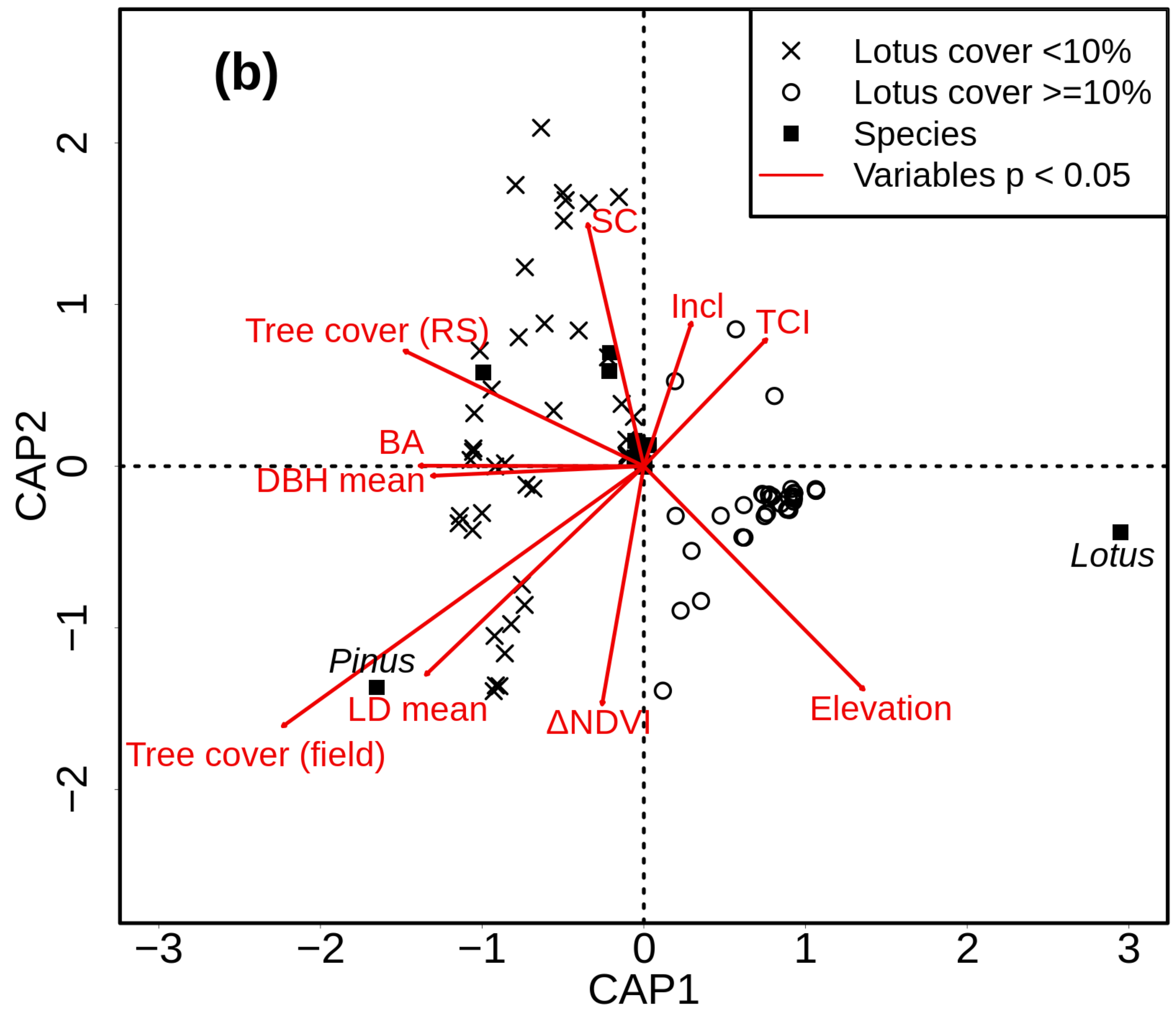
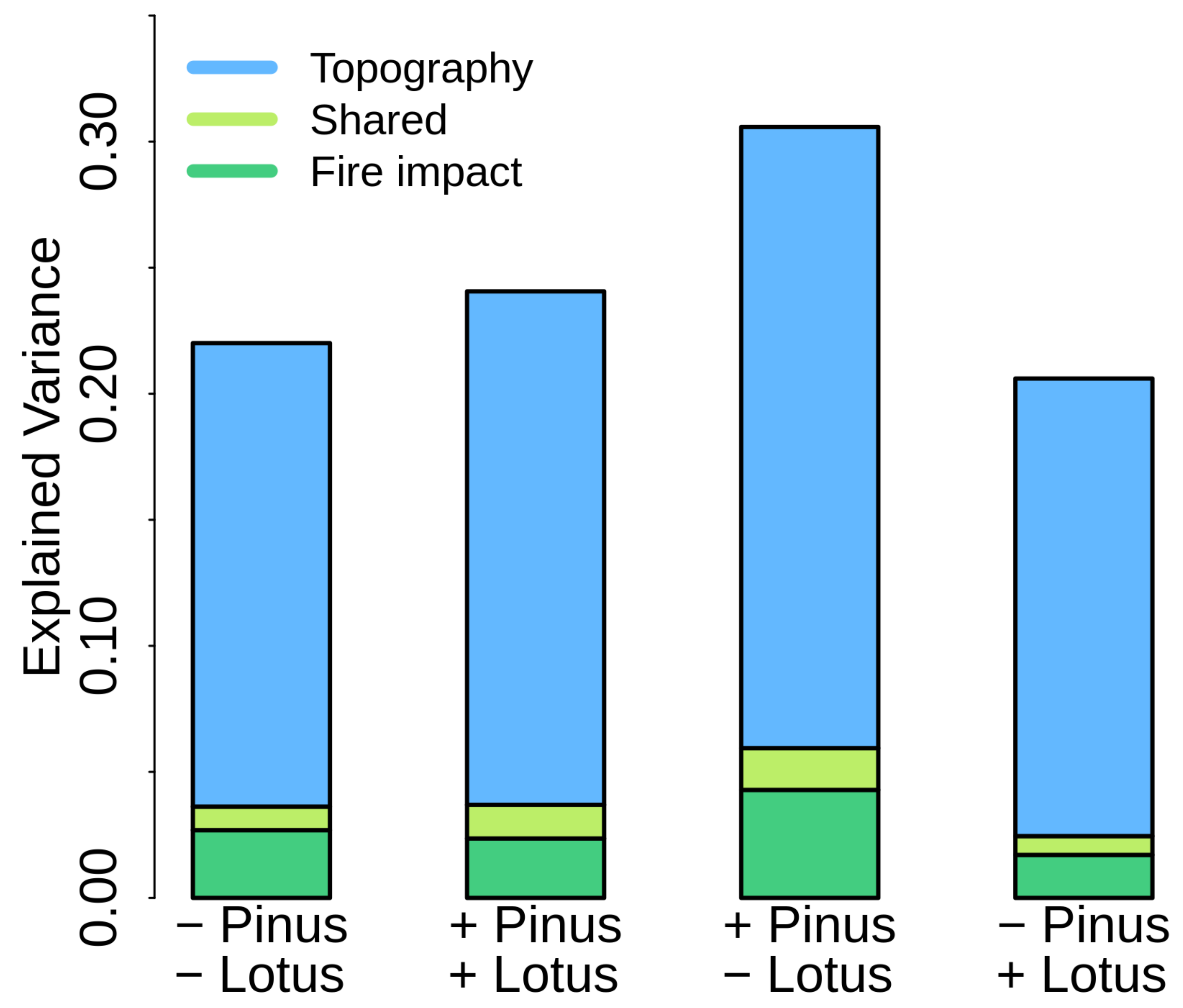
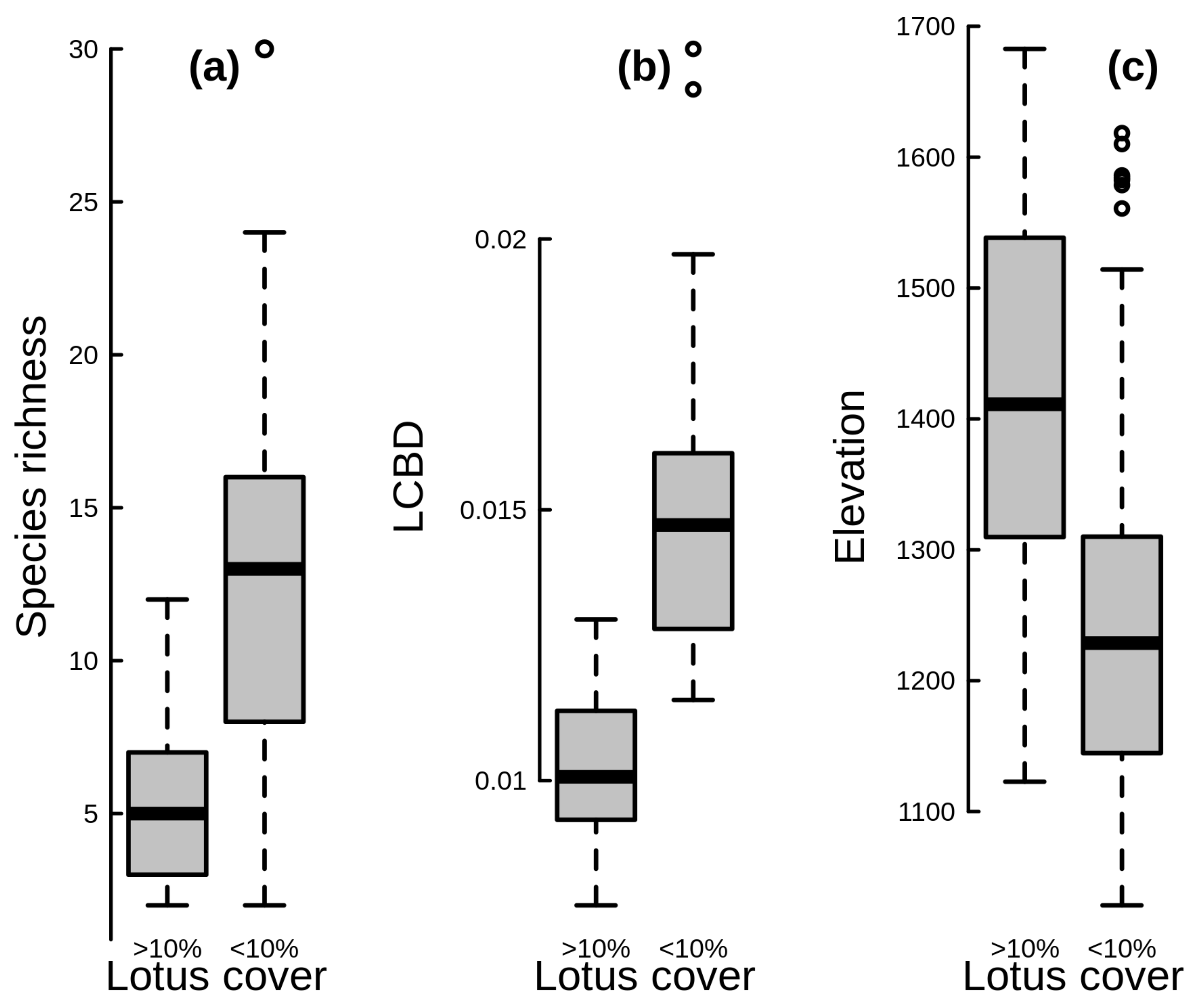
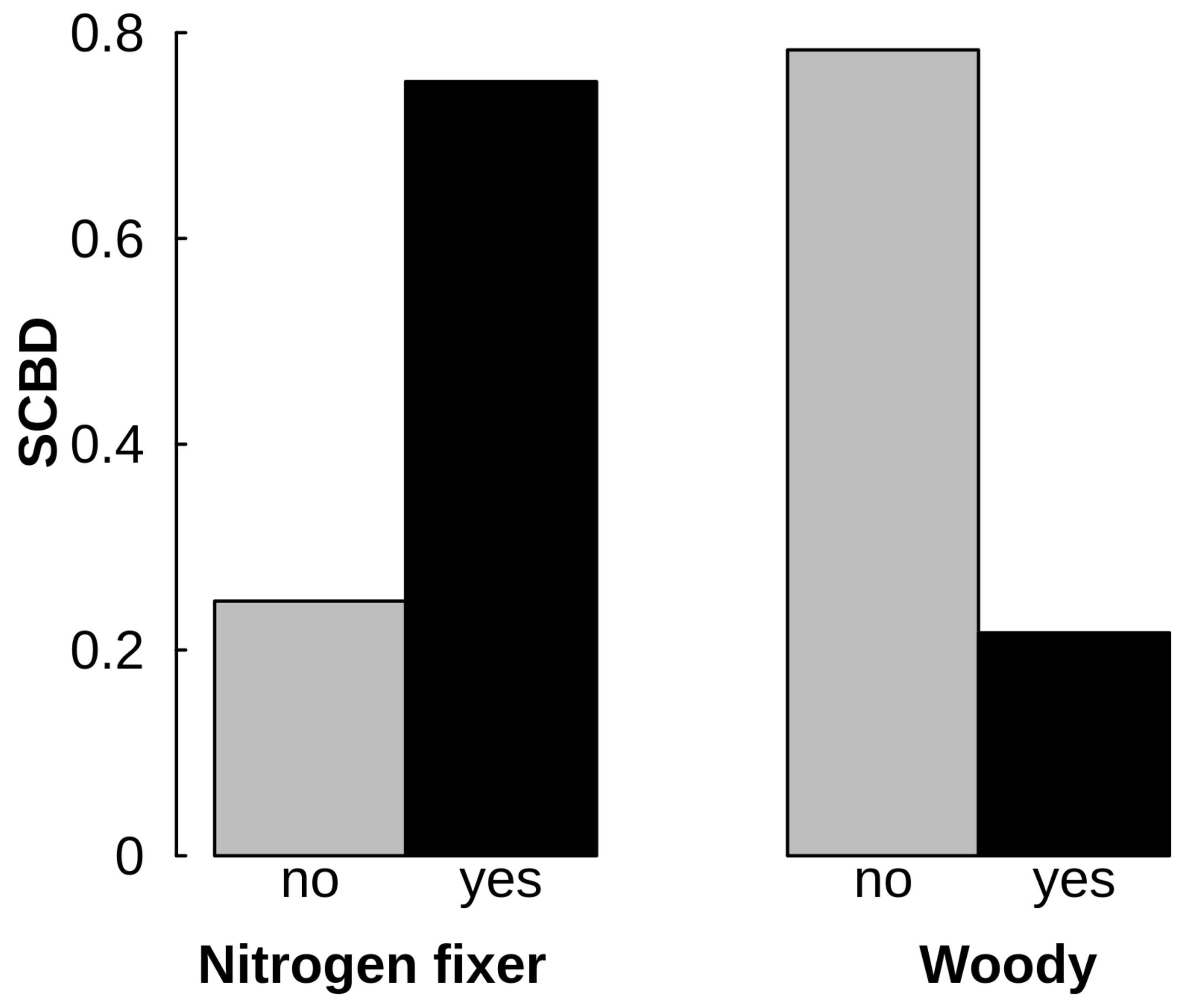
3. Results
4. Discussion
4.1. Alpha Diversity (Species Richness)
4.2. Beta Diversity
4.3. Role of Lotus campylocladus ssp. hillebrandii
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bowman, D.M.; Balch, J.K.; Artaxo, P.; Bond, W.J.; Carlson, J.M.; Cochrane, M.A.; D’Antonio, C.M.; DeFries, R.S.; Doyle, J.C.; Harrison, S.P. Fire in the earth system. Science 2009, 324, 481–484. [Google Scholar] [CrossRef]
- Bond, W.J.; Woodward, F.I.; Midgley, G.F. The global distribution of ecosystems in a world without fire. New Phytol. 2005, 165, 525–538. [Google Scholar] [CrossRef]
- Bond, W.J.; Keeley, J.E. Fire as a global ‘herbivore’: The ecology and evolution of flammable ecosystems. Trends Ecol. Evol. 2005, 20, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Keeley, J.E. A burning story: The role of fire in the history of life. Bioscience 2009, 59, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.G.; Hargrove, W.W.; Gardner, R.H.; Romme, W.H. Effects of fire on landscape heterogeneity in Yellowstone National Park, Wyoming. J. Veg. Sci. 1994, 5, 731–742. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.G.; Romme, W.H. Landscape dynamics in crown fire ecosystems. Landsc. Ecol. 1994, 9, 59–77. [Google Scholar] [CrossRef]
- Pausas, J.G.; Llovet, J.; Rodrigo, A.; Vallejo, R. Are wildfires a disaster in the Mediterranean basin?—A review. Int. J. Wildland Fire 2008, 17, 713–723. [Google Scholar] [CrossRef]
- Schimmel, J.; Granstrom, A. Fire severity and vegetation response in the boreal Swedish forest. Ecology 1996, 77, 1436–1450. [Google Scholar] [CrossRef]
- Cowan, A.D.; Smith, J.E.; Fitzgerald, S.A. Recovering lost ground: Effects of soil burn intensity on nutrients and ectomycorrhiza communities of ponderosa pine seedlings. For. Ecol. Manag. 2016, 378, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Butnor, J.R.; Johnsen, K.H.; Maier, C.A.; Nelson, C.D. Intra-annual variation in soil C, N and nutrients pools after prescribed fire in a Mississippi longleaf pine (Pinus palustris Mill.) plantation. Forests 2020, 11, 181. [Google Scholar] [CrossRef] [Green Version]
- Otto, R.; García-del-Rey, E.; Muñoz, P.G.; Fernández-Palacios, J.M. The effect of fire severity on first-year seedling establishment in a Pinus canariensis forest on Tenerife, Canary Islands. Eur. J. For. Res. 2010, 129, 499–508. [Google Scholar] [CrossRef]
- Méndez, J.; Morales, G.; de Nascimento, L.; Otto, R.; Gallardo, A.; Fernández-Palacios, J.M. Understanding long-term post-fire regeneration of a fire-resistant pine species. Ann. For. Sci. 2015, 72, 609–619. [Google Scholar] [CrossRef] [Green Version]
- Zagas, T.; Ganatsas, P.; Tsitsoni, T.; Tsakaldimi, M. Post-fire regeneration of Pinus halepensis Mill. stands in the Sithonia Peninsula, Northern Greece. Plant Ecol. 2004, 171, 91–99. [Google Scholar] [CrossRef]
- Spanos, I.A.; Daskalakou, E.N.; Thanos, C.A. Post-fire, natural regeneration of Pinus brutia forests in Thasos island, Greece. Acta Oecol. 2000, 21, 2013–2020. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, J.J. Ecology of sprouting in woody plants: The persistence niche. Trends Ecol. Evol. 2001, 16, 45–51. [Google Scholar] [CrossRef]
- Tsitsoni, T. Conditions determining natural regeneration after wildfires in the Pinus halepensis (Miller, 1768) forests of Kassandra Peninsula (North Greece). For. Ecol. Manag. 1997, 92, 199–208. [Google Scholar] [CrossRef]
- Romme, W.H.; Boyce, M.S.; Gresswell, R.E.; Merrill, E.H.; Minshall, G.W.; Whitlock, C.; Turner, M.G. Twenty years after the 1988 Yellowstone fires: Lessons about disturbance and ecosystems. Ecosystems 2011, 14, 1196–1215. [Google Scholar] [CrossRef]
- Jentsch, A.; White, P. A theory of pulse dynamics and disturbance in ecology. Ecology 2019, 100, e02734. [Google Scholar] [CrossRef]
- Höllermann, P. The impact of fire in the Canarian ecosystems 1983–1998. (Auswirkungen des Feuers in Ökosystemen der Kanarischen Inseln 1983–1998). Erdkunde 2000, 54, 70–75. [Google Scholar] [CrossRef]
- Gell-Mann, M. Complex adaptive systems. In Complexity: Metaphors, Models and Reality, 1st ed.; Cowan, G., Pines, D., Meltzer, D., Eds.; Addison-Wesley: Boston, MA, USA, 1994; Volume XIX, pp. 17–45. [Google Scholar]
- Arévalo, J.R.; Naranjo Cigala, A.; Fernández-Palacios, J.M.; Fernández-Lugo, S. Ecology and management of natural and reforested Canary Island pine stands. In Woodlands: Ecology, Managment and Conservation, 1st ed.; Wallace, E.R., Ed.; Nova Science Publishers: New York, NY, USA, 2011; pp. 1–23. [Google Scholar]
- Climent, J.; Tapias, R.; Pardos, J.A.; Gil, L. Fire adaptations in the Canary Islands pine (Pinus canariensis). Plant Ecol. 2004, 171, 185–196. [Google Scholar] [CrossRef]
- Molina-Terrén, D.M.; Fry, D.L.; Grillo, F.F.; Cardil, A.; Stephens, S.L. Fire history and management of Pinus canariensis forests on the western Canary Islands Archipelago, Spain. For. Ecol. Manag. 2016, 382, 184–192. [Google Scholar] [CrossRef]
- Stephens, S.L.; Ruth, L.W. Federal forest-fire policy in the United States. Ecol. Appl. 2005, 15, 532–542. [Google Scholar] [CrossRef] [Green Version]
- Steel, Z.L.; Koontz, M.J.; Safford, H.D. The changing landscape of wildfire: Burn pattern trends and implications for California’s yellow pine and mixed conifer forests. Landsc. Ecol. 2018, 33, 1159–1176. [Google Scholar] [CrossRef]
- Mora, J.L.; Armas-Herrera, C.M.; Guerra, J.A.; Arbelo, C.D.; Rodríguez-Rodríguez, A.; Notario del Pino, J.S. A comparative study of long-term effects on fire-affected volcanic soils in two different ecosystems in the Canary Islands. Land Degrad. Dev. 2016, 27, 1489–1500. [Google Scholar] [CrossRef]
- Kutiel, P.; Shaviv, A. Effects of soil type, plant composition and leaching on soil nutrients following a simulated forest fire. For. Ecol. Manag. 1992, 53, 329–343. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef]
- Durán, J.; Rodríguez, A.; Fernández-Palacios, J.M.; Gallardo, A. Changes in soil N and P availability in a Pinus canariensis fire chronosequence. For. Ecol. Manag. 2008, 256, 384–387. [Google Scholar] [CrossRef]
- Durán, J.; Rodríguez, A.; Fernández-Palacios, J.M.; Gallardo, A. Changes in net N mineralization rates and soil N and P pools in a pine forest wildfire chronosequence. Biol. Fertil. Soils 2009, 45, 781–788. [Google Scholar] [CrossRef]
- Durán, J.; Rodríguez, A.; Fernández-Palacios, J.M.; Gallardo, A. Changes in leaf nutrient traits in a wildfire chronosequence. Plant Soil 2010, 331, 69–77. [Google Scholar] [CrossRef]
- Durán, J.; Rodríguez, A.; Méndez, J.; Morales, G.; Fernández-Palacios, J.M.; Gallardo, A. Wildfires decrease the local-scale ecosystem spatial variability of Pinus canariensis forests during the first two decades post fire. Int. J. Wildland Fire 2019, 28, 288–294. [Google Scholar] [CrossRef] [Green Version]
- Turco, M.; von Hardenberg, J.; AghaKouchak, A.; Llasat, M.C.; Provenzale, A.; Trigo, R.M. On the key role of droughts in the dynamics of summer fires in Mediterranean Europe. Sci. Rep. 2017, 7, 1–10. [Google Scholar]
- Stephens, S.L.; Agee, J.K.; Fulé, P.Z.; North, M.P.; Romme, W.H.; Swetnam, T.W.; Turner, M.G. Managing forests and fire in changing climates. Science 2013, 342, 41–42. [Google Scholar] [CrossRef] [PubMed]
- Enright, N.J.; Fontaine, J.B.; Bowman, D.M.; Bradstock, R.A.; Williams, R.J. Interval squeeze: Altered fire regimes and demographic responses interact to threaten woody species persistence as climate changes. Front. Ecol. Environ. 2015, 13, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Abatzoglou, J.T.; Williams, A.P. Impact of anthropogenic climate change on wildfire across western US forests. Proc. Natl. Acad. Sci. USA 2016, 113, 11770–11775. [Google Scholar] [CrossRef] [Green Version]
- Turco, M.; Rosa-Cánovas, J.J.; Bedia, J.; Jerez, S.; Montávez, J.P.; Llasat, M.C.; Provenzale, A. Exacerbated fires in Mediterranean Europe due to anthropogenic warming projected with non-stationary climate-fire models. Nat. Commun. 2018, 9, 3821. [Google Scholar] [CrossRef] [PubMed]
- Harter, D.; Irl, S.; Seo, B.; Steinbauer, M.; Gillespie, R.G.; Triantis, K.A.; Fernandez-Palacios, J.M.; Beierkuhnlein, C. Impacts of global climate change on the floras of oceanic islands—Projections, implications and current knowledge. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 160–183. [Google Scholar] [CrossRef]
- Luis, V.C.; Jiménez, M.S.; Morales, D.; Kucera, J.; Wieser, G. Canopy transpiration of a Canary Islands pine forest. Agric. For. Meteorol. 2005, 135, 117–123. [Google Scholar] [CrossRef]
- Wieser, G.; Brito, P.; Lorenzo, J.R.; González-Rodríguez, Á.M.; Morales, D.; Jiménez, M.S. Canary Island pine (Pinus canariensis), an evergreen species in a semiarid treeline. Prog. Bot. 2016, 77, 415–435. [Google Scholar]
- Climent, J.; López, R.; González, S.; Gil, L. El pino canario (Pinus canariensis), una especie singular. Ecosistemas 2007, 16, 80–89. [Google Scholar]
- Whittaker, R.J.; Fernández-Palacios, J.M. Island Biogeography: Ecology, Evolution, and Conservation, 2nd ed.; Oxford University Press: Oxford, UK; New York, NY, USA, 2007; p. 416. [Google Scholar]
- Beierkuhnlein, C.; Walentowitz, A.; Welss, W. FloCan—A revised checklist for the flora of the Canary Islands. Diversity 2021, 13, 480. [Google Scholar] [CrossRef]
- Irl, S.D.; Steinbauer, M.J.; Messinger, J.; Blume-Werry, G.; Palomares-Martínez, Á.; Beierkuhnlein, C.; Jentsch, A. Burned and devoured—Introduced herbivores, fire, and the endemic flora of the high-elevation ecosystem on La Palma, Canary Islands. Arct. Antarc. Alp. Res. 2014, 46, 859–869. [Google Scholar] [CrossRef] [Green Version]
- Arévalo, J.R.; Fernández-Palacios, J.M.; Jiménez, M.J.; Gil, P. The effect of fire intensity on the understory species composition of two Pinus canariensis reforested stands in Tenerife (Canary Islands). For. Ecol. Manag. 2001, 148, 21–29. [Google Scholar] [CrossRef]
- Gómez-González, S.; Torres-Díaz, C.; Valencia, G.; Torres-Morales, P.; Cavieres, L.A.; Pausas, J.G. Anthropogenic fires increase alien and native annual species in the Chilean coastal matorral. Divers. Distrib. 2011, 17, 58–67. [Google Scholar] [CrossRef]
- Carracedo, H.C.; Pérez Torrado, F.J.; Ancochea, E.; Meco, J.; Hernán, F.; Cubas, C.R.; Casillas, R.; Rodriguez, E.; Ahijado, A. Cenozoic volcanism II: The Canary Island. In The Geology of Spain, 1st ed.; Gibbon, W., Moreno, T., Eds.; The Geological Society: London, UK, 2002; pp. 439–472. [Google Scholar]
- Garzón-Machado, V.; Otto, R.; del Arco Aguilar, M.J. Bioclimatic and vegetation mapping of a topographically complex oceanic island applying different interpolation techniques. Int. J. Biometeorol. 2014, 58, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Weigel, R.; Irl, S.D.; Treydte, K.; Beierkuhnlein, C.; Berels, J.; Field, R.; Miranda, J.C.; Steinbauer, A.; Steinbauer, M.J.; Jentsch, A. A novel dendroecological method finds a non-linear relationship between elevation and seasonal growth continuity on an island with trade wind-influenced water availability. AoB Plants 2018, 10, ply070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gascon, F.; Cadau, E.; Colin, O.; Hoersch, B.; Isola, C.; Fernández, B.L.; Martimort, P. Copernicus Sentinel-2 Mission: Products, Algorithms and Cal/Val. Earth Obs. Syst. XIX 2014, 9218, 92181E. [Google Scholar]
- Drusch, M.; Del Bello, U.; Carlier, S.; Colin, O.; Fernandez, V.; Gascon, F.; Hoersch, B.; Isola, C.; Laberinti, P.; Martimort, P.; et al. Sentinel-2: ESA’s optical high-resolution mission for GMES operational services. Remote Sens. Environ. 2012, 120, 25–36. [Google Scholar] [CrossRef]
- Navarro, G.; Caballero, I.; Silva, G.; Parra, P.C.; Vázquez, Á.; Caldeira, R. Evaluation of forest fire on Madeira Island using Sentinel-2A MSI imagery. Int. J. Appl. Earth Obs. Geoinf. 2017, 58, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Pádua, L.; Guimarães, N.; Adão, T.; Sousa, A.; Peres, E.; Sousa, J.J. Effectiveness of sentinel-2 in multi-temporal post-fire monitoring when compared with UAV imagery. ISPRS Int. J. Geoinf. 2020, 9, 225. [Google Scholar] [CrossRef] [Green Version]
- Quintano, C.; Fernández-Manso, A.; Fernández-Manso, O. Combination of landsat and sentinel-2 MSI data for initial assessing of burn severity. Int. J. Appl. Earth Obs. Geoinf. 2018, 64, 221–225. [Google Scholar] [CrossRef]
- Chen, X.; Vogelmann, J.E.; Rollins, M.; Ohlen, D.; Key, C.H.; Yang, L.; Hunga, C.; Shi, H. Detecting post-fire burn severity and vegetation recovery using multitemporal remote sensing spectral indices and field-collected composite burn index data in a ponderosa pine forest. Int. J. Remote Sens. 2011, 32, 7905–7927. [Google Scholar] [CrossRef]
- Justice, C.O.; Giglio, L.; Korontzi, S.; Owens, J.; Morisette, J.T.; Roy, D.; Descloitres, J.; Alleaume, S.; Petitcolin, F.; Kaufman, Y. The MODIS fire products. Remote Sens. Environ. 2002, 83, 244–262. [Google Scholar] [CrossRef]
- Hernández-Leal, P.A.; González-Calvo, A.; Arbelo, M.; Barreto, A.; Alonso-Benito, A. Synergy of GIS and remote sensing data in forest fire danger modeling. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2008, 1, 240–247. [Google Scholar] [CrossRef]
- Levin, N.; Heimowitz, A. Mapping spatial and temporal patterns of Mediterranean wildfires from MODIS. Remote Sens. Environ. 2012, 126, 12–26. [Google Scholar] [CrossRef]
- Frantz, D. FORCE—Landsat+ Sentinel-2 analysis ready data and beyond. Remote Sens. 2019, 11, 1124. [Google Scholar] [CrossRef] [Green Version]
- Bivand, R.; Keitt, T.; Rowlingson, B. Rgdal: Bindings for the ’Geospatial’ Data Abstraction Library, R package version 1.5-23; Available online: http://cran.r-project.org/%0Apackage=rgdal (accessed on 22 November 2021).
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling, R package version 3.4-5; Available online: https://cran.r-project.org/web/packages/raster/ (accessed on 22 November 2021).
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. Available online: https://www.r-project.org/ (accessed on 22 November 2021).
- Amici, V.; Rocchini, D.; Filibeck, G.; Bacaro, G.; Santi, E.; Geri, F.; Landi, S.; Scoppola, A.; Chiarucci, A. Landscape structure effects on forest plant diversity at local scale: Exploring the role of spatial extent. Ecol. Complex. 2015, 21, 44–52. [Google Scholar] [CrossRef]
- Chuvieco, E.; Martin, M.P.; Palacios, A. Assessment of different spectral indices in the red-near-infrared spectral domain for burned land discrimination. Int. J. Remote Sens. 2002, 23, 5103–5110. [Google Scholar] [CrossRef]
- Key, C.H.; Benson, N.; Ohlen, D.; Howard, S.; McKinley, R.; Zhu, Z. The normalized burn ratio and relationships to burn severity: Ecology, remote sensing and implementation. In Proceedings of the Ninth Forest Service Remote Sensing Applications Conference, San Diego, CA, USA, 8–12 April 2002; Greer, J.D., Ed.; American Society for Photogrammetry and Remote Sensing: Bethesda, MD, USA, 2002. [Google Scholar]
- Tree Cover Density 201—Copernicus Land Monitoring Service. Available online: https://land.copernicus.eu/pan-european/high-resolution-layers/forests/tree-cover-density/status-maps/2015 (accessed on 24 November 2021).
- Jenness, J.S. Calculating landscape surface area from digital elevation models. Wildl. Soc. Bull. 2004, 32, 829–839. [Google Scholar] [CrossRef]
- Riley, S.J.; DeGloria, S.D.; Elliot, R. A terrain ruggedness index that quantifies topographic heterogeneity. Intermt. J. Sci. 1999, 5, 23–27. [Google Scholar]
- Bitterlich, W. Die Winkelzählprobe. Forstwiss. Cent. 1952, 71, 215–225. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.84). Available online: https://cran.r-project.org/web/packages/corrplot/ (accessed on 22 November 2021).
- Signorell, A.; Aho, K.; Alfons, A.; Anderegg, N.; Aragon, T.; Arppe, A. DescTools: Tools for Descriptive Statistics, R package version 0.99; Available online: https://cran.r-project.org/web/packages/DescTools/ (accessed on 22 November 2021).
- Bivand, R.; Altman, M.; Anselin, L.; Assunção, R.; Berke, O.; Bernat, A.; Blanchet, G.; Blankmeyer, E.; Carvalho, M.; Christensen, B.; et al. Spdep: Spatial Dependence: Weighting Schemes, Statistics and Models. Available online: https://cran.r-project.org/web/packages/spdep/ (accessed on 22 November 2021).
- Bivand, R.; Piras, G. Spatialreg: Spatial Regression Analysis. Available online: https://cran.r-project.org/web/packages/spatialreg/ (accessed on 22 November 2021).
- Kissling, W.D.; Carl, G. Spatial autocorrelation and the selection of simultaneous autoregressive models. Glob. Ecol. Biogeogr. 2008, 17, 59–71. [Google Scholar] [CrossRef]
- Kreft, H.; Jetz, W. Global patterns and determinants of vascular plant diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 5925–5930. [Google Scholar] [CrossRef] [Green Version]
- Tognelli, M.F.; Kelt, D.A. Analysis of determinants of mammalian species richness in South America using spatial autoregressive models. Ecography 2004, 27, 427–436. [Google Scholar] [CrossRef]
- Irl, S.D.; Harter, D.E.; Steinbauer, M.J.; Gallego Puyol, D.; Fernández-Palacios, J.M.; Jentsch, A.; Beierkuhnlein, C. Climate vs. topography–spatial patterns of plant species diversity and endemism on a high-elevation island. J. Ecol. 2015, 103, 1621–1633. [Google Scholar] [CrossRef]
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: Testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Anderson, M.J.; Crist, T.O.; Chase, J.M.; Vellend, M.; Inouye, B.D.; Freestone, A.L.; Sanders, N.J.; Cornell, H.V.; Comita, L.S.; Davies, K.F.; et al. Navigating the multiple meanings of β diversity: A roadmap for the practicing ecologist. Ecol. Lett. 2011, 14, 19–28. [Google Scholar] [CrossRef]
- Odum, E.P. Bird populations of the Highlands (North Carolina) Plateau in relation to plant succession and avian invasion. Ecology 1950, 31, 587–605. [Google Scholar] [CrossRef]
- Legendre, P.; De Cáceres, M. Βeta-diversity as the variance of community data: Dissimilarity coefficients and partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, R package version 2.5-6; Available online: https://cran.r-project.org/web/packages/vegan/ (accessed on 22 November 2021).
- Wang, I. topoDistance: Calculating Topographic Paths and Distances, R package version 1.0.1; Available online: https://cran.r-project.org/web/packages/topoDistance/ (accessed on 22 November 2021).
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Dray, S.; Bauman, D.; Blanchet, G.; Borcard, D.; Clappe, S.; Guenard, G.; Jombart, T.; Larocque, G.; Legendre, P.; Madi, N.; et al. adespatial: Multivariate Multiscale Spatial Analysis, R package version 0.3-8; Available online: https://cran.r-project.org/web/packages/adespatial/ (accessed on 22 November 2021).
- Halpern, C.B. Early successional pathways and the resistance and resilience of forest communities. Ecology 1988, 69, 1703–1715. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Irl, S.; González, J.M.; Breiner, F.; Hernández Hernández, R.M.; Hopfenmüller, S.; Kidane, Y.; Jentsch, A.; Beierkuhnlein, C. Plant invasion and speciation along elevational gradients on the oceanic island La Palma, Canary Islands. Ecol. Evol. 2017, 7, 771–779. [Google Scholar] [CrossRef] [Green Version]
- Garzón-Machado, V.; del Arco Aguilar, M.J.; González, F.V.; Pérez-de-Paz, P.L. Fire as a threatening factor for endemic plants of the Canary Islands. Biodivers. Conserv. 2012, 21, 2621–2632. [Google Scholar] [CrossRef]
- Sánchez Sánchez, Y.; Martínez-Graña, A.; Santos Francés, F.; Mateos Picado, M. Mapping wildfire ignition probability using sentinel 2 and LiDAR (Jerte Valley, Cáceres, Spain). Sensors 2018, 18, 826. [Google Scholar] [CrossRef] [Green Version]
- Teodoro, A.; Amaral, A. A statistical and spatial analysis of Portuguese forest fires in summer 2016 considering Landsat 8 and Sentinel 2A data. Environments 2019, 6, 36. [Google Scholar] [CrossRef] [Green Version]
- Filipponi, F. Exploitation of sentinel-2 time series to map burned areas at the national level: A case study on the 2017 Italy wildfires. Remote Sens. 2019, 11, 622. [Google Scholar] [CrossRef] [Green Version]
- Evangelides, C.; Nobajas, A. Red-edge normalised difference vegetation index (NDVI705) from sentinel-2 imagery to assess post-fire regeneration. Remote Sens. Appl. Soc. Environ. 2020, 17, 100283. [Google Scholar] [CrossRef]
- Richter, C.; Rejmánek, M.; Miller, J.E.; Welch, K.R.; Weeks, J.; Safford, H. The species diversity × fire severity relationship is hump-shaped in semiarid yellow pine and mixed conifer forests. Ecosphere 2019, 10, e02882. [Google Scholar] [CrossRef]
- Miller, J.E.; Safford, H.D. Are plant community responses to wildfire contingent upon historical disturbance regimes? Glob. Ecol. Biogeogr. 2020, 29, 1621–1633. [Google Scholar] [CrossRef]
- Hoffmann, S.; Schmitt, T.M.; Chiarucci, A.; Irl, S.D.; Rocchini, D.; Vetaas, O.R.; Tanase, M.A.; Mermoz, S.; Bouvet, A.; Beierkuhnlein, C. Remote sensing of β-diversity: Evidence from plant communities in a semi-natural system. Appl. Veg. Sci. 2019, 22, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Tuomisto, H. A diversity of beta diversities: Straightening up a concept gone awry. Part 1. Defining beta diversity as a function of alpha and gamma diversity. Ecography 2010, 33, 2–22. [Google Scholar] [CrossRef]
- Tuomisto, H. A diversity of beta diversities: Straightening up a concept gone awry. Part 2. Quantifying beta diversity and related phenomena. Ecography 2010, 33, 23–45. [Google Scholar] [CrossRef]
- Jurasinski, G.; Retzer, V.; Beierkuhnlein, C. Inventory, differentiation, and proportional diversity—a consistent terminology for quantifying biodiversity. Oecologia 2009, 159, 15–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, J.A.; Chase, J.M.; Crandall, R.M.; Jiménez, I. Disturbance alters beta-diversity but not the relative importance of community assembly mechanisms. J. Ecol. 2015, 103, 1291–1299. [Google Scholar] [CrossRef] [Green Version]
- Sandral, G.; Remizowa, M.V.; Sokoloff, D.D. A taxonomic survey of Lotus section Pedrosia (Leguminosae, Loteae). Wulfenia 2006, 13, 97–192. [Google Scholar]
- Irl, S.D.; Obermeier, A.; Beierkuhnlein, C.; Steinbauer, M.J. Climate controls plant life-form patterns on a high-elevation oceanic island. J. Biogeogr. 2020, 47, 2261–2273. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Variable | Unit | Source |
|---|---|---|---|
| Vegetation | Species cover | % | Field measurement |
| Herb/shrub cover estimates | % | Field measurement | |
| Canopy cover | % | Field measurement | |
| Litter depth | cm | Field measurement | |
| Basal area | m2/ha | Field measurement | |
| Diameter at breast height (DBH) | cm | Field measurement | |
| Topography | Rock cover | % | Field measurement |
| Inclination | % | Field measurement | |
| Elevation | m | Cabildo Insular DEM (2m resolution) | |
| Aspect | Northness and Eastness | Field measurement, converted with the cosine and sine function |
| Species Richness~ | Estimate | Std. Error | z value | Pr (>|z|) | Significance |
|---|---|---|---|---|---|
| (Intercept) | 11.7 | 2.37 | 4.92 | <0.001 | *** |
| Elevation | −0.012 | 0.00363 | −3.3 | <0.001 | *** |
| Elevation2 | 3.57 × 10−6 | 1.37 × 10−6 | 2.61 | 0.009 | ** |
| R2 Nagelkerke | 0.81 | AIC | 464.68 | ||
| Species richness~ | Estimate | Std. Error | z value | Pr (>|z|) | |
| (Intercept) | 2.72 | 0.485 | 5.62 | <0.001 | *** |
| ΔNDVI | 5.05 | 2.55 | 1.98 | 0.0476 | * |
| ΔNDVI 2 | 8.9 | 3.23 | 2.75 | 0.0059 | ** |
| R2 Nagelkerke | 0.33 | AIC | 565.9 | ||
| Species richness~ | Estimate | Std. Error | z value | Pr (>|z|) | |
| (Intercept) Tree cover (RS) | 1.27 0.018 | 0.107 0.00179 | 11.9 10.1 | <0.001 <0.001 | *** |
| *** | |||||
| R2 Nagelkerke | 0.70 | AIC | 565.9 | ||
| Species richness~ | Estimate | Std. Error | z value | Pr (>|z|) | |
| (Intercept) | 7.63 | 2.99 | 2.55 | 0.0108 | * |
| Elevation | −0.0102 | 0.00457 | −2.23 | 0.0255 | * |
| Elevation2 | 3.02 × 10−6 | 1.71 × 10−6 | 1.77 | 0.0763 | |
| Tree cover (field) | 0.0101 | 0.0135 | 0.746 | 0.456 | |
| Tree cover (field)2 | −0.000382 | 0.000201 | −1.9 | 0.0578 | |
| Northness | 0.22 | 0.0844 | 2.61 | 0.00898 | ** |
| Eastness | −0.166 | 0.108 | −1.54 | 0.123 | |
| DBH mean | 0.0209 | 0.00677 | 3.09 | 0.00202 | ** |
| Litter depth mean | 0.126 | 0.047 | 2.66 | 0.00774 | ** |
| ΔNDVI | −6.2 | 2.77 | −2.24 | 0.0252 | * |
| ΔNDVI2 | −6.07 | 3.65 | −1.66 | 0.0963 | |
| Tree cover (RS) | 0.00619 | 0.00226 | 2.74 | 0.0062 | ** |
| R2 Nagelkerke | 0.935 | AIC | 400 |
| Bray–Curtis Dissimilarity | ΔΔNDVI | Topographic Distance | Environmental Distance | |
|---|---|---|---|---|
| Bray–Curtis Dissimilarity | - | 0.02 | 0.31 * | 0.35 * |
| ΔΔNDVI | 0.02 | - | −0.12 | 0.17 * |
| Topographic Distance | 0.31 * | −0.01 | - | 0.18 * |
| Environmental Distance | 0.35 * | 0.17 * | 0.18 * | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weiser, F.; Sauer, A.; Gettueva, D.; Field, R.; Irl, S.D.H.; Vetaas, O.; Chiarucci, A.; Hoffmann, S.; Fernández-Palacios, J.M.; Otto, R.; et al. Impacts of Forest Fire on Understory Species Diversity in Canary Pine Ecosystems on the Island of La Palma. Forests 2021, 12, 1638. https://doi.org/10.3390/f12121638
Weiser F, Sauer A, Gettueva D, Field R, Irl SDH, Vetaas O, Chiarucci A, Hoffmann S, Fernández-Palacios JM, Otto R, et al. Impacts of Forest Fire on Understory Species Diversity in Canary Pine Ecosystems on the Island of La Palma. Forests. 2021; 12(12):1638. https://doi.org/10.3390/f12121638
Chicago/Turabian StyleWeiser, Frank, Anna Sauer, Daria Gettueva, Richard Field, Severin D. H. Irl, Ole Vetaas, Alessandro Chiarucci, Samuel Hoffmann, José María Fernández-Palacios, Rüdiger Otto, and et al. 2021. "Impacts of Forest Fire on Understory Species Diversity in Canary Pine Ecosystems on the Island of La Palma" Forests 12, no. 12: 1638. https://doi.org/10.3390/f12121638
APA StyleWeiser, F., Sauer, A., Gettueva, D., Field, R., Irl, S. D. H., Vetaas, O., Chiarucci, A., Hoffmann, S., Fernández-Palacios, J. M., Otto, R., Jentsch, A., Provenzale, A., & Beierkuhnlein, C. (2021). Impacts of Forest Fire on Understory Species Diversity in Canary Pine Ecosystems on the Island of La Palma. Forests, 12(12), 1638. https://doi.org/10.3390/f12121638