
Evaluation of LandscapeDNDC Model Predictions of CO2 and N2O Fluxes from an Oak Forest in SE England


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. LandscapeDNDC Model Framework
2.3. Model Evaluation
2.4. Soil Gas Flux and Environmental Measurements
2.5. CO2 Flux Data from Eddy Covariance
3. Results
3.1. Data–Model Comparisons and Model Adjustment
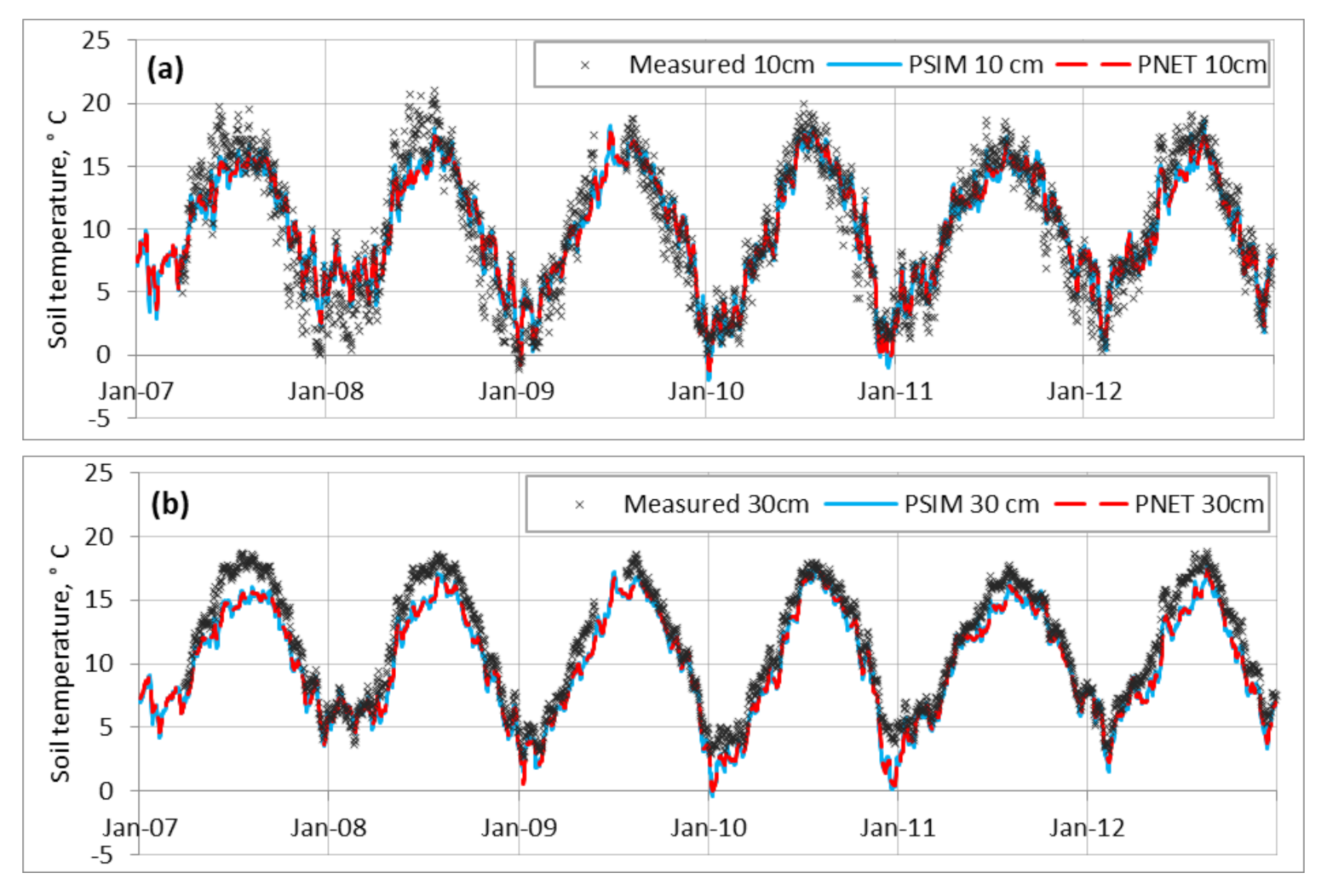
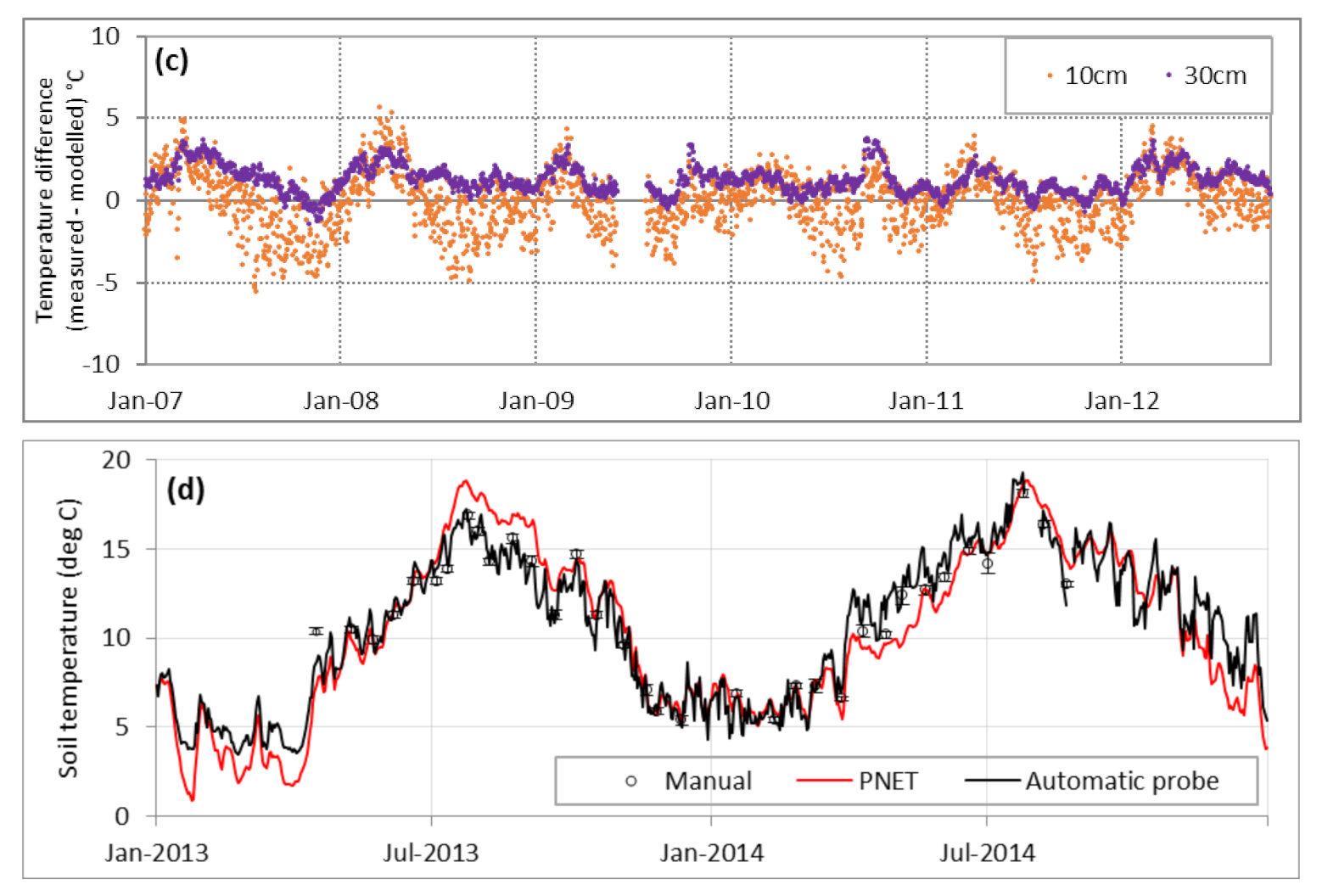
3.1.1. Environmental Conditions
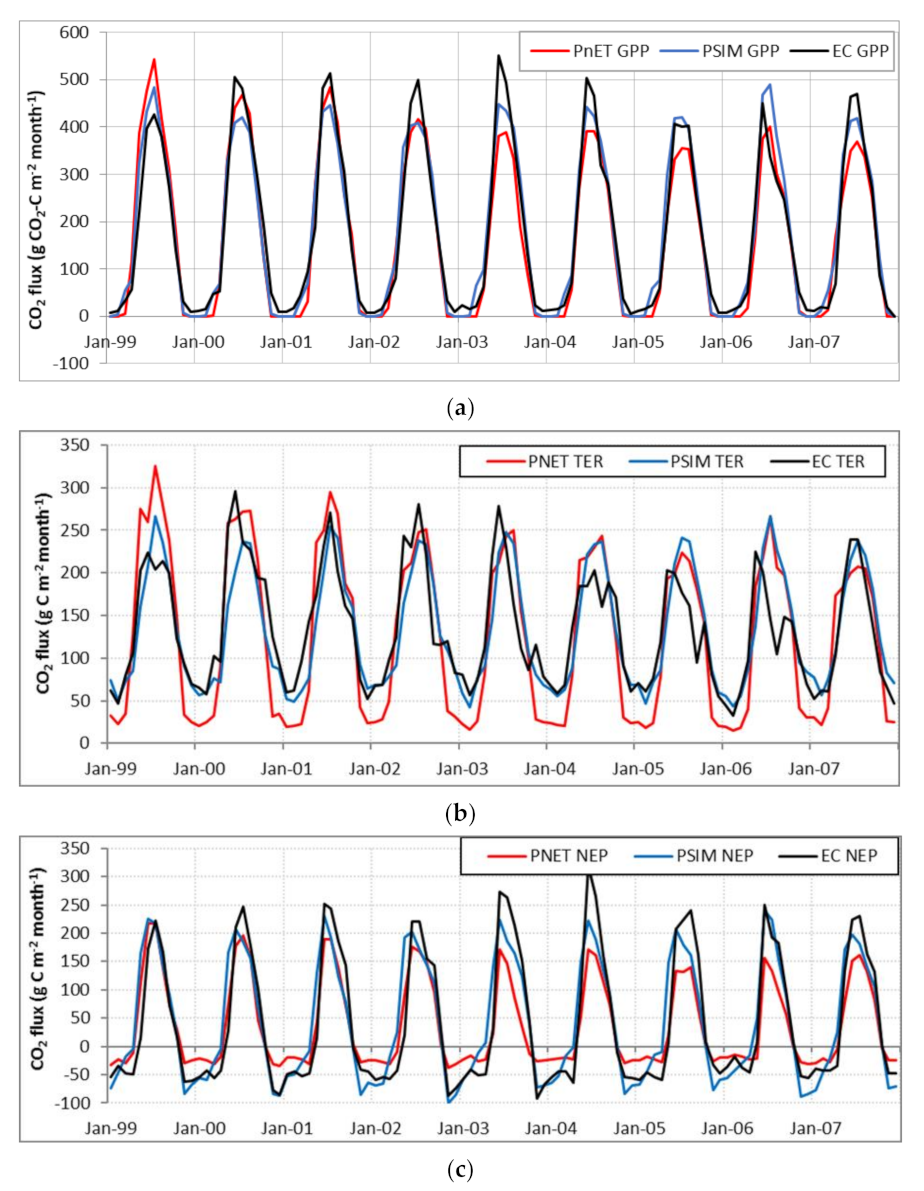
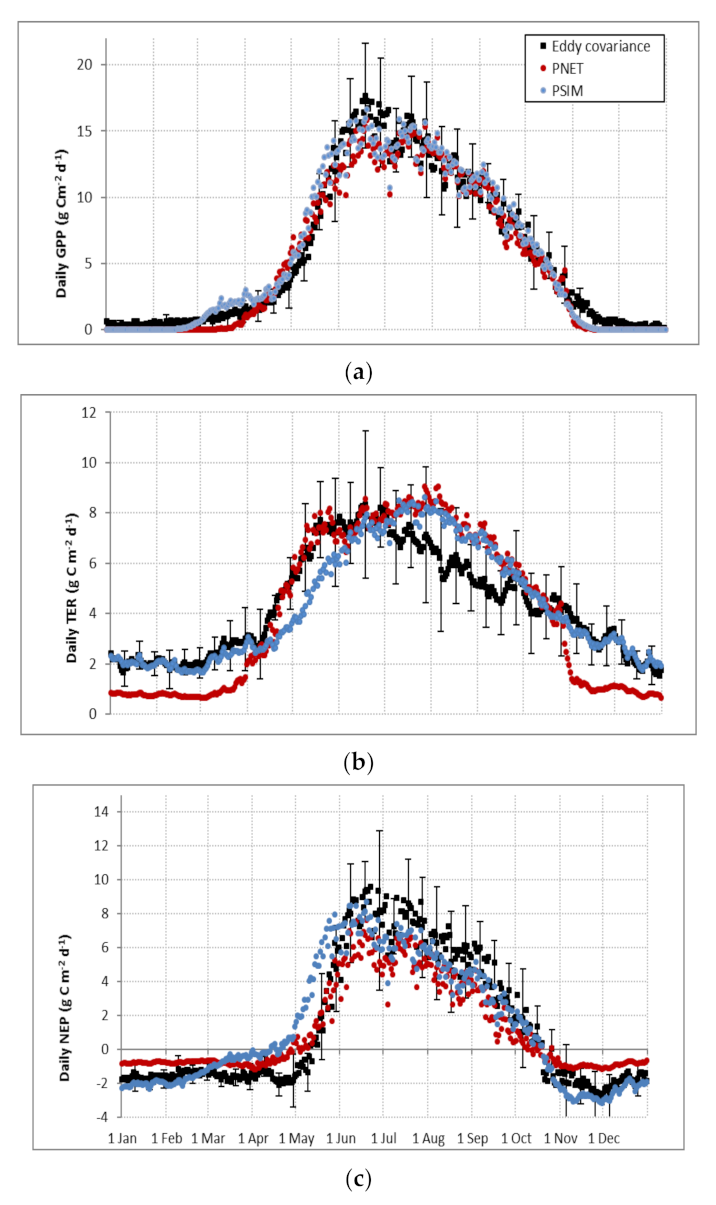
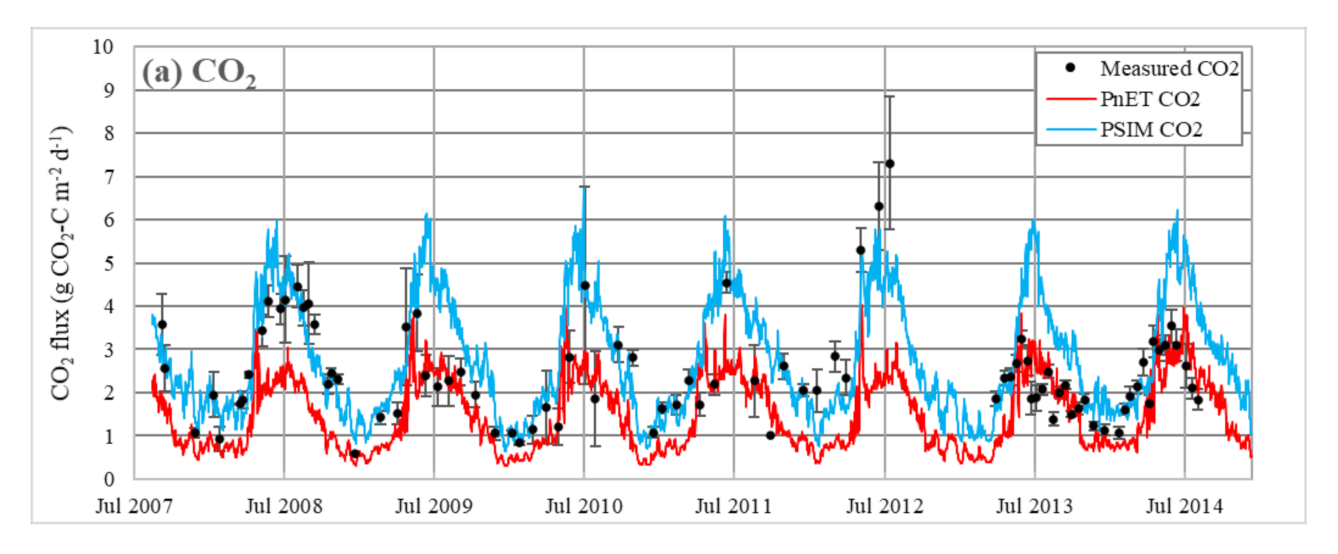
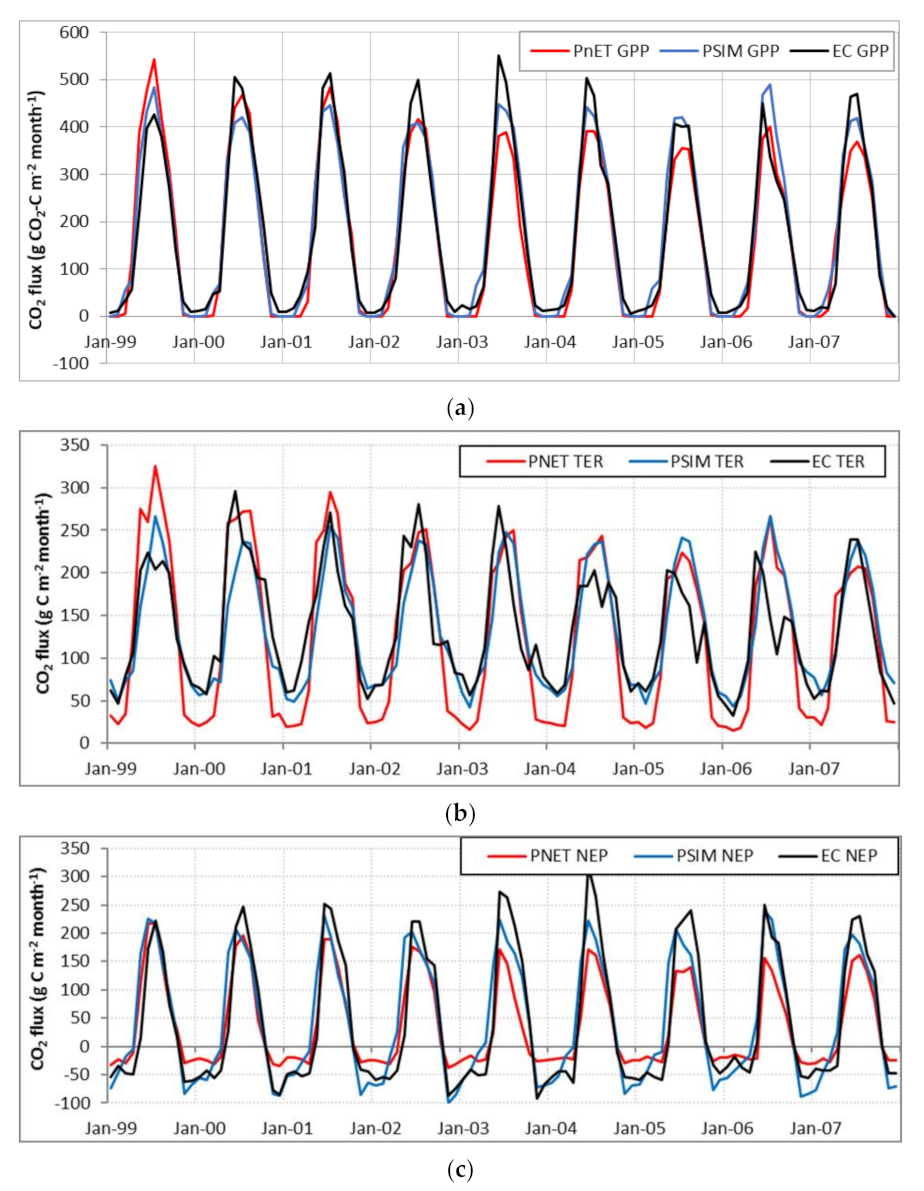
3.1.2. Ecosystem CO2 Fluxes
3.1.3. Soil CO2 Effluxes
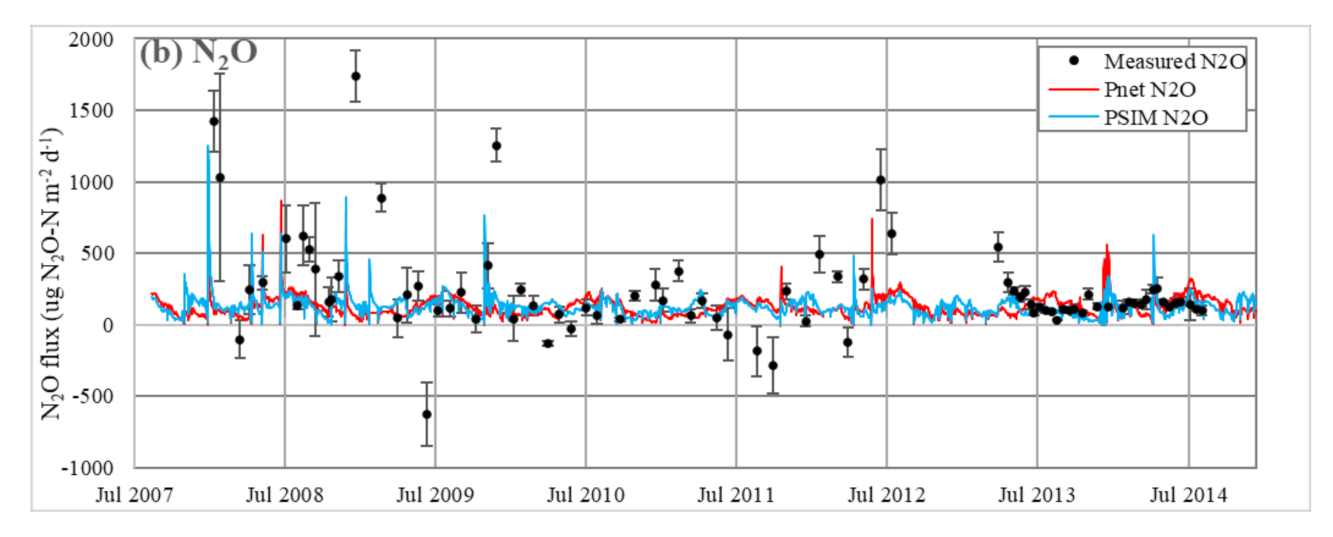
3.1.4. Soil Gaseous N Fluxes
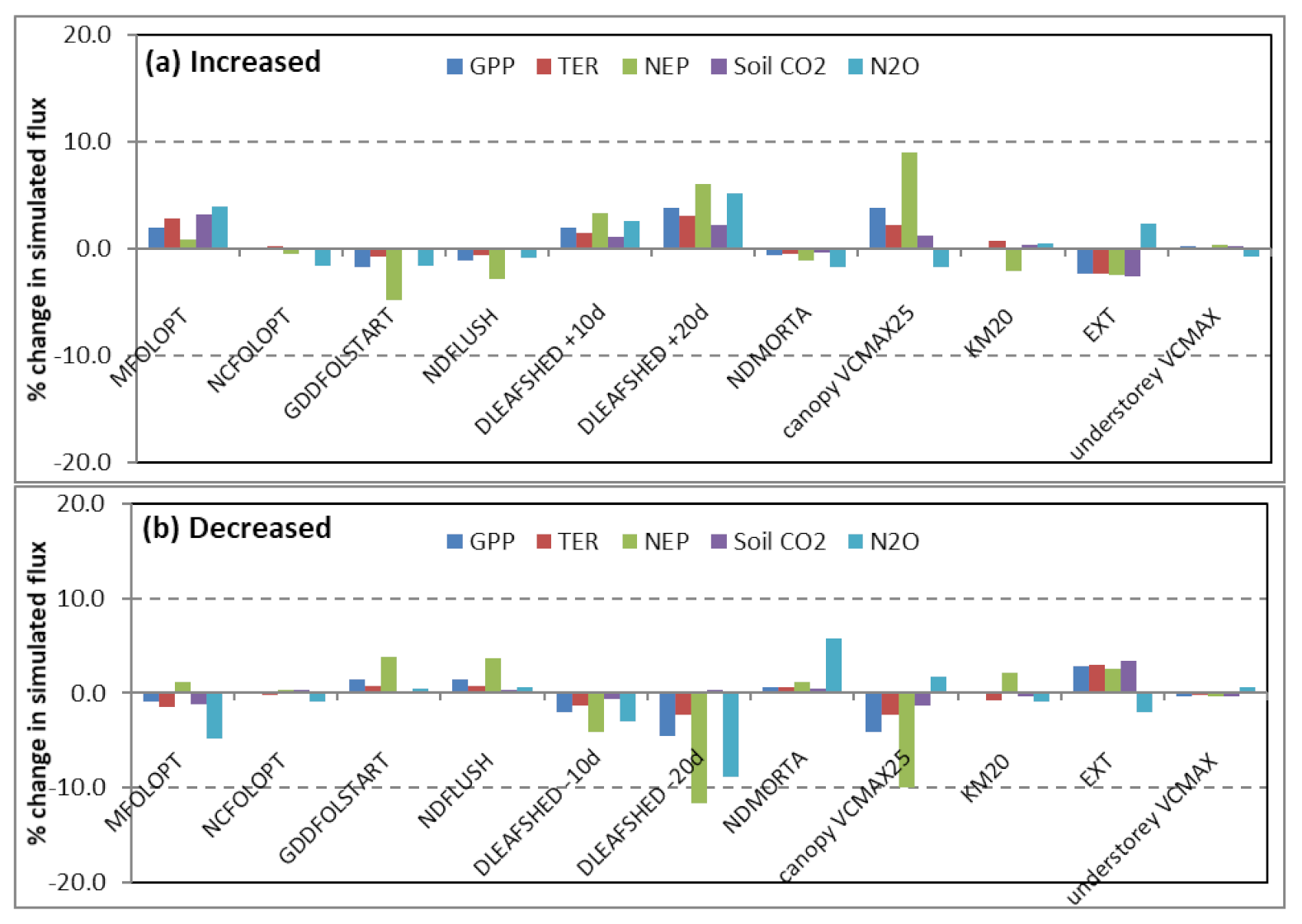
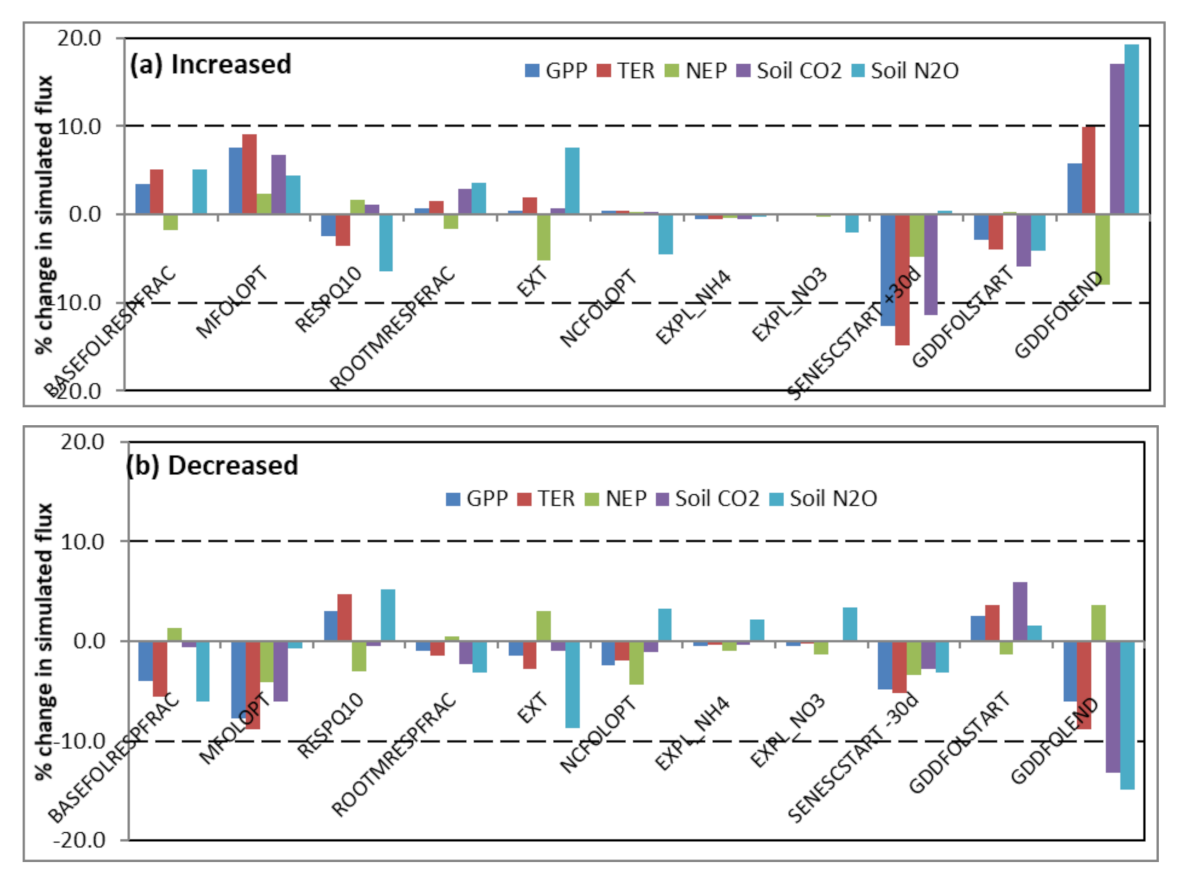
3.2. Model Sensitivity Analysis
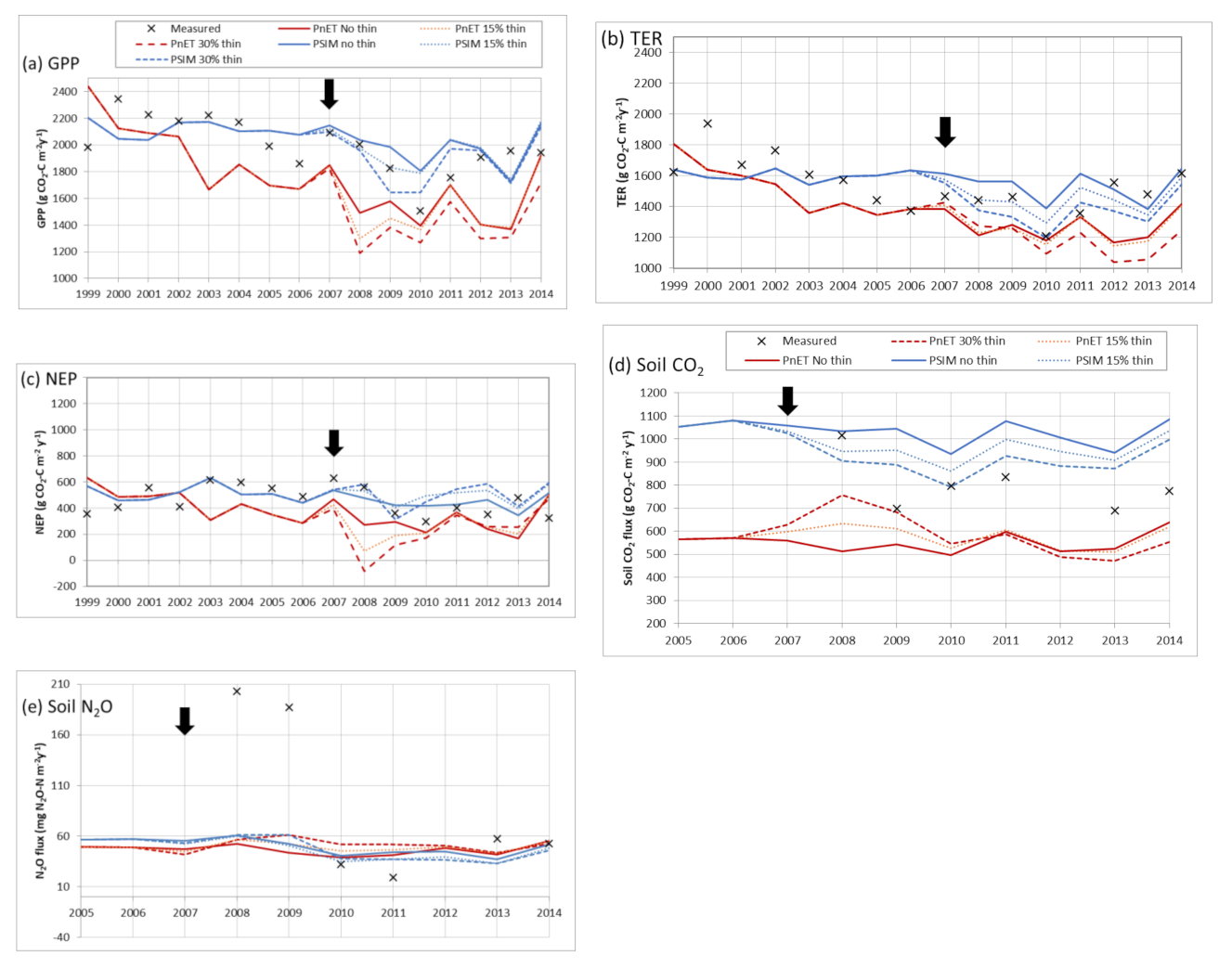
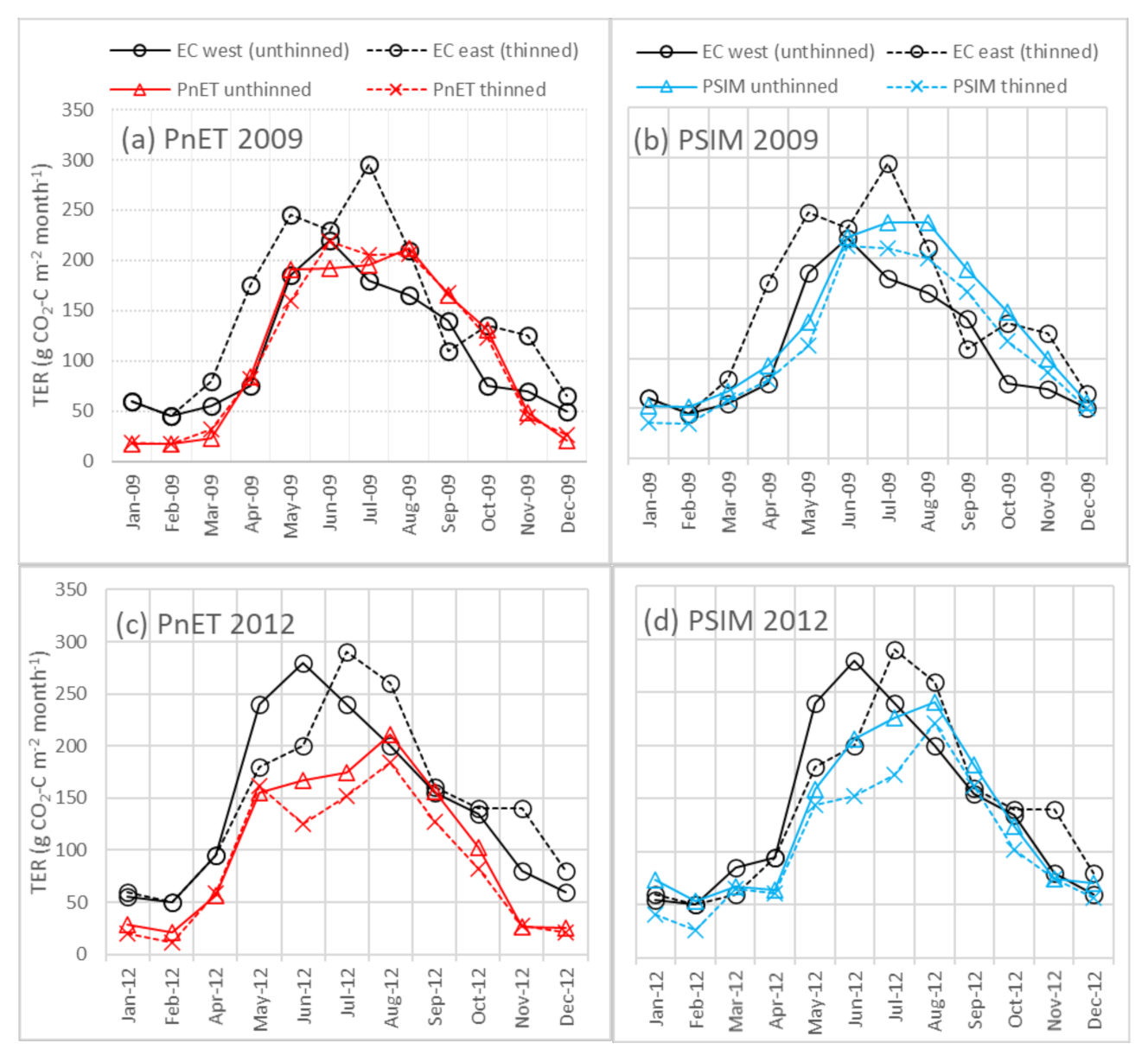
3.3. Thinning
4. Discussion
4.1. Simulation of Environmental Conditions
4.2. Vegetation Species Parameters and Simulation of CO2 Fluxes
4.3. Simulating Soil Gas Fluxes and Measurement Uncertainty
4.4. Simulating Thinning/Response to Management Change
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ciais, P.; Sabine, C.; Bala, G.; Bopp, L.; Brovkin, V.; Canadell, J.; Chhabra, A.; DeFries, R.; Galloway, J.; Heimann, M.; et al. 2013: Carbon and Other Biogeochemical Cycles. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M.M.B., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [Green Version]
- Finzi, A.C.; Giasson, M.A.; Barker Plotkin, A.A.; Aber, J.D.; Boose, E.R.; Davidson, A.E.; Dietze, M.C.; Ellison, A.M.; Frey, S.D.; Goldman, E.; et al. Carbon budget of the Harvard Forest Long-Term Ecological Research site: Pattern, process, and response to global change. Ecol. Monogr. 2020, 90, e01423. [Google Scholar] [CrossRef]
- Thurner, M.; Beer, C.; Santoro, M.; Carvalhais, N.; Wutzler, T.; Schepaschenko, D.; Shvidenko, A.; Kompter, E.; Ahrens, B.; Levick, S.R. Carbon stock and density of northern boreal and temperate forests. Global Ecol. Biogeogr. 2014, 23, 297–310. [Google Scholar] [CrossRef]
- Blagodatsky, S.; Yevdokimov, I.; Larionova, A.; Richter, J. Microbial growth in soil and nitrogen turnover: Model calibration with laboratory data. Soil Biol. Biochem. 1998, 30, 1757–1764. [Google Scholar] [CrossRef]
- Ingwersen, J.; Butterbach-Bahl, K.; Gasche, R.; Papen, H.; Richter, O. Barometric process separation: New method for quantifying nitrification, denitrification, and nitrous oxide sources in soils. Soil Sci. Soc. Am. J. 1999, 63, 117–128. [Google Scholar] [CrossRef]
- Schindlbacher, A.; Zechmeister-Boltenstern, S.; Butterbach-Bahl, K. Effects of soil moisture and temperature on NO, NO2, and N2O emissions from European forest soils. J. Geophys. Res. Atmos. 2004, 109, D17302. [Google Scholar] [CrossRef]
- Pilegaard, K.; Skiba, U.; Ambus, P.; Beier, C.; Brüggemann, N.; Butterbach-Bahl, K.; Dick, J.; Dorsey, J.; Duyzer, J.; Gallagher, M. Factors controlling regional differences in forest soil emission of nitrogen oxides (NO and N2O). Biogeosciences 2006, 3, 651–661. [Google Scholar] [CrossRef] [Green Version]
- Luo, G.; Brüggemann, N.; Wolf, B.; Gasche, R.; Grote, R.; Butterbach-Bahl, K. Decadal variability of soil CO2, NO, N2O, and CH4 fluxes at the Höglwald Forest, Germany. Biogeosciences 2012, 9, 1741–1763. [Google Scholar] [CrossRef] [Green Version]
- Haas, E.; Klatt, S.; Fröhlich, A.; Kraft, P.; Werner, C.; Kiese, R.; Grote, R.; Breuer, L.; Butterbach-Bahl, K. LandscapeDNDC: A process model for simulation of biosphere-atmosphere-hydrosphere exchange processes at site and regional scale. Landsc. Ecol. 2013, 28, 615–636. [Google Scholar] [CrossRef]
- Cameron, D.R.; Van Oijen, M.; Werner, C.; Butterbach-Bahl, K.; Grote, R.; Haas, E.; Heuvelink, G.B.M.; Kiese, R.; Kros, J.; Kuhnert, M.; et al. Environmental change impacts on the C- and N-cycle of European forests: A model comparison study. Biogeosciences 2013, 10, 1751–1773. [Google Scholar] [CrossRef] [Green Version]
- Aber, J.D.; Federer, C.A. A generalized, lumped-parameter model of photosynthesis, evapotranspiration and net primary production in temperate and boreal forest ecosystems. Oecologia 1992, 92, 463–474. [Google Scholar] [CrossRef]
- Aber, J.D.; Ollinger, S.V.; Federer, C.A.; Reich, P.B.; Goulden, M.L.; Kicklighter, D.W.; Melillo, J.; Lathrop Jr, R.G. Predicting the effects of climate change on water yield and forest production in the northeastern United States. Clim. Res. 1995, 5, 207–222. [Google Scholar] [CrossRef]
- Grote, R. Sensitivity of volatile monoterpene emission to changes in canopy structure: A model-based exercise with a process-based emission model. New Phytol. 2007, 173, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Grote, R.; Kiese, R.; Grünwald, T.; Ourcival, J.; Granier, A. Modelling forest carbon balances considering tree mortality and removal. Agric. For. Meteorol. 2011, 151, 179–190. [Google Scholar] [CrossRef]
- Li, C.; Frolking, S.; Frolking, T.A. A model of nitrous oxide evolution from soil driven by rainfall events: 1. Model structure and sensitivity. J. Geophys. Res. 1992, 97, 9759–9776. [Google Scholar] [CrossRef]
- Li, C.; Trettin, C.; Sun, G.; McNulty, S.; Butterbach-Bahl, K. Modeling carbon and nitrogen biogeochemistry in forest ecosystems. In Proceedings of the 3rd International Nitrogen Conference, Nanjing, China, 12–16 October 2004; pp. 893–898. [Google Scholar]
- Werner, C.; Butterbach-Bahl, K.; Haas, E.; Hickler, T.; Kiese, R. A global inventory of N2O emissions from tropical rainforest soils using a detailed biogeochemical model. Glob. Biogeochem. Cycles 2007, 21. [Google Scholar] [CrossRef]
- Status and trends in sustainable forest management in Europe. In Proceedings of the Forest Europe and FAO: State of Europe’s Forests 2015, Ministerial Conference on the Protection of Forests in Europe, FOREST EUROPE Liaison Unit, Madrid, Spain, 20–21 October 2015; p. 314.
- Pilli, R.; Grassi, G.; Kurz, W.A.; Fiorese, G.; Cescatti, A. The European forest sector: Past and future carbon budget and fluxes under different management scenarios. Biogeosciences 2017, 14, 2387–2405. [Google Scholar] [CrossRef] [Green Version]
- Porté, A.; Bartelink, H. Modelling mixed forest growth: A review of models for forest management. Ecol. Model. 2002, 150, 141–188. [Google Scholar] [CrossRef]
- Groenendijk, M.; Dolman, A.; Van der Molen, M.; Leuning, R.; Arneth, A.; Delpierre, N.; Gash, J.; Lindroth, A.; Richardson, A.; Verbeeck, H. Assessing parameter variability in a photosynthesis model within and between plant functional types using global Fluxnet eddy covariance data. Agric. For. Meteorol. 2011, 151, 22–38. [Google Scholar] [CrossRef]
- Molina-Herrera, S.; Grote, R.; Santabárbara-Ruiz, I.; Kraus, D.; Klatt, S.; Haas, E.; Kiese, R.; Butterbach-Bahl, K. Simulation of CO2 Fluxes in European Forest Ecosystems with the Coupled Soil-Vegetation Process Model “LandscapeDNDC”. Forests 2015, 6, 1779–1809. [Google Scholar] [CrossRef] [Green Version]
- Perkins, D.; Uhl, E.; Biber, P.; du Toit, B.; Carraro, V.; Rötzer, T.; Pretzsch, H. Impact of climate trends and drought events on the growth of oaks (Quercus robur L. and Quercus petraea (Matt.) Liebl.) within and beyond their natural range. Forests 2018, 9, 108. [Google Scholar] [CrossRef] [Green Version]
- Landuyt, D.; Perring, M.P.; Seidl, R.; Taubert, F.; Verbeeck, H.; Verheyen, K. Modelling understorey dynamics in temperate forests under global change—Challenges and perspectives, Perspect. Plant Ecol. Evol. Syst. 2018, 31, 44–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moncrieff, J.B.; Massheder, J.; De Bruin, H.; Elbers, J.; Friborg, T.; Heusinkveld, B.; Kabat, P.; Scott, S.; Søgaard, H.; Verhoef, A. A system to measure surface fluxes of momentum, sensible heat, water vapour and carbon dioxide. J. Hydrol. 1997, 188, 589–611. [Google Scholar] [CrossRef]
- Aubinet, M.; Grelle, A.; Ibrom, A.; Rannik, Ü.; Moncrieff, J.; Foken, T.; Kowalski, A.; Martin, P.; Berbigier, P.; Bernhofer, C. Estimates of the Annual Net Carbon and Water Exchange of Forests: The EUROFLUX Methodology. Adv. Ecol. Res. 2000, 30, 113–175. [Google Scholar]
- Benham, S.; Vanguelova, E.; Pitman, R. Short and long term changes in carbon, nitrogen and acidity in the forest soils under oak at the Alice Holt Environmental Change Network site. Sci. Total. Environ. 2012, 421, 82–93. [Google Scholar] [CrossRef]
- Wilkinson, M.; Eaton, E.; Broadmeadow, M.; Morison, J.I.L. Inter-annual variation of carbon uptake by a plantation oak woodland in south-eastern England. Biogeosciences 2012, 9, 5373–5389. [Google Scholar] [CrossRef] [Green Version]
- Pitman, R.; Broadmeadow, M. Leaf Area, Biomass and Physiological Parameterisation of Ground Vegetation of Lowland Oak Woodland; Forestry Commission: Edinburgh, UK, 2001. Available online: https://www.forestresearch.gov.uk/ (accessed on 1 November 2021).
- Pyatt, D.G. Soil Classification, Forestry Commission Research Information Note 68/82/SSN; Forestry Commission: Edinburgh, UK, 1982.
- Grote, R.; Korhonen, J.; Mammarella, I. Challenges for evaluating process-based models of gas exchange at forest sites with fetches of various species. For. Syst. 2011, 20, 389–406. [Google Scholar] [CrossRef] [Green Version]
- Bossel, H. TREEDYN3 forest simulation model. Ecol. Model. 1996, 90, 187–227. [Google Scholar] [CrossRef]
- Li, C.; Aber, J.; Stange, F.; Papen, H.; Butterbach-Bahl, K. A process-oriented model of N2O and NO emissions from forest soils: 1. Model development. J. Geophys. Res. Atmos. 2000, 105, 4369–4384. [Google Scholar] [CrossRef]
- Dirnböck, T.; Kraus, D.; Grote, R.; Klatt, S.; Kobler, J.; Schindlbacher, A.; Seidl, R.; Thom, D.; Kiese, R. Substantial understory contribution to the C sink of a European temperate mountain forest landscape. Landsc. Ecol. 2020, 35, 483–499. [Google Scholar] [CrossRef] [Green Version]
- Grote, R.; Lehmann, E.; Brümmer, C.; Brüggemann, N.; Szarzynski, J.; Kunstmann, H. Modelling and observation of biosphere–atmosphere interactions in natural savannah in Burkina Faso, West Africa. Phys. Chem. Earth, Parts A/B/C 2009, 34, 251–260. [Google Scholar] [CrossRef]
- Kiese, R.; Heinzeller, C.; Werner, C.; Wochele, S.; Grote, R.; Butterbach-Bahl, K. Quantification of nitrate leaching from German forest ecosystems by use of a process oriented biogeochemical model. Environ. Pollut. 2011, 159, 3204–3214. [Google Scholar] [CrossRef]
- Field, C.; Mooney, H. On the Economy of Plant Form and Function. Proceedings of the Sixth Maria Moors Cabot Symposium, “Evolutionary Constraints on Primary Productivity: Adaptive Patterns of Energy Capture in Plants”; Givnish, T.J., Ed.; Cambridge University Press: Cambridge, MA, USA, 1986; pp. 25–56. [Google Scholar]
- Kattge, J.; Knorr, W.; Raddatz, T.; Wirth, C. Quantifying photosynthetic capacity and its relationship to leaf nitrogen content for global-scale terrestrial biosphere models. Glob. Chang. Biol. 2009, 15, 976–991. [Google Scholar] [CrossRef]
- Farquhar, G.; von Caemmerer, S.v.; Berry, J. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckley, T.; Mott, K.; Farquhar, G. A hydromechanical and biochemical model of stomatal conductance. Plant Cell Environ. 2003, 26, 1767–1785. [Google Scholar] [CrossRef] [Green Version]
- Thornley, J.H.M.; Cannell, M.G.R. Modelling the components of plant respiration: Representation and realism. Ann. Bot. 2000, 85, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Cermak, J. Leaf distribution in large trees and stands of the floodplain forest in southern Moravia. Tree Physiol. 1998, 18, 727–737. [Google Scholar] [CrossRef]
- Broadmeadow, M.; Pitman, R.; Jackson, S.; Randle, T.; Durrant, D.; Lodge, A.H. Upgrading the Level II Protocol for Physiological Modelling of Cause-Effect Relationships: A Pilot Study; Forestry Commission: Edinburgh, UK, 2000.
- Wilkinson, M.; Crow, P.; Eaton, E.L.; Morison, J.I.L. Effects of management thinning on CO2 exchange by a plantation oak woodland in south-eastern England. Biogeosciences 2016, 13, 2367–2378. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.; Smith, J.; Powlson, D.; McGill, W.; Arah, J.; Chertov, O.; Coleman, K.; Franko, U.; Frolking, S.; Jenkinson, D. A comparison of the performance of nine soil organic matter models using datasets from seven long-term experiments. Geoderma 1997, 81, 153–225. [Google Scholar] [CrossRef]
- Nash, J.E.; Sutcliffe, J.V. River flow forecasting through conceptual models, Part I—A discussion of principles. J. Hydrol. 1970, 10, 282–290. [Google Scholar] [CrossRef]
- Oren, R.; Hsieh, C.; Stoy, P.; Albertson, J.; McCarthy, H.R.; Harrell, P.; Katul, G.G. Estimating the uncertainty in annual net ecosystem carbon exchange: Spatial variation in turbulent fluxes and sampling errors in eddy-covariance measurements. Glob. Chang. Biol. 2006, 12, 883–896. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.; Albanito, F.; Bell, M.; Bellarby, J.; Blagodatskiy, S.; Datta, A.; Dondini, M.; Fitton, N.; Flynn, H.; Hastings, A.; et al. Systems approaches in global change and biogeochemistry research. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2012, 367, 311–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamulki, S.; Morison, J.I.L. Annual greenhouse gas fluxes from a temperate deciduous oak forest floor. Forestry 2017, 90, 541–552. [Google Scholar] [CrossRef]
- Yamulki, S.; Anderson, R.; Peace, A.; Morison, J.I.L. Soil CO2, CH4 and N2O fluxes from an afforested lowland raised peatbog in Scotland: Implications for drainage and restoration. Biogeosciences 2013, 10, 1051–1065. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, J.; Taylor, J. On the temperature dependence of soil respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Valentini, R.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Chang. Biol. 2005, 11, 1–16. [Google Scholar] [CrossRef]
- Butterbach-Bahl, K.; Gasche, R.; Breuer, L.; Papen, H. Fluxes of NO and N2O from temperate forest soils: Impact of forest type, N deposition and of liming on the NO and N2O emissions. Nutr. Cycl. Agroecosyst. 1997, 48, 79–90. [Google Scholar] [CrossRef]
- Taylor, J.; Wilson, B.; Mills, M.S.; Burns, R.G. Comparison of microbial numbers and enzymatic activities in surface soils and subsoils using various techniques. Soil Biol. Biochem. 2002, 34, 387–401. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Stange, F.; Butterbach-Bahl, K.; Papen, H.; Zechmeister-Boltenstern, S.; Li, C.; Aber, J. A process-oriented model of N2O and NO emissions from forest soils. 2. Sensitivity analysis and validation. J. Geophys. Res. 2000, 105, 4385–4398. [Google Scholar] [CrossRef] [Green Version]
- Saggar, S.; Andrew, R.M.; Tate, K.R.; Hedley, C.B.; Rodda, N.J.; Townsend, J.A. Modelling nitrous oxide emissions from dairy-grazed pastures. Nutr. Cycl. Agroecosys. 2004, 68, 243–255. [Google Scholar] [CrossRef]
- Li, C.; Farahbakhshazad, N.; Jaynes, D.B.; Dinnes, D.L.; Salas, W.; McLaughlin, D. Modeling nitrate leaching with a biogeochem-ical model modified based on observations in a row-crop field in Iowa. Ecol. Model. 2006, 196, 116–130. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, C.; Shang, Q.; Su, L.; Wang, L. Responses of soil respiration to spring drought and precipitation pulse in a temperate oak forest. Agric. For. Meteorol. 2019, 268, 289–298. [Google Scholar] [CrossRef]
- Goulden, M.L.; Munger, J.W.; Fan, S.-M.; Daube, B.C.; Wofsy, S.C. Exchange of carbon dioxide by a deciduous forest: Response to interannual climate variability. Science 1996, 271, 1576–1578. [Google Scholar] [CrossRef] [Green Version]
- Pumpanen, J.; Kolari, P.; Ilvesniemi, H.; Minkkinen, K.; Vesala, T.; Niinistö, S.; Lohila, A.; Larmola, T.; Morero, M.; Pihlatie, M.; et al. Comparison of different chamber techniques for measuring soil CO2 efflux. Agric. For. Meteorol. 2004, 123, 159–176. [Google Scholar] [CrossRef]
- Levy, P.E.; Gray, A.; Leeson, S.R.; Gaiawyn, J.; Kelly, M.P.C.; Cooper, M.D.A.; Dinsmore, K.J.; Jones, S.K.; Sheppard, L.J. Quantification of uncertainty in trace gas fluxes measured by the static chamber method. Eur. J. Soil Sci. 2011, 62, 811–821. [Google Scholar] [CrossRef]
- Heinemeyer, A.; Wilkinson, M.; Vargas, R.; Subke, J.; Casella, E.; Morison, J.I.; Ineson, P. Exploring the “overflow tap” theory: Linking forest soil CO2 fluxes and individual mycorrhizosphere components to photosynthesis. Biogeosciences 2012, 9, 79–95. [Google Scholar] [CrossRef] [Green Version]
- Rochette, P.; Eriksen-Hamel, N.S. Chamber measurements of soil nitrous oxide flux: Are absolute values reliable? Soil Sci. Soc. Am. J. 2008, 72, 331–342. [Google Scholar] [CrossRef]
- Venterea, R.T.; Spokas, K.A.; Baker, J.M. Accuracy and precision analysis of chamber-based nitrous oxide gas flux estimates. Soil Sci. Soc. Am. J. 2009, 73, 1087–1093. [Google Scholar] [CrossRef] [Green Version]
- Vanguelova, E.; Benham, S.; Pitman, R.; Moffat, A.; Broadmeadow, M.; Nisbet, T.; Durrant, D.; Barsoum, N.; Wilkinson, M.; Bochereau, F. Chemical fluxes in time through forest ecosystems in the UK–Soil response to pollution recovery. Environ. Pollut. 2010, 158, 1857–1869. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Value |
|---|---|
| Latitude | 51° 09′ N |
| Longitude | 0° 51′ W |
| Average annual rainfall | 877 mm |
| Average annual temperature | 10.3 °C |
| Annual N deposition as NH4 | 0.89 g m−3 |
| Annual N deposition as NO3 | 0.51 g m−3 |
| Total annual N deposition | 1.4 g m−3 |
| Slope | 0° |
| Altitude | 80.0 m AMSL |
| Upper storey trees: main species | Pedunculate oak (Q. robur) |
| Number of trees per hectare | 442 |
| Height | 16.5 m |
| Diameter at breast height | 0.233 m |
| Understorey trees: species (aggregated, for PSIM only) | European Ash (F. excelsior) (representing hazel, hawthorn, ash and holly) |
| Number of trees per hectare | 8000 |
| Height | 3.0 m |
| Diameter at breast height | 0.03 m |
| Modelling start date | 1/1/1995 |
| Thinning event (eastern half of Inclosure) | 20/9/2007 |
| Proportion of stemwood removed by thinning | 0.3 for upper storey; 0.6 for understorey |
| Horizon | Thickness mm | Organic C | Organic N | pH | Bulk Density g cm−3 | Field Capacity mm m−3 | Wilting Point mm m−3 | Clay Fraction |
|---|---|---|---|---|---|---|---|---|
| O | 20 | 0.2162 | 0.0114 | 5.1 | 0.0670 | - | - | 0.035 |
| A | 80 | 0.0560 | 0.0038 | 5.4 | 0.7043 | 530 | 240 | 0.520 |
| E | 80 | 0.0287 | 0.0023 | 5.2 | 0.9682 | 530 | 240 | 0.516 |
| B | 200 | 0.0159 | 0.0010 | 5.4 | 1.1334 | 480 | 240 | 0.510 |
| BC | 380 | 0.0108 | 0.0003 | 6.2 | 0.9350 | 530 | 240 | 0.601 |
| C | 260 | 0.0146 | 0.0005 | 5.4 | 1.0123 | 520 | 240 | 0.578 |
| Parameter | Description | Units | PnET | PSIM (Understorey Value) |
|---|---|---|---|---|
| CO2 exchange parameters | ||||
| BASEFOLRESPFRAC | Dark respiration as fraction of photosynthesis | 0–1 | 0.15 | na |
| AMAXB | Maximal net photosynthetic rate | nmol CO2 g−1 s−1/%N | 55 | na |
| RESPQ10 2 | Temperature dependency of leaf respiration | - | 2 | na |
| ROOTMRESPFRAC | Ratio of fine root maintenance respiration to biomass production | - | 2 | na |
| VCMAX25 | Maximum RubP saturated rate of carboxylation at 25 °C for leaves in full sun | µmol m−2 s−1 | na | 90 (85) |
| KM20 | Respiration maintenance coefficient at reference temperature | 0–1 | na | 0.3 (0.1) |
| Phenology-related parameters | ||||
| GDDFOLSTART3 | Daily temperature sum for start of foliage budburst | °days | 500 | 500 (0) |
| GDDFOLEND 3 | Daily temperature sum for end of foliage growth (maximum leaf area) | °days | 1100 | na |
| MFOLOPT 1 | Foliage biomass under optimal, closed canopy conditions | kg DW m−2 | 0.47 | 0.47 (0.36) |
| NDFLUSH 3 | Time required to complete growth of new foliage | days | na | 45 (40) |
| NDMORTA 4 | Time required to complete leaf fall | days | na | 100 (40) |
| DLEAFSHED 4 | Day by which leaf fall is complete | day of year | na | 330 (310) |
| SENESCSTART 4 | Day of year after which leaf death can occur | day of year | 300 | na |
| Resource acquisition parameters | ||||
| NCFOLOPT | Optimum nitrogen concentration of foliage | g N gDW−1 | 0.024 | 0.024 (0.032) |
| EXPL_NH4 | Relative exploitation rate of NH4 | % | 0.3 | na |
| EXPL_NO3 | Relative exploitation rate of NO3 | % | 0.16 | na |
| EXT 5 | Light extinction (attenuation) coefficient | 0–1 | 0.4 | 0.4 (0.65) |
| Year | GPP (g C m−2 year−1) | TER (g C m−2 year−1) | NEP (g C m−2 year−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| EC | PnET | PSIM | EC | PnET | PSIM | EC | PnET | PSIM | |
| 1999 | 1983 | 2440 | 2204 | 1625 | 1805 | 1638 | 357 | 634 | 566 |
| 2000 | 2346 | 2127 | 2046 | 1940 | 1641 | 1588 | 406 | 487 | 458 |
| 2001 | 2227 | 2089 | 2037 | 1670 | 1600 | 1576 | 557 | 489 | 462 |
| 2002 | 2180 | 2062 | 2171 | 1767 | 1545 | 1650 | 412 | 517 | 521 |
| 2003 | 2223 | 1666 | 2173 | 1606 | 1360 | 1542 | 617 | 307 | 631 |
| 2004 | 2172 | 1856 | 2102 | 1573 | 1423 | 1599 | 600 | 433 | 503 |
| 2005 | 1992 | 1697 | 2109 | 1441 | 1348 | 1601 | 551 | 349 | 508 |
| 2006 | 1862 | 1671 | 2077 | 1374 | 1385 | 1635 | 488 | 286 | 442 |
| 2007 | 2094 | 1851 | 2148 | 1466 | 1386 | 1614 | 629 | 466 | 534 |
| Mean | 2120 | 1940 | 2119 | 1607 | 1499 | 1605 | 513 | 441 | 514 |
| SD | 151 | 261 | 59 | 175 | 158 | 34 | 101 | 111 | 59 |
| % Difference | −8.5 | −0.06 | −6.7 | −0.13 | −14.1 | 0.17 | |||
| (a) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Statistic | Annual GPP 1999–2007 | Annual TER 1999–2007 | Annual NEP 1999–2007 | |||||
| PnET | PSIM | PnET | PSIM | PnET | PSIM | |||
| RMSE (%) | 14.8 + | 7.8 + | 10.9 + | 10.7 + | 37.4 + | 19.5 + | ||
| RMSE (au) | 313.1 | 165.1 | 174.5 | 172.3 | 191.8 | 99.9 | ||
| ME | −3.85 | −0.35 | −0.13 | −0.10 | −3.05 | −0.10 | ||
| CD | 0.22 | 6.52 | 0.80 | 26.3 | 0.56 | 2.92 | ||
| Relative Error (E; %) | 8.24 * | −0.44 * | 6.34 * | −0.92 * | 7.91 * | −3.84 * | ||
| Mean Difference (M; au) | 180.0 | 1.33 | 107.7 | 2.11 | 72.11 | −0.89 | ||
| Student’s t of M (t) | 2.0 * | 0.02 * | 2.22 * | 0.03 * | 1.15 * | −0.03 * | ||
| Correlation coefficient (r) | 0.21 | −0.25 | 0.62 | −0.15 | −0.58 | 0.21 | ||
| (b) | ||||||||
| Statistic | Monthly GPP 1999–2007 | Monthly TER 1999–2007 | Monthly NEP 1999–2007 | |||||
| PnET | PSIM | PnET | PSIM | PnET | PSIM | |||
| RMSE (%) | 27.81 + | 23.30 + | 36.6 + | 28.9 + | 115.6 + | 110.4 + | ||
| RMSE (mu) | 49.13 | 41.16 | 49.0 | 38.7 | 49.4 | 47.16 | ||
| ME | 0.92 | 0.94 | 0.47 | 0.67 | 0.82 | 0.83 | ||
| CD | 1.04 | 1.02 | 0.49 | 0.96 | 2.23 | 1.19 | ||
| Relative Error (E; %) | 39.3 * | 17.83 * | 18.6 * | −2.63 * | 34.0 * | −52.9 * | ||
| Mean Difference (M; mu) | 14.99 | 0.10 | 8.98 | 0.18 | 6.08 | −0.05 | ||
| Student’s t of M (t) | 1.49 * | 0.01 * | 1.93 * | 0.05 * | 0.87 * | −0.01 * | ||
| Correlation coefficient (r) | 0.96 | 0.97 | 0.88 | 0.84 | 0.95 | 0.91 | ||
| (c) | ||||||||
| Statistic | Monthly Soil CO2 | Monthly N2O | ||||||
| 2008–2012 | 2013–14 | 2008–2012 | 2013–14 | |||||
| Model | PnET | PSIM | PnET | PSIM | PnET | PSIM | PnET | PSIM |
| RMSE (%) | 60.1 + | 87.4 + | 28.9 | 68.1 | 154 + | 147 + | 61.0 | 42.0 |
| RMSE (mu) | 47.2 | 68.5 | 18.8 | 44.4 | 14.4 | 13.8 | 2.8 | 1.9 |
| ME | −0.16 | 0.38 | 0.08 | −4.11 | −0.19 | −0.09 | −2.25 | −0.54 |
| CD | 1.19 | 1.20 | 0.52 | 0.14 | 5.16 | 5.89 | 0.83 | 0.83 |
| Relative Error (E; %) | 36.2 * | −24.0 * | 14.4 * | −49.0 | 76.6 * | 50.9 * | −15.3 * | 13.4 * |
| Mean Difference (M; mu) | 33.1 | −8.1 | 8.25 | −31.9 | 5.6 | 5.2 | 0.06 | 0.87 |
| Student’s t of M (t) | 6.77 | −1.64 * | 1.89 * | −3.99 | 2.75 | 2.70 | 0.09 * | 2.00 |
| Correlation coefficient (r) | 0.66 | 0.68 | 0.76 | 0.68 | 0.03 | 0.36 | −0.47 | 0.35 |
| No. of values | 48 | 48 | 16 | 16 | 44 | 44 | 16 | 16 |
| Year | Soil CO2 (g C m−2 year−1) | Soil N2O (mg Nm−2 year−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SC | PnET | PnET | PSIM | PSIM | SC | PnET | PnET | PSIM | PSIM | |
| Thinning: | 0% | 30% | 0% | 30% | 0% | 30% | 0% | 30% | ||
| 2006 | Na | 571 | 571 | 1081 | 1081 | Na | 49.1 | 49.1 | 57.3 | 57.3 |
| 2007 | Na | 561 | 629 | 1059 | 1026 | Na | 47.0 | 41.7 | 55.2 | 52.7 |
| 2008 | 1015.5 | 514 | 758 | 1033 | 904 | Na | 52.6 | 56.5 | 60.3 | 61.1 |
| 2009 | 697.0 | 543 | 682 | 1046 | 888 | 155.7 | 43.8 | 61.1 | 52.3 | 61.3 |
| 2010 | 797.3 | 496 | 547 | 934 | 793 | 33.8 | 38.9 | 51.8 | 40.7 | 37.6 |
| 2011 | 835.8 | 598 | 586 | 1078 | 926 | 15.4 | 41.4 | 51.6 | 43.9 | 37.1 |
| 2012 | Na 1 | 514 | 488 | 1007 | 884 | Na 1 | 48.2 | 50.5 | 44.7 | 36.4 |
| 2013 | 689.3 2 | 523 | 473 | 941 | 871 | 57.4 2 | 41.7 | 43.3 | 36.9 | 32.7 |
| 2014 | Na | 638 | 554 | 1087 | 998 | Na | 55.2 | 52.1 | 51.5 | 46.1 |
| Mean | 807 | 547 | 584 | 1018 | 897 | 65.6 | 46.0 | 52.4 | 47.2 | 46.0 |
| SD | 132.6 | 52.0 | 103.0 | 61.2 | 67.3 | 62.5 | 6.2 | 5.5 | 8.0 | 12.6 |
| Difference | - | −32% | 26% | - | −30% | −28% | ||||
| PnET | PSIM | |||
|---|---|---|---|---|
| Level of Thinning: | 0% | 30% | 0% | 30% |
| 2009 | ||||
| RMSE (%) | 28.8 + | 39.2 + | 36.5 + | 39.3 + |
| Average total error | 31.7 | 57.9 | 40.1 | 58.2 |
| Modelling efficiency | 0.73 | 0.46 | 0.56 | 0.45 |
| Coefficient of determination | 0.61 | 0.83 | 0.69 | 1.16 |
| Correlation coefficient, r | 0.92 | 0.85 | 0.88 | 0.80 |
| No. of values | 12 | 12 | 12 | 12 |
| 2012 | ||||
| RMSE (%) | 39.4 + | 48.1 + | 26.5 + | 33.5 + |
| Average total error | 54.9 | 68.8 | 37.1 | 47.8 |
| Modelling efficiency | 0.51 | 0.18 | 0.78 | 0.60 |
| Coefficient of determination | 0.91 | 0.78 | 1.31 | 1.19 |
| Correlation coefficient, r | 0.91 | 0.90 | 0.89 | 0.92 |
| No. of values | 12 | 12 | 12 | 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cade, S.M.; Clemitshaw, K.C.; Molina-Herrera, S.; Grote, R.; Haas, E.; Wilkinson, M.; Morison, J.I.L.; Yamulki, S. Evaluation of LandscapeDNDC Model Predictions of CO2 and N2O Fluxes from an Oak Forest in SE England. Forests 2021, 12, 1517. https://doi.org/10.3390/f12111517
Cade SM, Clemitshaw KC, Molina-Herrera S, Grote R, Haas E, Wilkinson M, Morison JIL, Yamulki S. Evaluation of LandscapeDNDC Model Predictions of CO2 and N2O Fluxes from an Oak Forest in SE England. Forests. 2021; 12(11):1517. https://doi.org/10.3390/f12111517
Chicago/Turabian StyleCade, Shirley M., Kevin C. Clemitshaw, Saúl Molina-Herrera, Rüdiger Grote, Edwin Haas, Matthew Wilkinson, James I. L. Morison, and Sirwan Yamulki. 2021. "Evaluation of LandscapeDNDC Model Predictions of CO2 and N2O Fluxes from an Oak Forest in SE England" Forests 12, no. 11: 1517. https://doi.org/10.3390/f12111517
APA StyleCade, S. M., Clemitshaw, K. C., Molina-Herrera, S., Grote, R., Haas, E., Wilkinson, M., Morison, J. I. L., & Yamulki, S. (2021). Evaluation of LandscapeDNDC Model Predictions of CO2 and N2O Fluxes from an Oak Forest in SE England. Forests, 12(11), 1517. https://doi.org/10.3390/f12111517