Hybrid Pine (Pinus attenuata × Pinus radiata) Somatic Embryogenesis: What Do You Prefer, Mother or Nurse?

 ,
,  ,
,  , and
, and
Abstract
1. Introduction
- Does the presence of initiation environment inhibit or enhance the success of the process?
- What is the effect of the mother (i.e., P. radiata or P. attenuata as the female parent) in embryonal mass conversion to somatic plants?
2. Materials and Methods
2.1. Plant Material
2.2. Culture Conditions and Initiation of Embryonal Masses
- Treatment 1:
- Megagametophyte + Glitz medium
- Treatment 2:
- Excised zygotic embryo (ZE) + Glitz medium
- Treatment 3:
- Excised ZE + Glitz medium + Nurse
2.3. Maturation and Germination of Embryos
2.4. Acclimatization
2.5. Long Term Conservation at −80 °C
2.6. Data Collection and Analysis
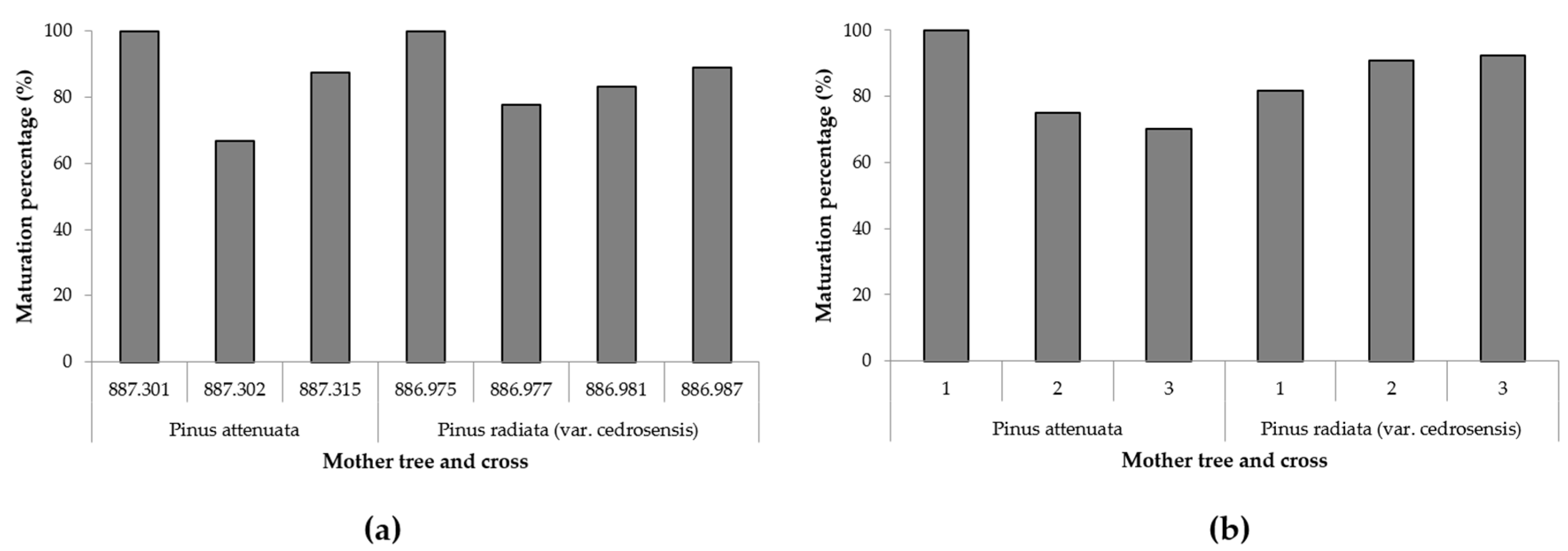
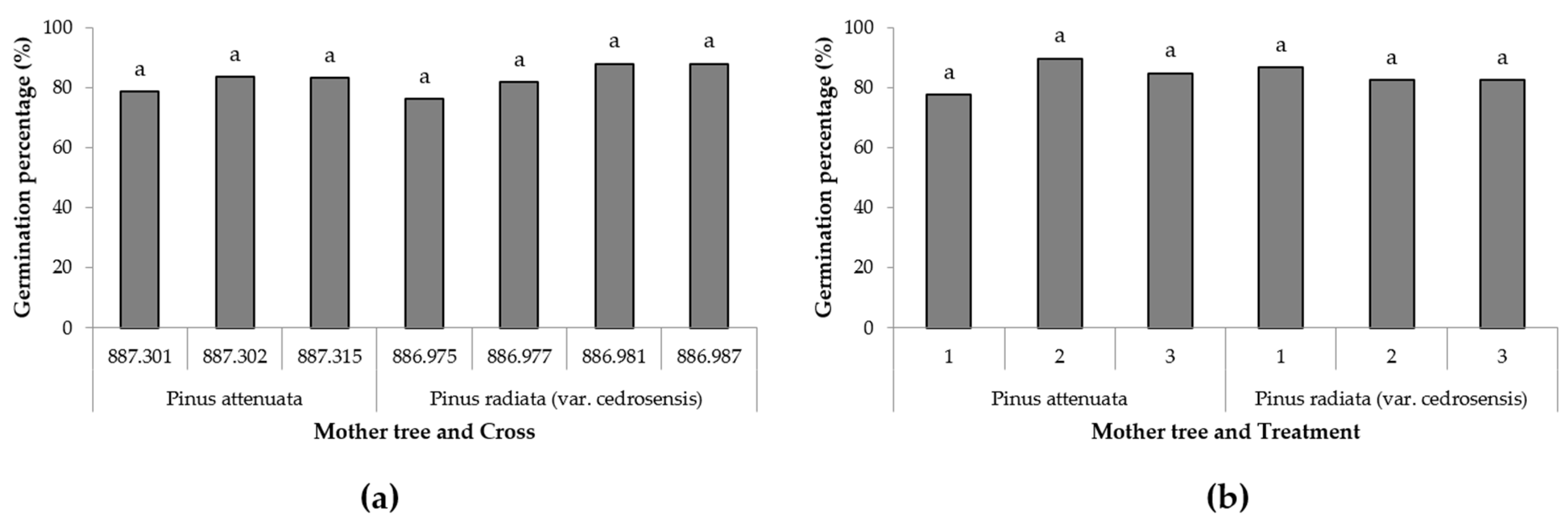
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mitchell, R.G. Reducing the Risks of Pitch Canker Disease (Caused by Fusarium circinatum) to Pinus patula in South Africa. Ph.D. Thesis, Faculty of Natural and Agricultural Science, University of Pretoria, Pretoria, South Africa, 2012. [Google Scholar]
- Mitchell, R.G.; Wingfield, M.J.; Hodge, G.R.; Dvorak, W.S.; Continho, T.A. The tolerance of Pinus patula × Pinus tecunumanii, and other pine hybrids, to Fusarium circinatum in greenhouse trials. New For. 2013, 44, 443–456. [Google Scholar] [CrossRef]
- Camcore. Progress in pine hybrid testing. In 2012 Camcore Annual Report; Department of Forestry and Environmental Resources, NC State University: Raleigh, NC, USA, 2012; p. 18. [Google Scholar]
- Camcore. Status of pine hybrid project. In 2013 Camcore Annual Report; Department of Forestry and Environmental Resources, NC State University: Raleigh, NC, USA, 2013; p. 20. [Google Scholar]
- Sheffield, J.; Wood, E.F. Global trends and variability in soil moisture and drought characteristics, 1950–2000, from observation-driven simulations of the terrestrial hydrologic cycle. J. Clim. 2008, 21, 432–458. [Google Scholar] [CrossRef]
- Dungey, H.S.; Carson, M.J.; Low, C.B.; King, N.G. Potential and niches for inter-specific hybrids with Pinus radiata in New Zealand. N. Z. J. For. Sci. 2003, 33, 295–318. [Google Scholar]
- Hargreaves, C.; Sigley, M.; Menzies, M.; Dungey, H. Hybrid pines: Opportunities for life on the edge. Proc. Int. Plant Propagators Soc. 2008, 58, 62–167. [Google Scholar]
- Hargreaves, C.; Reeves, C.; Gough, K.; Montalbán, I.A.; Low, C.; van Ballekom, S.; Dungey, H.S.; Moncaleán, P. Nurse tissue for embryo rescue: Testing new conifer somatic embryogenesis methods in a F1 hybrid pine. Trees 2017, 31, 273–283. [Google Scholar] [CrossRef]
- De Diego, N.; Pérez-Alfocea, F.; Cantero, E.; Lacuesta, M.; Moncaleán, P. Physiological response to drough in radiata pine: Phytohormone implication at leaf level. Tree Physiol. 2012, 32, 435–449. [Google Scholar] [CrossRef] [PubMed]
- De Diego, N.; Sampedro, M.C.; Barrio, R.J.; Saiz-Fernández, I.; Moncaleán, P.; Lacuesta, M. Solute accumulation and elastic modulus changes in six radiata pine breeds exposed to drought. Tree Physiol. 2013, 33, 69–80. [Google Scholar] [CrossRef]
- De Diego, N.; Rodríguez, J.L.; Dodd, I.C.; Pérez-Alfocea, F.; Moncaleán, P.; Lacuesta, M. Immunolocalization of IAA and ABA in roots and needles of radiata pine (Pinus radiata) during drought and rewatering. Tree Physiol. 2013, 33, 537–549. [Google Scholar] [CrossRef]
- De Diego, N.; Saiz-Fernández, I.; Rodríguez, J.L.; Pérez-Alfocea, P.; Sampedro, M.C.; Barrio, R.J.; Lacuesta, M.; Moncaleán, P. Metabolites and hormones are involved in the intraspecific variability of drought hardening in radiata pine. J. Plant Physiol. 2015, 188, 64–71. [Google Scholar] [CrossRef]
- Hargreaves, C.L.; Reeves, C.B.; Find, J.I.; Gough, K.; Josekutty, P.; Skudder, D.B.; van der Maas, S.A.; Sigley, M.R.; Menzies, M.I.; Low, C.B.; et al. Improving initiation, genotype capture, and family representation in somatic embryogenesis of Pinus radiata by a combination of zygotic embryo maturity, media, and explant preparation. Can. J. For. Res. 2009, 39, 1566–1574. [Google Scholar] [CrossRef]
- Montalbán, I.A.; De Diego, N.; Moncaleán, P. Bottlenecks in Pinus radiata somatic embryogenesis: Improving maturation and germination. Trees 2010, 24, 1061–1071. [Google Scholar] [CrossRef]
- Montalbán, I.A.; De Diego, N.; Moncaleán, P. Enhancing initiation and proliferation in radiata pine (Pinus radiata D.Don) somatic embryogenesis through seed family screening, zygotic embryo staging and media adjustments. Acta Physiol. Plant. 2012, 34, 451–460. [Google Scholar] [CrossRef]
- Castander-Olarieta, A.; Montalbán, I.A.; De Medeiros Oliveira, E.; Dell’Aversana, E.; D’Amelia, L.; Carillo, P.; Steiner, S.; De Freitas Fraga, H.P.; Guerra, M.P.; Goicoa, T.; et al. Effect of thermal stress on tissue ultrastructure and metabolite profiles during initiation of radiata pine somatic embryogenesis. Front. Plant Sci. 2019, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- García-Mendiguren, O.; Montalbán, I.A.; Goicoa, T.; Ugarte, M.D.; Moncaleán, P. Environmental conditions at the initial stages of Pinus radiata somatic embryogenesis affect the production of somatic embryos. Trees Struct. Funct. 2016, 30, 949–958. [Google Scholar] [CrossRef]
- Pereira, C.; Montalbán, I.A.; García-Mendiguren, O.; Goicoa, T.; Ugarte, M.D.; Correia, S.; Canhoto, J.M.; Moncaleán, P. Pinus halepensis somatic embryogenesis is affected by the physical and chemical conditions at the initial stages of the process. J. For. Res. 2016, 21, 143–150. [Google Scholar] [CrossRef]
- Montalbán, I.A.; García-Mendiguren, O.; Goicoa, T.; Ugarte, M.D.; Moncaleán, P. Cold storage of initial plant material affects positively somatic embryogenesis in Pinus radiata. New For. 2015, 46, 309–317. [Google Scholar] [CrossRef]
- Litvay, J.D.; Verma, D.C.; John, M.A. Influence of loblolly pine (Pinus taeda L.) culture medium and its components on growth and somatic embryogenesis of the wild carrot (Daucus carota L.). Plant Cell Rep. 1985, 4, 325–328. [Google Scholar] [CrossRef]
- Walter, C.; Find, J.I.; Grace, L.J. Somatic embryogenesis and genetic transformation in Pinus radiata. In Protocol for Somatic Embryogenesis in Woody Plants; Jain, S.M., Gupta, P.K., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 11–24. [Google Scholar]
- Quoirin, M.; Lepoivre, P. Études des milieux adaptés aux cultures in vitro de Prunus. Acta Hort. 1977, 78, 437–442. [Google Scholar] [CrossRef]
- Aitken-Christie, J.; Singh, A.P.; Davies, H. Multiplication of meristematic tissue: A new tissue culture system for radiata pine. In Genetic Manipulation of Woody Plants; Hanover, J.W., Keathley, D.E., Eds.; Plenum: New York, NY, USA, 1988; pp. 413–432. [Google Scholar]
- Montalbán, I.A.; Moncaleán, P. Long term conservation at −80 °C of Pinus radiata embryogenic cell lines: Recovery, maturation and germination. CryoLetters 2017, 38, 202–209. [Google Scholar]
- Hargreaves, C.L.; Grace, L.J.; Holden, D.G. Nurse culture for efficient recovery of cryopreserved Pinus radiata D. Don embryogenic cell lines. Plant Cell Rep. 2002, 21, 40–45. [Google Scholar]
- Pereira, C.; Castander-Olarieta, A.; Montalbán, I.A.; Pěnčík, A.; Petřík, I.; Pavlović, I.; Medeiros Oliveira, E.D.; de Freitas Fraga, H.P.; Guerra, M.P.; Novák, O.; et al. Embryonal masses induced at high temperatures in Aleppo pine: Cytokinin profile and cytological characterization. Forests 2020, 11, 807. [Google Scholar] [CrossRef]
- Arrillaga, I.; Morcillo, M.; Zanón, I.; Lario, F.; Segura, J.; Sales, E. New approaches to optimize somatic embryogenesis in maritime pine. Front. Plant Sci. 2019, 10, 138. [Google Scholar] [CrossRef] [PubMed]
- Miguel, C.; Gonçalves, S.; Tereso, S.; Marum, L.; Maroco, J.; Oliveira, M.M. Somatic embryogenesis from 20 open-pollinated families of Portuguese plus trees of maritime pine. Plant Cell Tissue Org. Cult. 2004, 76, 121–130. [Google Scholar] [CrossRef]
- Niskanen, A.-M.; Lu, J.; Seitz, S.; Keinonen, K.; Von Weissenberg, K.; Pappinen, A. Effect of parent genotype on somatic embryogenesis in Scots pine (Pinus sylvestris). Tree Physiol. 2004, 24, 1259–1265. [Google Scholar] [CrossRef] [PubMed]
- Carneros, E.; Celestino, C.; Klimaszewska, K.; Park, Y.-S.; Toribio, M.; Bonga, J.M. Plant regeneration in Stone pine (Pinus pinea L.) by somatic embryogenesis. Plant Cell Tissue Org. Cult. 2009, 98, 165–178. [Google Scholar] [CrossRef]
- Salaj, T.; Moravcikova, J.; Salaj, J. Somatic embryogenesis in Pinus nigra arn.: Some physiological, structural and molecular aspects. Plant Cell Mongr. 2005, 2, 141–156. [Google Scholar]
- Salaj, T.; Matusova, R.; Salaj, J. Conifer somatic embryogenesis—An efficient plant regeneration system for theoretical studies and mass propagation. Dendrobiology 2015, 74, 69–76. [Google Scholar] [CrossRef]
- Nunes, S.; Marum, L.; Farinha, N.; Pereira, V.T.; Almeida, T.; Sousa, D.; Mano, N.; Figueiredo, J.; Dias, M.C.; Santos, C. Somatic embryogenesis of hybrid Pinus elliottii var. elliottii × P. caribaea var. hondurensis and ploidy assessment of somatic plants. Plant Cell Tissue Org. Cult. 2018, 132, 71–84. [Google Scholar] [CrossRef]
- Egertsdotter, U. Plant physiological and genetical aspects of the somatic embryogenesis process in conifers. Scan. J. For. Res. 2019, 34, 360–369. [Google Scholar] [CrossRef]
- Miller, J.T.; Knowles, F.B. Introduced Forest Trees in New Zealand: Recognition, Role and Seed Source Pinus attenuata Lemmon—Knobcone Pine; New Zealand Ministry of Forestry, Forest Research Institute: Rotorua, New Zealand, 1988. [Google Scholar]
- Lüttge, U. Father hands it down: Patrilineal inheritance of chloroplast DNA in conifers may help to uncover relations of reproduction, population biology and longevity of trees. Trees Struct. Func. 2016, 30, 73–74. [Google Scholar] [CrossRef][Green Version]
- Kormutak, A.; Galgoci, M.; Sukenikova, D.; Bolecek, P.; Libantova, J.; Gőmőry, D. Maternal inheritance of chloroplast DNA in Pinus mugo Turra: A case study of Pinus mugo × Pinus sylvestris crossing. Plant Syst. Evol. 2018, 304, 71–76. [Google Scholar] [CrossRef]
- Chamberland, V.; Robichaud, F.; Perron, M.; Gélinas, N.; Bousquet, J.; Beaulieu, J. Conventional versus genomic selection for white spruce improvement: A comparison of costs and benefits of plantations on Quebec public lands. Tree Genet. Genomes 2020, 16, 17. [Google Scholar] [CrossRef]
- Krajňáková, J.; Bertolini, A.; Gömöry, D.; Vianello, A.; Häggman, H. Initiation, long-term cryopreservation, and recovery of Abies alba Mill. embryogenic cell lines. Cel. Dev. Biol. Plant 2013, 49, 560–571. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Chemicals (mg/L) | Glitz | EDM | LP ½ Modified |
|---|---|---|---|
| Ca(NO3)2·4H2O | 417.5 | ||
| CaCl2·2H2O | 11 | 25 | |
| KNO3 | 950 | 1431 | 900 |
| MgSO4·7H2O | 925 | 400 | 220 |
| NaNO3 | 310 | ||
| NH4H2PO4 | 225 | ||
| NH4NO3 | 825 | 200 | |
| KH2PO4 | 170 | 337.5 | |
| FeSO4·7H2O | 30 | 30 | 30 |
| Na2EDTA·2H2O | 40 | 40 | 40 |
| ZnSO4·7H2O | 43 | 25 | 8.6 |
| H3BO3 | 31 | 8 | 6.2 |
| MnSO4·4H2O | 21 | 3.6 | 1 |
| CuSO4·5H2O | 0.5 | 2.4 | 0.025 |
| KI | 4.15 | 1 | 0.08 |
| CoCl2·6H2O | 0.125 | 0.2 | 0.025 |
| Na2MoO4·H2O | 1.25 | 0.2 | 0.25 |
| Thiamine HCl | 5 | 5 | 0.1 |
| Nicotinic acid | 5 | 5 | 0.5 |
| Pyridoxine HCl | 0.5 | 0.5 | 0.5 |
| Casein hydrolisate | 1000 | ||
| Glutamine * | 500 | 550 | |
| Asparagine * | 525 | ||
| Arginine * | 175 | ||
| L-Citrulline * | 19.75 | ||
| L-Ornithine * | 19 | ||
| L-Lysine * | 13.74 | ||
| L-Alanine * | 10 | ||
| L-Proline * | 8.75 | ||
| Myo-inositol | 100 | 1000 | 100 |
| Sucrose | 30,000 | 60,000 | 30,000 |
| Medium pH | 5.7 | 5.7 | 5.8 |
| Mother Tree | Cross | Sample Mean ± Standard Error |
|---|---|---|
| P. attenuata | 887.301 | 80.728 ± 14.995 |
| P. attenuata | 887.302 | 189.123 ± 37.872 |
| P. attenuata | 887.315 | 376.549 ± 98.988 |
| P. radiata | 886.975 | 353.125 ± 51.390 |
| P. radiata | 886.977 | 230.052 ± 49.358 |
| P. radiata | 886.981 | 329.393 ± 73.431 |
| P. radiata | 886.987 | 248.116 ± 35.880 |
| Mother Tree | Treatment | Sample Mean ± Standard Error |
|---|---|---|
| P. attenuata | 1 | 172.997 ± 33.339 |
| P. attenuata | 2 | 370.332 ± 115.092 |
| P. attenuata | 3 | 97.882 ± 19.893 |
| P. radiata | 1 | 271.589 ± 41.779 |
| P. radiata | 2 | 394.495 ± 74.453 |
| P. radiata | 3 | 211.397 ± 30.642 |
| Source | X2 Test | df | p Value |
|---|---|---|---|
| Cross | 3.163 | 6 | 0.788 |
| Mother | 0.002 | 1 | 0.967 |
| Treatment | 0.533 | 2 | 0.767 |
| Mother × Treatment | 1.312 | 2 | 0.519 |
| Mother Tree | Treatment | Height (cm) |
|---|---|---|
| P. attenuata | 1 | 8.89 ± 1.48 b |
| P. attenuata | 2 | 9.37 ± 2.31 b |
| P. attenuata | 3 | 9.23 ± 1.99 b |
| P. radiata | 1 | 13.99 ± 4.16 a |
| P. radiata | 2 | 13.29 ± 3.33 a |
| P. radiata | 3 | 12.17 ± 3.07 a |
| Mother Tree | Cross | Height (cm) |
|---|---|---|
| P. attenuata | 887.301 | 9.39 ± 0.66 |
| P. attenuata | 887.302 | 9.45 ± 0.42 |
| P. attenuata | 887.315 | 9.03 ± 0.20 |
| P. radiata | 886.975 | 12.35 ± 0.46 |
| P. radiata | 886.977 | 15.67 ± 0.67 |
| P. radiata | 886.981 | 13.26 ± 0.31 |
| P. radiata | 886.987 | 11.96 ± 0.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montalbán, I.A.; Castander-Olarieta, A.; Hargreaves, C.L.; Gough, K.; Reeves, C.B.; van Ballekom, S.; Goicoa, T.; Ugarte, M.D.; Moncaleán, P. Hybrid Pine (Pinus attenuata × Pinus radiata) Somatic Embryogenesis: What Do You Prefer, Mother or Nurse? Forests 2021, 12, 45. https://doi.org/10.3390/f12010045
Montalbán IA, Castander-Olarieta A, Hargreaves CL, Gough K, Reeves CB, van Ballekom S, Goicoa T, Ugarte MD, Moncaleán P. Hybrid Pine (Pinus attenuata × Pinus radiata) Somatic Embryogenesis: What Do You Prefer, Mother or Nurse? Forests. 2021; 12(1):45. https://doi.org/10.3390/f12010045
Chicago/Turabian StyleMontalbán, Itziar A., Ander Castander-Olarieta, Cathy L. Hargreaves, Keiko Gough, Cathie B. Reeves, Shaf van Ballekom, Tomás Goicoa, María Dolores Ugarte, and Paloma Moncaleán. 2021. "Hybrid Pine (Pinus attenuata × Pinus radiata) Somatic Embryogenesis: What Do You Prefer, Mother or Nurse?" Forests 12, no. 1: 45. https://doi.org/10.3390/f12010045
APA StyleMontalbán, I. A., Castander-Olarieta, A., Hargreaves, C. L., Gough, K., Reeves, C. B., van Ballekom, S., Goicoa, T., Ugarte, M. D., & Moncaleán, P. (2021). Hybrid Pine (Pinus attenuata × Pinus radiata) Somatic Embryogenesis: What Do You Prefer, Mother or Nurse? Forests, 12(1), 45. https://doi.org/10.3390/f12010045